

Полинуклеотидная цепь со строго определенной последовательностью нуклеотидов в ней называется первичной структурой.
РНК и ДНК построены из связанных ковалентно рибонуклеотидных (РНК) или дезоксирибонуклеотидных (ДНК) звеньев. Нуклеотиды соединены друг с другом фосфодиэфирной связью между 5’- гидроксильной группой одного нуклеотида и 3’-гидроксильной груп-
пой следующего.
5' |
|
|
|
NH2 |
|
O |
|
|
|
|
|
O P OH |
N |
|
N |
|
|
Ê |
|
N |
|
N |
|
CH2 |
O |
|
|
|
|
|
|
|
|
||
O |
|
OH |
|
O |
|
|
|
|
|
||
O |
P OH |
N |
|
NH |
|
|
Ê |
|
|
||
|
|
N |
|
|
|
|
CH2 |
O |
N |
NH2 |
|
|
|
||||
Фосфодиэфирная |
|
|
|
|
|
связь |
O |
|
OH |
|
|
3'
В молекуле нуклеиновой кислоты выделяют 5’ и 3’-концы. 5’-
конец имеет нуклеотид со свободным гидроксилом у 5 атома углерода пентозы. 3’-конец имеет нуклеотид со свободным гидроксилом у 3
атома пентозы. Последовательность оснований в составе нуклеиновой кислоты читается в направлении от 5’ к 3’-концу. О первом и послед-
нем нуклеотиде говорят, что они находятся на 5’ и 3’ концах соответственно. Для схематического обозначения последовательности нуклеотидов пишут символ нуклеотида, начиная с 5’-конца, например АГТ...
Иногда добавляется символ р или д (рибоза или дезоксирибоза) дТдЦдА… или символы минорных оснований.
Правила Чаргаффа.
Чаргаффом был установлен ряд закономерностей соотношения нуклеотидов в нуклеиновых кислотах.
1.Количество пуриновых оснований равно количеству пиримидиновых, т.е. А+Г=T+Ц и отношение А+Г/Т+Ц=1.
2.Количество аденина равно количеству тимина А=Т, а количество гуанина равно количеству цитозина Г=Ц и отношения А/Т=1,
Г/Ц=1.
3.Количество оснований, содержащих аминогруппу в 4-ом положении пиримидинового кольца и 6-ом положении пуринового коль-
272

ца, равно количеству оснований, содержащих в этих же положениях кето-группу.
Опираясь на правила Чаргаффа и рентгеноструктурные исследования, проведенные Франклином и Уилкинсоном, Уотсон и Крик в 1953 г. создали модель строения вторичной структуры ДНК.
Вторичная структура:
Вторичной структурой ДНК называют пространственную организацию полинуклеотидных цепей в ее молекуле.
Вторичная структура может существовать в различных конформационных формах. В настоящее время известно 6 форм: А,В,С,Д,Е и Z. При физиологических условиях преобладающим типом ДНК является B-форма. Она представляет собой 2 антипараллельные полинуклеотидные цепи, образующие правую спираль вокруг одной общей оси.
Обе цепи удерживаются между собой водородными связями, возникающими между комплементарными азотистыми основаниями:
|
HNH |
|
O |
|
O |
|
HNH |
|
|
|
|
|
|
||
N |
N |
HN |
|
CH3 |
N |
NH |
N |
|
|
HN |
|
|
|||
HN |
|
|
|
|
NH |
O N |
|
N |
O N |
|
N |
||||
|
|
|
|
H |
|||
|
A T |
H |
|
|
Á |
Ö |
|
|
|
|
|
||||
Сахарно-фосфатные остатки обеих цепей обращены наружу и
образуют остов спирали. За счет остатка фосфорной кислоты они имеют отрицательный заряд. Азотистые основания ориентированы внутрь спирали и располагаются параллельно друг другу в виде «монетных столбиков». Гидрофобные взаимодействия между азотистыми основаниями (стекинг-взаимодействие) удерживают структуру двойной спи-
рали. Таким образом, стабилизация вторичной структуры ДНК обеспечивается: 1) водородными связями, 2) гидрофобным взаимодействием.
На один шаг спирали приходится примерно 10 пар оснований. Шаг спирали равен 3,4 нм (10 0,34=3,4). Диаметр двойной спирали равен примерно 2 нм. Спираль имеет «большой» и «малый» желобки. Они отграничиваются отрицательно заряженными остатками фосфорной кислоты.
Третичная структура ДНК. Компактизация ДНК, хромосомы.
Третичной структурой ДНК называют пространственное расположение полинуклеотидных цепей в их взаимосвязи с гистоновыми белками.
Если всю ДНК, содержащуюся в ядре клетки, вытянуть в одну
273

линию, то ее длина составит 1,74 м. Такой размер ДНК не соизмерим с размерами ядра. Поэтому ДНК у эукариотических клеток находятся в ядре в виде хромосом. Хромосомы – это конденсированные структурные комплексы ДНК и белков. Каждая хромосома образуется из одной длинной молекулы ДНК и содержит от 50 106 до 250 106 нуклеотид-
ных пар. Вся генетическая информация, хранящаяся в хромосомах, называется геномом. Основной структурной единицей хромосом является нуклеосома. Нуклеосома состоит из 8 молекул гистонов – по 2 молекулы Н2а, Н2b, Н3, Н4. Эти гистоны образуют белковое ядро, на которое
накручивается фрагмент молекулы двухцепочечной ДНК. В интактном хроматине ДНК тянется в виде непрерывной нити от нуклеосомы к нуклеосоме.
2Н2а |
2Н3 |
|
|
2Н2b |
2Н |
|
Линкерная последовательность |
Нуклеосомная последовательность в виде «четок» или «бус» |
|
Каждая нуклеосома соединена с соседней линкерной последовательностью. Длина такой последовательности от 0 до 80 нуклеотид-
ных пар.
Молекула ДНК оборачивается вокруг белкового ядра, делая 13/4
оборота, причем на каждый оборот приходится 83 пары нуклеотидов. В среднем, последовательность нуклеосома + линкерная часть повторяется через 200 нуклеотидов. Образование нуклеосомной последовательности способствует компактизации молекулы ДНК примерно в 7 раз. Дальнейшая упаковка молекулы ДНК происходит путем скручивания нуклеосомной последовательности в виде спирали или «соленоида» диаметром 30 нм. При этом достигается компактизация еще примерно в 6 раз. Считают, что этому процессу способствуют гистоновые белки Н1.
Нуклеосомная структура в виде «четок» или «бус» многократно скручивается в виде спирали. Образуется структура диаметром
30 нм.
Каждая м екула гистона Н1 связывается глобулярной частью с определенным местом на нуклеосоме, а C- и N-концевые аминокис-
274

лотные последовательности присоединяются к соседним нуклеосомам. При этом нуклеосомы стягиваются вместе и образуют регулярную, повторяющуюся структуру. Области ак-
тивного хроматина (участки ДНК, в ко- |
NH |
OOH |
торых происходит считывание генети- |
|
|
ческой информации) обладают низкой |
|
|
способностью связывать Н1 гистон и, |
|
|
поэтому, они менее конденсированы и |
|
|
доступны для считывания информации. В дальнейшем формируются нити хроматина диаметром около 100 нм, как располагаются нуклеосомы в этой структуре не выяснено. Упаковка ДНК в хроматиновой
рилла, |
фибрилле позволяет уменьшить |
|
ее линейные размеры |
до 1 мм |
|
нутая в |
(компактиза-ция в 400 раз), а |
|
виде петель |
||
|
диаметр ядра обычно не превы- |
|
|
шает 5 мкм. Поэтому должны |
|
Д |
существовать и другие, более |
|
высокие уровни компактизации. |
||
|
Считают, что далее |
фибрилла |
укладывается в виде петель за счет ДНК-связывающих белков, кото-
рые сближают и удерживают определенные части фибрилл. Типичная хромосома человека может содержать до 2000 таких петель. В последующем, образовавшиеся петлеобразные структуры многократно скручиваются и упаковываются в хромосому.
Активный хроматин составляет в разных клетках 2-11%. Таким
образом, основная часть хроматина клетки не активна. Компактизация ДНК обусловливает избирательную активность участков ДНК, которые обеспечивают синтез белков, специфичных для метаболизма данной клетки.
2.2.Нуклеотидный состав и структура РНК
Вотличие от ДНК РНК построена из 4 основных типов рибонуклеотидов: АМФ, ГМФ, ЦМФ, УМФ.
Известны 5 типов РНК:
1)Информационная РНК (и-РНК или м-РНК (mesenger – по-
средник)).
2)Транспортная РНК (т-РНК).
3)Рибосомная РНК (р-РНК).
4)Гетерогенные ядерные РНК (гигантские или гяРНК).
5)Малые ядерные РНК (мяРНК).
Информационная РНК (и-РНК) Синтезируется в ядре из гетеро-
генных ядерных РНК. Является переносчиком генетической информа-
275

ции от ДНК к белку. На 3’-конце имеет полиаденилатную последовательность (около 200 нуклеотидов), на 5’-конце находится 7-
метилгуанин («кэп» – участок).
Транспортная РНК (т-РНК) состоит примерно из 75 нуклеоти-
дов. Имеет молекулярную массу около 25000Да. Вторичная структура по форме напоминает «Клеверный лист». Р. Холли в связи с этим дал название вторичной структуре – структура «клеверного листа». Структура имеет 4 стебля. Каждый стебель состоит из двух антипараллельных цепей, удерживающихся водородными связями между комплементарными азотистыми основаниями. Стебли имеют форму правой двой-
ной спирали, на 1 виток приходится 11 |
Антикодоновая |
|
пар оснований, шаг спирали равен 3,1 нм. |
петля |
|
Акцепторный стебель на 3’-конце |
|
|
имеет ЦЦА последовательность. Эта по- |
Псевдоуриди- |
|
следовательность необходима для присое- |
ловая петля |
|
динения к т-РНК соответствующей ами- |
игидроуриди- |
|
нокислоты. Характеристика фермента, |
ловая петля |
|
присоединяющего аминокислоту, и меха- |
Акцепторный |
|
низм его действия будут рассмотрены |
стебель |
|
позже. |
Ц |
|
А |
||
Антикодоновый стебель заканчива- |
||
|
ется антикодоновой петлей. Она содержит специфичный для каждой т-
РНК триплет нуклеотидов (антикодон). Антикодон специфически соединяется с соответствующим кодоном и-РНК.
Псевдоуридиловый стебель заканчивается псевдоуридиловой петлей. Название связано с наличием в составе этой петли минорного азотистого основания – псевдоуридина. Петля используется для присоединения т-РНК к рибосоме.
Дигидроуридиловый стебель – заканчивается дигидроуридиловой петлей (в состав входит минорное азотистое основание дигидроурацид). Эта петля необходима для связывания с ферментом аминоацил-т-
РНКсинтетазой (присоединяет к тРНК соответствующую ей аминокислоту). Иногда могут встречаться и добавочные петли. В третичной структуре молекула т-РНК напоминает букву Г. Акцепторный и псев-
доуридиловый стебли образуют «перекладину» буквы Г, а антикодоновый и дигидроуридиловый стебли формируют «ножку».
Рибосомная РНК (р-РНК) вместе с рибосомными белками участ-
вует в формировании рибосом. В рибосомах эукариот на долю РНК приходится около 50% их массы. р-РНК обладают различной молеку-
лярной массой и при ультрацентрифугировании имеют различные константы седиментации (S – константа Сведберга). Все р-РНК классифицируют по константе седиментации. У эукариот р-РНК представлена в большой субъединице рибосом 28S РНК (400 нуклеотидов), 5S (121
276
нуклеотид), 5,8S РНК (155 нуклеотидов). Малая субъединица рибосом имеет в своем составе одну молекулу р-РНК с константой седимента-
ции 18S (около 200 нуклеотидов). Третичная структура РНК является скелетом рибосомы. Имеет форму палочки или клубочка. Снаружи располагаются рибосомальные белки. Определенные участки РНК могут играть важную роль в прикреплении и-РНК к рибосомам.
Гетерогенные ядерные РНК (гигантские РНК, гяРНК) Являются предшественниками РНК. Синтезируются на ДНК и являются их точной копией, несущей избыточную информацию, которая удаляется в процессе созревания и превращения в соответствующую и-РНК.
Малые ядерные РНК (мяРНК) Состоят из 90-220 нуклеотидов. В
настоящее время известно более 30 типов мяРНК. Биологическая роль до конца не изучена. Однако, известно, что некоторые мяРНК участвуют в процессе созревания и-РНК (и сплайсинге). Обнаружено, что
эти РНК проявляют ферментативную активность, нарушая, таким образом, монополию белков в отношении катализа.
2.3. Видовые различия первичной структуры нуклеиновых кислот
По нуклеотидному составу ДНК можно определить видоспецифичность организмов (геносистематика по Белозерскому). Так, коэффициент специфичности (Г+Ц/А+Т) у эукариот меньше 1, а у прокариот чаще больше или равен 1. При одинаковом нуклеотидном составе, строение конкретной ДНК зависит от последовательности нуклеотидов. Для РНК коэффициент специфичности чаще превышает 1. Считают, что по нуклеотидному составу ДНК можно, в перспективе, построить все родословное дерево живого мира.
3. Физико-химические свойства нуклеиновых кислот
Подобно белкам нуклеиновые кислоты могут денатурировать. Денатурация нуклеиновых кислот состоит в расхождении цепей двойной спирали ДНК и двуспиральных участков РНК. В качестве денатурирующих агентов могут быть кислоты, щелочи, спирты... В результате денатурации каждая из цепей нуклеиновых кислот приобретает форму беспорядочного клубка. Азотистые основания имеют максимум светопоглощения при длине волны 260 нм. При денатурации нуклеиновых кислот их оптическая плотность увеличивается примерно на 40% в сравнении с нативной. Такое явление получило название гипер-
хромного эффекта. Поэтому о денатурации нуклеиновых кислот судят по увеличению оптической плотности их растворов при 260 нм. При нагревании гиперхромный эффект отмечается в узком диапазоне тем-
277
ператур. Этот диапазон получил название точки плавления. Он равен
80-85оС.
Если устранить денатурирующий агент, то структура ДНК может быть восстановлена за счет взаимодействия комплементарных азотистых оснований. Этот процесс получил название ренативации. На явлении ренативации основан метод гибридизации ДНК. Этот метод применяется при исследовании нуклеиновых кислот, полученных из разных источников.
Обмен нуклеотидов
1.Распад нуклеиновых кислот
1.1.Краткая характеристика и классификация нуклеаз
Нуклеазами называют обширную группу ферментов, специфически действующих на нуклеиновые кислоты, разрушая в них фосфодиэфирные связи с высвобождением полинуклеотидных фрагментов различной длины и отдельных нуклеотидов, без выделения неорганического фосфата.
По месту действия нуклеазы подразделяют на эндонуклеазы и экзонуклеазы.
Эндонуклеазы – разрушают сложноэфирные связи в различных местах внутри молекул нуклеиновых кислот. Ряд эндонуклеаз обладают чрезвычайно высокой специфичностью к строго определенным последовательностям нуклеотидов и действуют только на определенные связи в этой последовательности. Такие эндонуклеазы называют эндо-
нуклеазами рестрикции или рестриктазами. Способность рестриктаз образовывать высокоспецифичные фрагменты широко используется в настоящее время для клонирования ДНК и генной инженерии, в криминалистике. Эндонуклеазы могут быть специфичными к одному из 2-
х типов нуклеиновых кислот, и в соответствии с этим называются ри-
бонуклеазами (РНК-азы) и дезоксирибонуклеазами (ДНК-азы).
Неспецифические эндонуклеазы – способны атаковать как ДНК,
так и РНК.
Экзонуклеазы – способны последовательно отщеплять от одного из концов полинуклеотидной цепи по нуклеотиду. Существует небольшая группа эндонуклеаз, обладающих строгой специфичностью к ДНК или РНК. Однако большинство способно атаковать как ДНК, так и РНК. Как и в случае с ДНКазами I и II, некоторые РНКазы способны образовывать в ходе гидролиза РНК 3’-фосфорпроизводные (эндонуклеазы селезенки) и 5’-фосфорпроизводные (ферменты змеиного яда). Разрыв фосфо-эфирной связи происходит между фосфором и кислоро-
дом:
278

OH
O P
P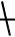 O-
O-
R
1.2. Нуклеазы пищеварительного тракта
Нуклеиновые кислоты поступают с пищей, главным образом, в составе сложных белков – нуклеопротеидов. Белковая часть разрушается в желудочно-кишечном тракте протеолитическими ферментами до
отдельных аминокислот. В результате высвобождается свободная нуклеиновая кислота, которая подвергается действию нуклеаз с образованием нуклеотидов. Нуклеотиды разрушаются нуклеотидазами до нуклеозида и неорганического фосфата. Нуклеозиды способны всасываться в кишечнике или разрушаться нуклеозидазами до пуриновых или пиримидиновых азотистых оснований и рибозы или дезоксирибозы. Пуриновые основания могут окисляться до мочевой кислоты, которая всасывается в кишечнике и затем фильтруется с мочой через почки. Переваривание белковой части нуклеопротеидов и нуклеиновых кислот тесно связаны и дополняют друг друга. Так, панкреатическая ДНК-аза действует более эффективно только в комплексе с протеоли-
тическими ферментами. При гидролизе белковой части нуклеопротеидов высвобождаются лизин, аргинин и гистидин (преобладают в составе гистонов), которые активируют ДНК-азу. Другие аминокислоты не
обладают способностью активировать этот фермент.
Пищевые нуклеотиды, полученные в результате переваривания нуклеопротеидов, практически не используются для построения нуклеиновых кислот, хотя введенные парентерально нуклеотиды способны включаться в нуклеиновые кислоты (причина такой избирательности до конца не выяснена).
1.3. Нуклеазы тканей
Во внутренней среде организма (клетки, межклеточное пространство, кровь), а так же на границах с окружающей средой (эпителий кожи и слизистых оболочек) определяется высокая активность нуклеаз. В результате действия этих ферментов, как и в предыдущих случаях, образуются нуклеотиды, затем нуклеозиды и, наконец, азотистые основания и пентозы. Функции этих ферментов, в основном, можно свести к следующим:
1) Защита от внедрения чужеродного генетического материала.
279
Эктоферменты – рибонуклеазы кожи разрушают рибонуклеиновые кислоты на кожной поверхности.
2)Поддержание постоянства собственного генетического аппарата (репарация).
ДНК-репарирующие нуклеазы распознают участки поврежден-
ной ДНК, осуществляют гидролиз фосфодиэфирных связей между поврежденными нуклеотидами и остальной частью молекулы ДНК, в результате происходит удаление поврежденных участков.
3)Гомеостатическая.
Регулируя активность РНК-аз, можно регулировать процессы
биосинтеза и деградации нуклеиновых кислот и белков. Так, активность нуклеотидаз контролируется концентрацией нуклеотидов. Баланс между активностью нуклеотидаз и нуклеозидаз обеспечивает поддержание уровня нуклеотидов, используемых для биосинтеза нуклеиновых кислот. и-РНК, использованные для биосинтеза белка, под-
вергаются действию нуклеаз и разрушаются. В результате прекращается биосинтез белка. Ряд предшественников РНК созревают при участии нуклеаз. Предшественник тирозиновой т-РНК E.Coly содержит 129 нуклеотидов. РНК-азы отщепляют 44 нуклеотида и превращают предшественник в зрелую т-РНК.
1.4. Распад пуриновых нуклеотидов
Конечными продуктом распада пуринов человека является моче-
вая кислота.
У многих животных, в том числе и у ряда млекопитающих, мочевая кислота разрушается до алантоина ферментом уриказа. Однако у человека этот фермент отсутствует. Считают, что распад пуринов до мочевой кислоты обусловлен невозможностью синтеза аскорбиновой кислоты.
Урат, в какой-то мере, способен заменять аскорбиновую кислоту,
поскольку обладает антиоксидантной активностью. Как показано на рисунке, аденин может быть гидролилитически дезаминирован в гипоксантин, а гуанин – в ксантин. Фермент ксантиноксидаза окисляет далее гипоксантин и ксантин в мочевую кислоту (в некоторых растениях ксантин при его метилировании превращается в кофеин). Ксантиноксидаза – это флавиновый фермент, относится к группе флавопротеидов (в его состав входит производное витамина В2 –рибофлавин).
Молекула фермента является димером и содержит 2 молекулы ФАД, 2 атома молибдена и 8 атомов железа.
280

Схема распада пуриновых нуклеотидов
|
|
|
NH 2 |
|
|
|
|
|
O |
|
|
N |
N |
|
|
|
|
N |
NH |
CH |
OH |
N |
N |
|
CH |
OH |
N |
NH 2 |
|
2 |
O |
|
енозин- |
|
2 |
O |
|
||
|
|
|
|
|
|
||||
|
|
Н2О |
заминаза |
|
|
|
Pi |
||
OH |
OH |
|
|
|
OH |
OH |
Рибозо-1- |
||
Аденозин N |
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
фосфат |
|
|
|
|
|
|
|
|
|
O |
|
|
N |
|
|
|
|
|
|
NH |
CH 2OH |
N |
|
|
|
|
|
|
||
|
|
|
|
|
|
NH 2 |
|||
|
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Гуанин |
OH |
OHPi |
|
О2 |
|
|
|
|
Н3 |
|
Инозин |
|
|
|
|
HКсантин |
|
|||
Рибозо-1-фосфат |
O |
Ксантин- |
|
|
|||||
|
|
|
|
|
О2 |
|
|
||
|
|
N |
|
оксидаза |
|
|
|
||
|
|
|
NH |
|
|
|
|
|
|
|
|
HN |
N |
|
H |
|
|
|
|
|
|
|
|
N |
|
|
NH |
|
|
|
|
Гипоксантин |
O |
|
|
|
|||
|
|
HN |
|
|
O |
|
|||
|
|
|
|
|
|
N |
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
H |
|
|
|
|
|
|
|
Мочевая кислота |
|
|||
Н – обозначает кислотный протон
РКа =5,4
Пути регенерации пуриновых нуклеотидов
Существует 2 основных механизма регенерации:
1) Фосфорибозилирование свободных пуриновых оснований.
Осуществляется двумя ферментами:
а) аденинфосфорибозилтрансфераза – переносит фосфорибо-
зилпирофосфат на аденин с образованием АМФ.
б) более активный – гипоксантин-гуанинфосфорибозилтрансфе-
раза – фосфорилирует ксантин и гуанин до ИМФ и ГМФ.
2) Фосфорилирование пуриновых нуклеозидов. Катализируется
аденозинкиназой.
281