

Полезные материалы за все 6 курсов / Ответы к занятиям, экзаменам / 2. Кора больших полушарий
.pdfТаким образом, теменная кора приматов содержит нейроны, которые специфически реагируют, когда животное обращает внимание на поведенчески значимый стимул, и сила реакции отражает количество внимания, уделяемого стимулу.
Избирательная активация нейронов в теменной коре головного мозга обезьяны как функция внимания. В этом случае внимание макаки-резуса направлено на свет, связанный с вознаграждением за фруктовый сок. (А) Хотя базовый уровень активности исследуемого здесь нейрона остается неизменным, когда обезьяна игнорирует визуальную цель (слева),частота срабатывания резко возрастает, когда обезьяна реагирует на тот же стимул (справа). Гистограммы показывают частоту потенциала действия в единицу времени. (Б) Когда обезьяне предоставляется выбор места для посещения, она уделяет все больше внимания конкретной визуальной цели, когда за это можно ожидать большего вознаграждения фруктовым соком (слева), и соответственно увеличивается частота срабатывания исследуемого теменного нейрона
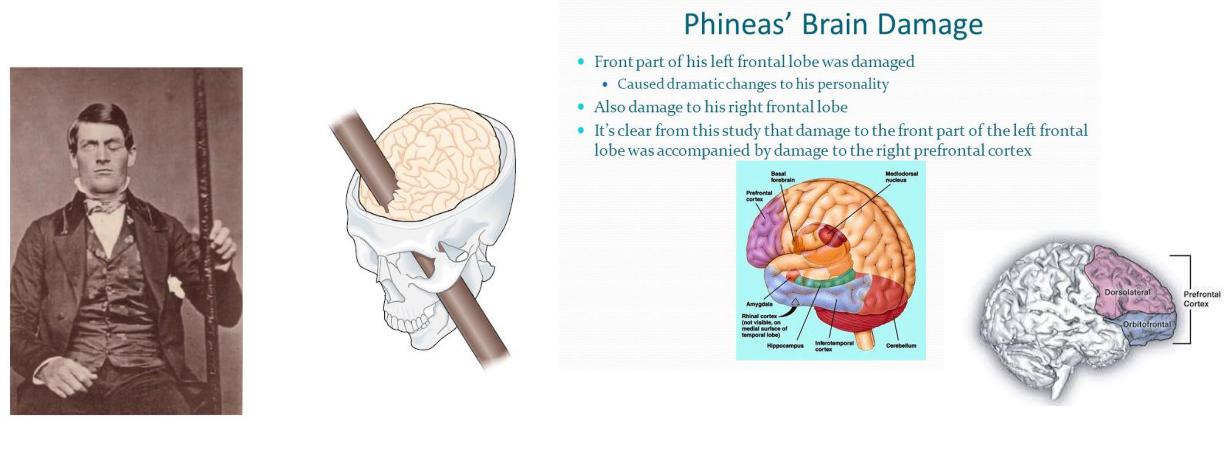
Функции кратковременной памяти расположены более дорсолатерально, а функции планирования и социального сдерживания расположены более вентромедиально.
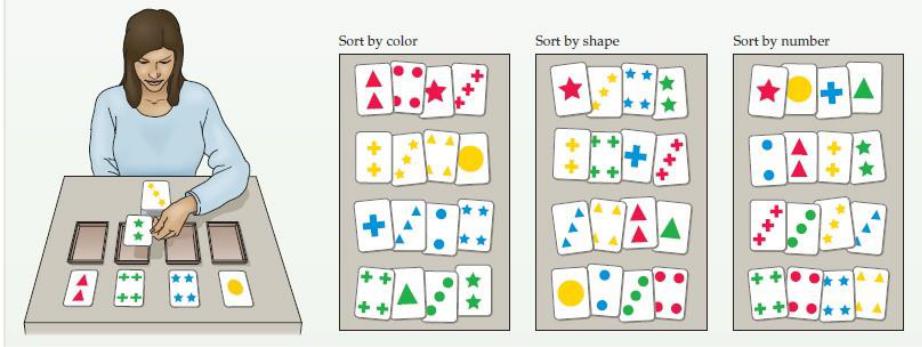
В этом тесте экзаменатор кладет четыре карточки с символами, которые отличаются по количеству, форме или цвету, перед испытуемым, которому дается набор карточек с ответами с похожими символами на них. Затем испытуемого просят поместить соответствующую карточку ответа перед карточкой стимула на основе правила сортировки, установленного, ноне заявленного экзаменатором (т.е. сортировать по цвету, номеру или форме). Затем экзаменатор указывает, является ли ответ “правильным” или “неправильным”. После десяти последовательных правильных ответов экзаменатор изменяет правило сортировки, просто говоря “неправильно”. Затем субъект должен определить новое правило сортировки и выполнить десять правильных проб. Затем правило сортировки снова изменяется до тех пор, пока не будет завершено шесть циклов. Общепризнанным объяснением чувствительности задачи сортировки карточек в Висконсине к дефициту лобных долей является аспект “планирования” этого теста. Чтобы ответить правильно, испытуемый должен сохранить информацию опредыдущем испытании, которая затем используется для руководства поведением в будущих испытаниях. Обработка такого рода информации характерна для нормальной функции лобной доли
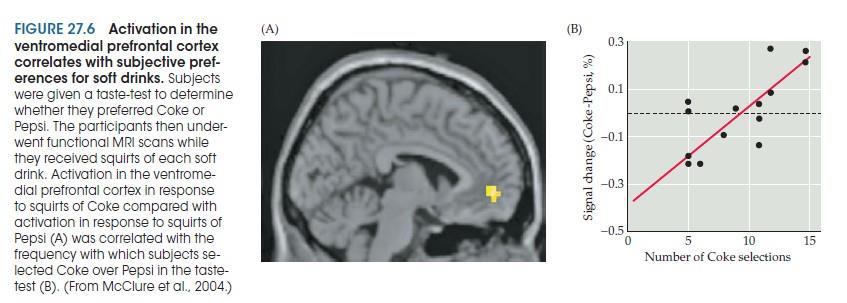
Например, дорсальный и латеральный участки лобной коры активируются, когда нормальные субъекты активно подавляют поведенческую реакцию, когда нарушается ожидаемый паттерн событий, чтобы генерировать соответствующее поведение. Эти результаты подтверждают рольэ той части мозга в сохранении информации в кратковременной памяти и использовании этой информации для подавления автоматических реакций на ожидаемые события. Напротив, личные предпочтения в отношении различных видов вознаграждений коррелируют с индивидуальными различиями в активация мозга в вентромедиальной префронтальной коре, предполагая, что эта часть мозга сигнализирует о ценности, которую люди придают вознаграждению, независимо от его физических характеристик. Такие сигналы имеют решающее значение для принятия адаптивных решений, и влияние нарушенной обработки вознаграждения в вентромедиальной префронтальной коре на принятие решений слишком очевидно в неупорядоченном поведении пациентов (таких как Финеас Гейдж) с повреждением этой области. К сожалению, последствия повреждения лобных долей также были задокументированы многими тысячами лобных лоботомии выполнялись в 1930-1940-х годах как средство лечения психических заболеваний.
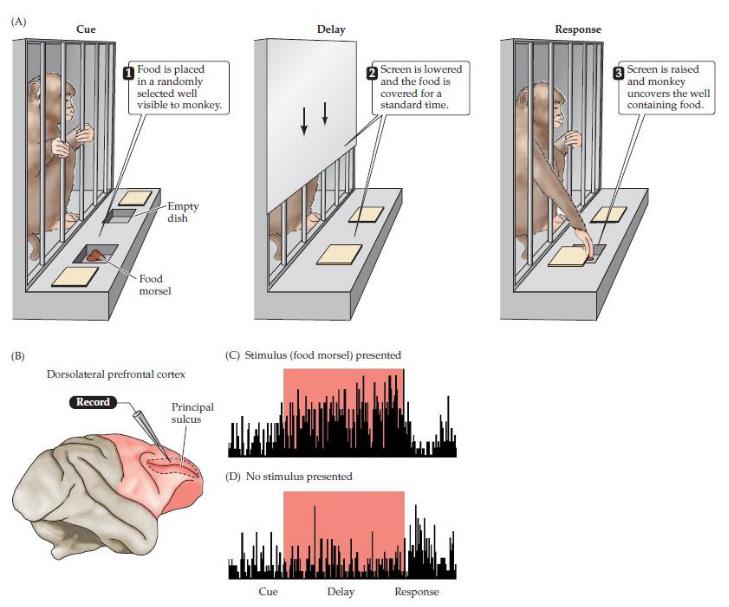
Активация нейронов вблизи главной борозды лобной доли вовремя выполнения задачи с задержкой ответа. (А) Иллюстрация задачи. Экспериментатор случайным образом меняет лунку, в которую помещается пища.
Обезьяна наблюдает, как накрывается кусочек, а затем экран опускается на стандартное время. Когда экран поднят, обезьяне разрешается открыть только один колодец, чтобы достать еду. Нормальные обезьяны быстро усваивают эту задачу, обычно выполняя ее на уровне 90% правильно после менее чем 500 тренировочных испытаний, в то время как обезьяны с лобными поражениями работают плохо.
(B) Регион записи.(C) Активность специфичного к задержке нейрона в префронтальной коре макакирезуса, записанная вовремя задания с задержкой ответа, показанного в (A). Гистограммы показывают количество потенциалов действия в течение периодов сигнала, задержки и ответа. Нейрон начинает работать быстрее, когда экран опущен, и остается активным в течение всего периода задержки. (D) Когда экран опускается и поднимается, но еда не подается, тот же самый нейрон менее активен. (После Голдмана-Ракича, 1987.)
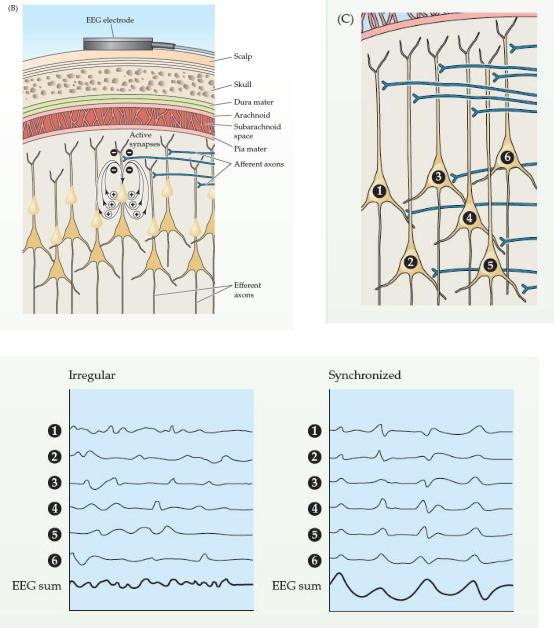
(Б) Электрод на коже головы измеряет активность очень большого количества нейронов в ниже лежащих областях мозга, каждый из которых генерирует небольшое электрическое поле, которое меняется с течением времени. Эта активность (которая, как полагают, в основном синаптическая)делает более поверхностное внеклеточное пространство отрицательным по отношению к более глубоким областям коры. Электрод ЭЭГ измеряет синхронный сигнал, потому что многие тысячи клеток реагируют одинаково в более или менее одно и то же время.
(C) Генерация синхронной активности, которая характеризует глубокий сон. В слое пирамидальных клеток под электродом ЭЭГ каждый нейрон получает тысячи синаптических сигналов. Если входные сигналы нерегулярны или не в фазе, их алгебраическая сумма будет иметь небольшую амплитуду, как это происходит в состоянии бодрствования. Если, однако, нейроны активируются примерно в одно и то же время, то волны ЭЭГ будут иметь тенденцию быть в фазе, а амплитуда будет намного больше, как это происходит в дельта-волнах, которые характеризуют IV стадию сна.
Сонные веретена - это периодические всплески активности примерно от 10 до 12 Гц, которые обычно длятся 1-2 секунды и возникают в результате взаимодействия между таламическими и кортикальными нейронами
В настоящее время известно, что саккадоподобные движения глаз, которые определяют быстрый сон, возникают из-за того, что в отсутствие внешних зрительных стимулов эндогенно генерируются сигналы от понтиновой ретикулярной формации передаются в двигательную область верхних холмиков Колликулярные нейроны проецируются в парамедиальную понтиновую ретикулярную формацию (PPRF)и ростральное интерстициальное ядро, которое координирует время и направление движений глаз.
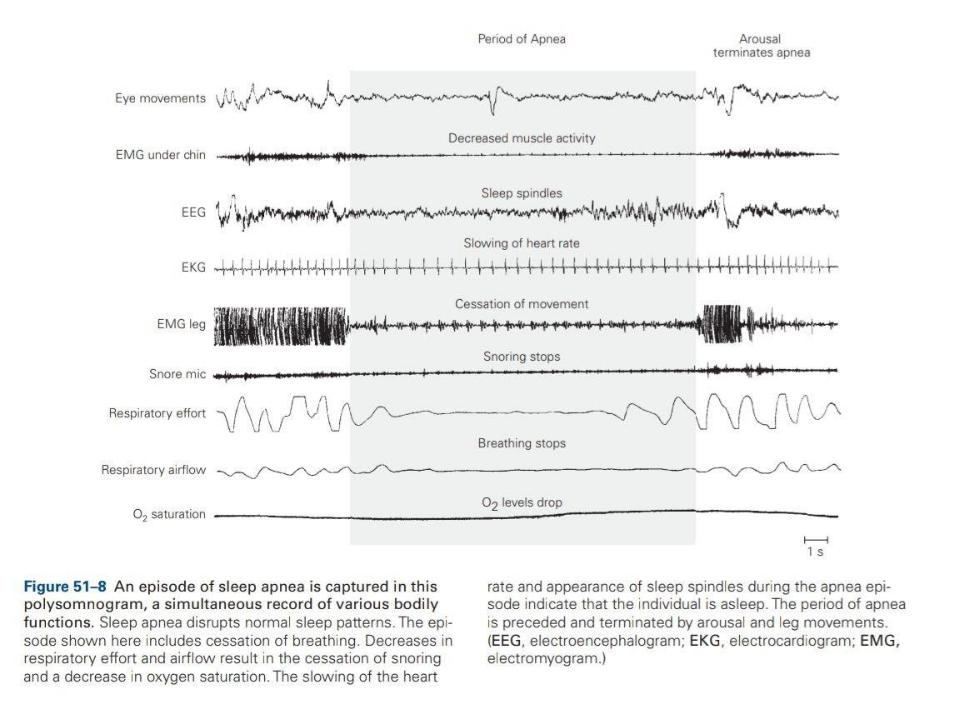
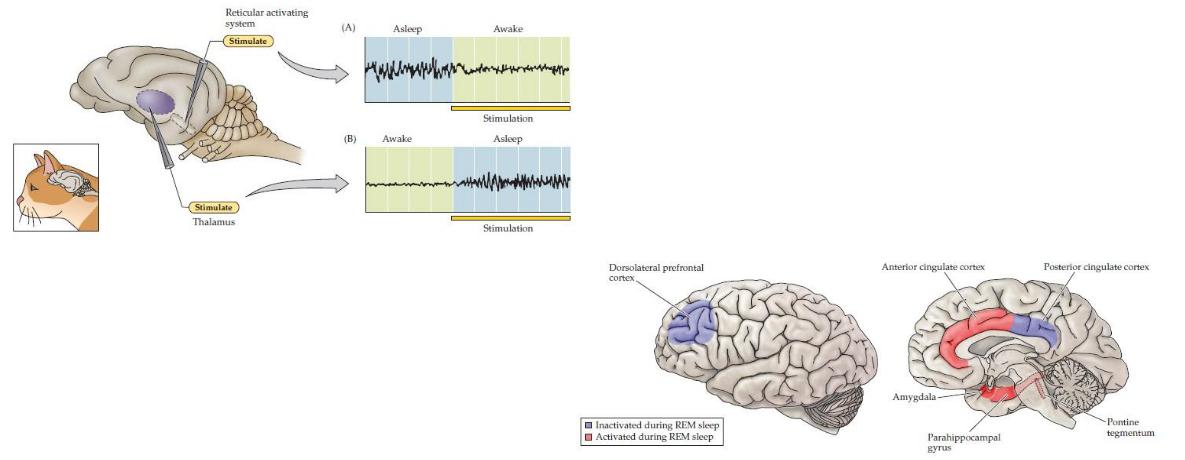
Швейцарский физиолог Вальтер Хесс обнаружил, что стимуляция таламуса у бодрствующей кошки низкочастотными импульсами вызывает медленный сон. Эти ключевые эксперименты показали, что сон влечет за собой закономерное взаимодействие между стволом мозга, таламусом и корой головного мозга. Быстрый сон также характеризуется волнами ЭЭГ, которые возникают в понтиновой ретикулярной формации и распространяются через латеральное коленчатое ядро таламуса к затылочной коре. Эти понтино-геникуло-затылочные волны (PGO) служат полезным маркером начала быстрого сна;
(А) Электрическая стимуляция холинергических нейронов вблизи соединения моста и среднего мозга(ретикулярная активирующая система) вызывает пробуждение спящей кошки. (Б) Медленная электрическая стимуляция таламуса заставляет бодрствующую кошку засыпать. Графики показывают записи ЭЭГ дои во время стимуляции.
Активность в миндалине, парагиппокампе, понтиновом тегментуме и передней поясной коре увеличивается во время быстрого сна, тогда как активность в дорсолатеральной префронтальной и задней поясной коре уменьшается