

Учреждение образования
”ПОЛЕССКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ“
Факультет биотехнологический
Кафедра биохимии и биоинформатики
Управляемая самостоятельная работа № 1
на тему:
Биосинтез нуклеиновых кислот (репликация и транскрипция)
Подготовил |
|
Децук Валерия Петровна |
|||
Студент 2 курса, гр.22БХ-1 |
(подпись) __________________2023 |
||||
|
|
|
|
||
Проверил |
|
Аль Меселмани Моханад Али |
Доцент |
(подпись) ___________________2023 |
|
ПИНСК 2023
ОГЛАВЛЕНИЕ
Глава 1. Синтез днк (Репликация) 3
Глава 2. Синтез рнк (Трансляция) 9
СПИСОК ИСПОЛЬЗУЕМОЙ ЛИТЕРАТУРЫ 13
Глава 1. Синтез днк (Репликация)
Молекула ДНК состоит из двух полинуклеотидных цепей, объединённых в единую молекулу за счёт многочисленных водородных связей, возникающих между комплементарными азотистыми основаниями сближенных в пространстве мононуклеотидов (уотсон-криковские пары)
ДНК локализуется в ядре клеток в комплексе с определёнными ядерными белками, формируя особую субстанцию - хроматин. Хроматин состоит из 60 % белка, 35 % ДНК и 5 % РНК. В хроматине ДНК, объединяясь с белками гистонами, формируют нуклеосомы.
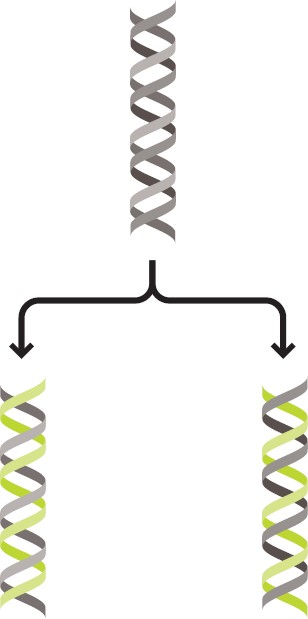 Функция
ДНК заключается в хранении генетической
информации. При деленииклетки каждая
из дочерних клеток получает точную
копию ДНК материнской клетки. Процесс
удвоения ДНК называется репликацией.
Функция
ДНК заключается в хранении генетической
информации. При деленииклетки каждая
из дочерних клеток получает точную
копию ДНК материнской клетки. Процесс
удвоения ДНК называется репликацией.
Рисунок 1 – Репликация ДНК
Для того чтобы произошел процесс репликации, необходимо, чтобы отдельные нити молекулы ДНК расплелись и на каждой из них, как на матрице, произошло построение комплементарной дочерней полинуклеотидной цепи. Подобный тип репликации получил название «полуконсервативный».
В нем участвует множество различных белков, прежде всего ДНК-полимеразы. Каждая мономерная единица, присоединяющаяся к растущей цепи, находится в форме дезоксирибонуклеозид-5*-трифосфата (дНТФ): фосфатная группа, связанная с 5'-углеродным атомом дезоксирибозы, обозначается буквой а, к ней присоединены β-фосфат и далее - γ-фосфат.
В ходе репликации β- и γ-фосфатные группы отщепляются в виде пирофосфата, a α-фосфатная группа связывается с 3'-ОН-группой последнего нуклеотида растущей цепи.
С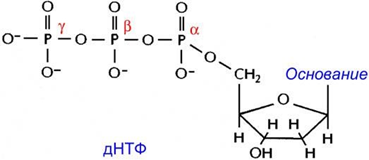 интез
ДНК идет в направлении 5'→3', поскольку
рост цепи ДНК идёт за счёт формирования
фосфодиэфирной связи между 3*-кислородом
растущей цепи и α-фосфатом
дНТФ.
интез
ДНК идет в направлении 5'→3', поскольку
рост цепи ДНК идёт за счёт формирования
фосфодиэфирной связи между 3*-кислородом
растущей цепи и α-фосфатом
дНТФ.
Каждая из цепей ДНК служит матрицей для синтеза комплементарной цепи. Комплементарность оснований противоположных цепей гарантирует идентичность новосинтезированной и исходной ДНК.
Синтез дочерней молекулы ДНК осуществляется ферментом ДНК-зависимая ДНК-полимераза (или просто ДНК-полимераза). Этот фермент представляет олигомерный белок - металлофермент. В состав его активного центра входят два иона магния Mg2+, электростатически удерживаемые боковыми радикалами аспарагиновых кислот.
Ф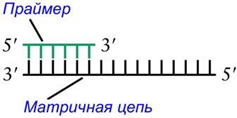 ермент
ДНК-полимераза не может самостоятельно
инициировать новый (de novo) синтез цепи.
Для этого ей нужно, чтобы уже существовала
цель или ДНК, или РНК, которая называется
праймер.
ермент
ДНК-полимераза не может самостоятельно
инициировать новый (de novo) синтез цепи.
Для этого ей нужно, чтобы уже существовала
цель или ДНК, или РНК, которая называется
праймер.
Праймер синтезирует фермент ДНК-праймаза (DNA primase). Когда уже есть праймер, спаренный с матричной нитью ДНК, тогда ДНК-полимераза катализирует реакцию присоединения к гидроксильной группе на 3'-конце праймера того дНТФ, который закодирован в матричной цепи.
Для того чтобы дуплекс ДНК
мог работать как матрица в процессе
репликации, две его нити должны быть
расплетены (или ещё говорят, расплавлены,
melted), чтобы обеспечить доступ к нуклеиновым
основаниям тех дНТФ, из которых
ДНК-полимераза формирует дочернюю нить.
Это расплетение, используя знергию
гидролиза АТФ, производят ферменты
геликазы (helicase). Расплетение начитнается
со специфических последовательностей
в ДНК, которые называются началом
репликации (replication origin) или просто
ориджином (ог). Последовательности
нуклеотидов в ориджинах разных организмов
сильно варьируются, хотя они имеют и
общее свойство - в ориджинах повышенная
концентрация А-Т пар. В таких областях
меньше водородных связей между нитями
ДНК, и поэтому «легче» осуществлять
«плавление» (расплетение) Д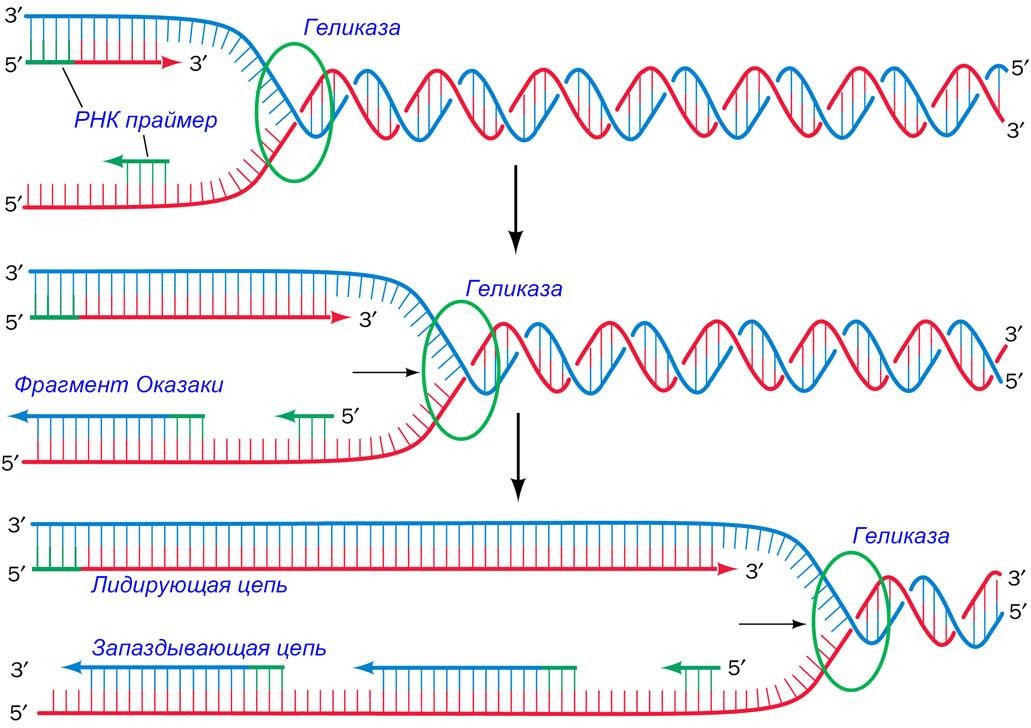 НК.
НК.
Как только геликаза расплетёт дуплекс, фермент праймаза (специализированная РНК-полимераза) синтезирует короткий РНК-праймер, комплементарный расплетённому участку матричной цепи. Спаренный с ДНК праймер затем удлиняется ДНК- полимеразой, формируя дочернюю цепь.
Область ДНК, в которой собраны все эти ферменты для синтеза дочерних цепей, называется репликационная вилка (replication fork) или вилка роста.
В ходе репликации репликационная вилка и ассоциированные с нею белки удаляются от ориджина, что сопровождается ростом торсионных напряжений в ДНК.
Эти напряжения снимаются ферментом топоизомераза (topoisomerase).
Для того чтобы процесс репликации протекал непрерывно и ДНК-полимераза перемещалась и копировала дуплекс ДНК, геликаза должна непрерывно расплетать дуплекс, а топоизомераза должна удалять образующиеся суперспирали.
Главная проблема при «обработке» ДНК репликационной вилкой вытекает из двух факторов:
две нити родительской ДНК антипараллельны;
ДНК-полимераза катализирует рост дочерней цепи только в направлении 5'3'.
Синтез дочерней цепи, которая называется лидирующая цепь (leading strand), может происходить непрерывно, начиная с единственного праймера в 5'3' направлении, то есть в том направлении, в котором перемещается репликационная вилка.
Проблемы возникают при синтезе другой дочерней цепи, которая называется запаздывающей (lagging strand).
Поскольку рост запаздывающей цепи должен происходить в 5'3' направлении, копирование матрицы должно осуществляться в направлении противоположном движению репликационной вилки.
 Клетка
решает эту проблему, синтезируя новые
праймеры, через каждые несколько сотен
оснований на матричной цепи, по мере
того, как дуплекс расплетается, и всё
5' более оснований освобождаются при
распределении.
Клетка
решает эту проблему, синтезируя новые
праймеры, через каждые несколько сотен
оснований на матричной цепи, по мере
того, как дуплекс расплетается, и всё
5' более оснований освобождаются при
распределении.
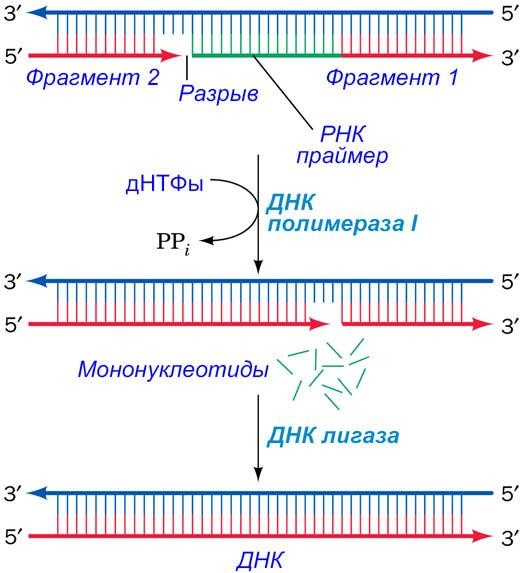
Каждый из этих праймеров, спаренный с матричной цепью, удлиняется в 5'→3' направлении, образуя непрерывные сегменты, которые называются фрагменты Оказаки).
РНК-праймер каждого фрагмента Оказаки удаляется и замещается цепью соседнего растущего фрагмента Оказаки.
Фермент ДНК-лигаза (DNA (gase) сшивает соседние фрагменты.
После расплетения уже небольшого участка молекулы ДНК, к каждой освободившейся полинуклеотидной цепи присоединяются молекулы ДНК-связывающего белка (single-stranded-binding protein, SSB), препятствующего обратному соединению комплементарных пар азотистых оснований.
ДНК-полимераза надёжно зафиксирована на нити ДНК белковым скользящим захватом (siding clamp).
Р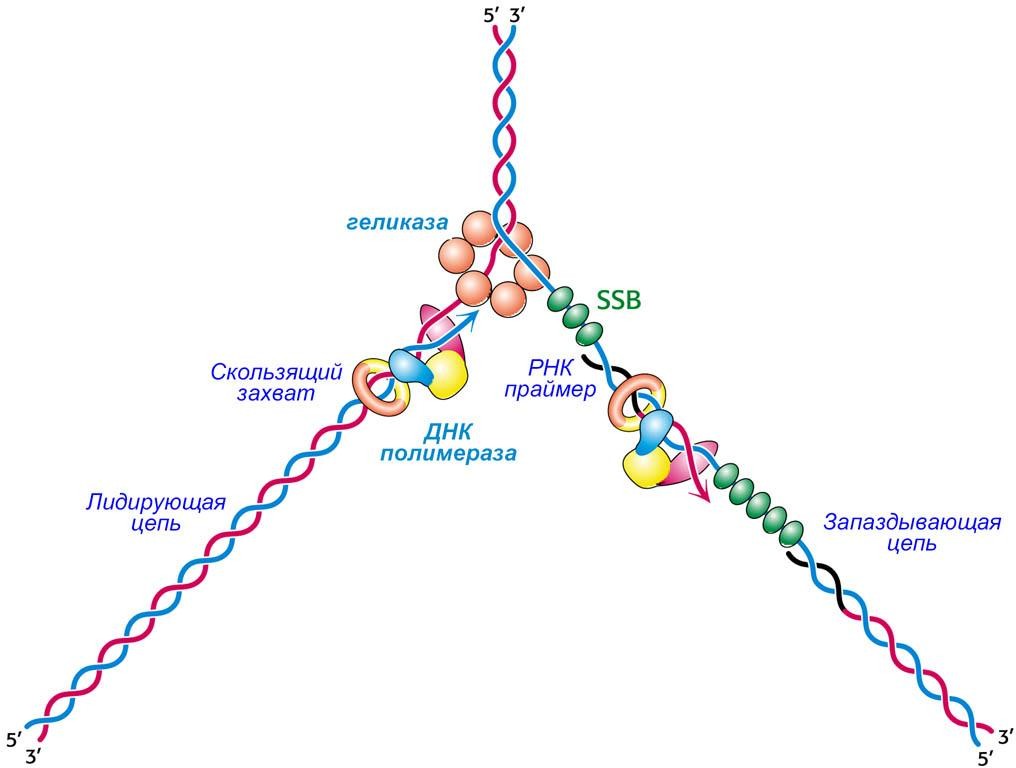 асплетение
цепей ДНК идёт с очень большой скоростью.
Экспериментально установлено, что
угловая скорость вращения расплетающихся
полинуклеотидных цепей близка к 4500
об/мин. Настолько большая скорость
создаёт реальную угрозу для механического
повреждения хрупкой молекулы ДНК Защита
ДНК и всей хромосомы от высокой скорости
вращения обеспечивается ферментом
молеизомераза. Этот фермент производит
временный разрыв одной из полинуклеотидных
цепей в непосредственной близости от
участка расплетения, после чего вновь
сшивает ее.
асплетение
цепей ДНК идёт с очень большой скоростью.
Экспериментально установлено, что
угловая скорость вращения расплетающихся
полинуклеотидных цепей близка к 4500
об/мин. Настолько большая скорость
создаёт реальную угрозу для механического
повреждения хрупкой молекулы ДНК Защита
ДНК и всей хромосомы от высокой скорости
вращения обеспечивается ферментом
молеизомераза. Этот фермент производит
временный разрыв одной из полинуклеотидных
цепей в непосредственной близости от
участка расплетения, после чего вновь
сшивает ее.
В клетках существует несколько разновидностей ДНК-полимераз, хотя не все они принимают участие в удвоении ДНК.
Помимо этих ферментов, в репликации участвуют ещё более двух десятков белковых факторов, которые объединяются в единую ДНК-репликазную систему.
Процесс репликации характеризуется высокой степенью точности матричного синтеза. В большей мере это обусловлено многофункциональностью ДНК-полимеразы.
Этот фермент не только обеспечивает рост дочерней полинуклеотидной цепи, но и «проверяет» правильность встраивания в неё мононуклеотидов.
ДНК-полимераза «обнаруживает» неправильно встроенные мононуклеотиды, вырезает их и встраивает на их место новые, «правильные» мононуклеотиды. Благодаря этому, частота появления ошибок в процессе синтеза дочерней полинуклеотидной цепи составляет не более чем 1 на 1010 нуклеотидных пар, что обеспечивает высокую точность копирования генетической информации, а значит и возможность сохранения генетического постоянства вида.