

Глава 2. Теории кроветворения
унитарная теория (А. А. Максимов, 1909 г.) - все форменные элементы крови развиваются из единого предшественникастволовой клетки;
дуалистическая теория - два источника кроветворения, для миелоидного и лимфоидного;
полифилетическая теория - для каждого форменного элемента свой источник развития.
В настоящее время общепринятой является унитарная теория кроветворения, на основании которой разработана схема кроветворения (И. Л. Чертков и А. И. Воробьев, 1973 г.).
Основоположником современной унитарной теории кроветворения является гистолог Максимов (работал на кафедре гистологии ВМА Санкт Петербурге). Еще в 1907 году Максимов утверждал, что все клетки крови развиваются из единой одной и той же родоначальной клетки; мало того, он назвал эту клетку — морфологически это малый лимфоцит. Однако имеющиеся в то время методы исследований не позволяли экспериментально доказать верность этой теории.
Максимов в ходе гемоцитопоэза клетки крови подразделял на 4 группы: 1 группа — клетки с неограничанной возможностью превращений, т.е. родоначальная клетка, способная развиваться и превратиться в любой форменный элемент крови. 2 группа — клетки с частично ограниченный способностью развиваться в ту или иную форму клеток крови. 3 группа — клетки со строго ограничанной возможностью развития. 4 группа — клетки крови не способные изменяться. Последующие исследования показали верность унитарной теории кроветворения Максимова.
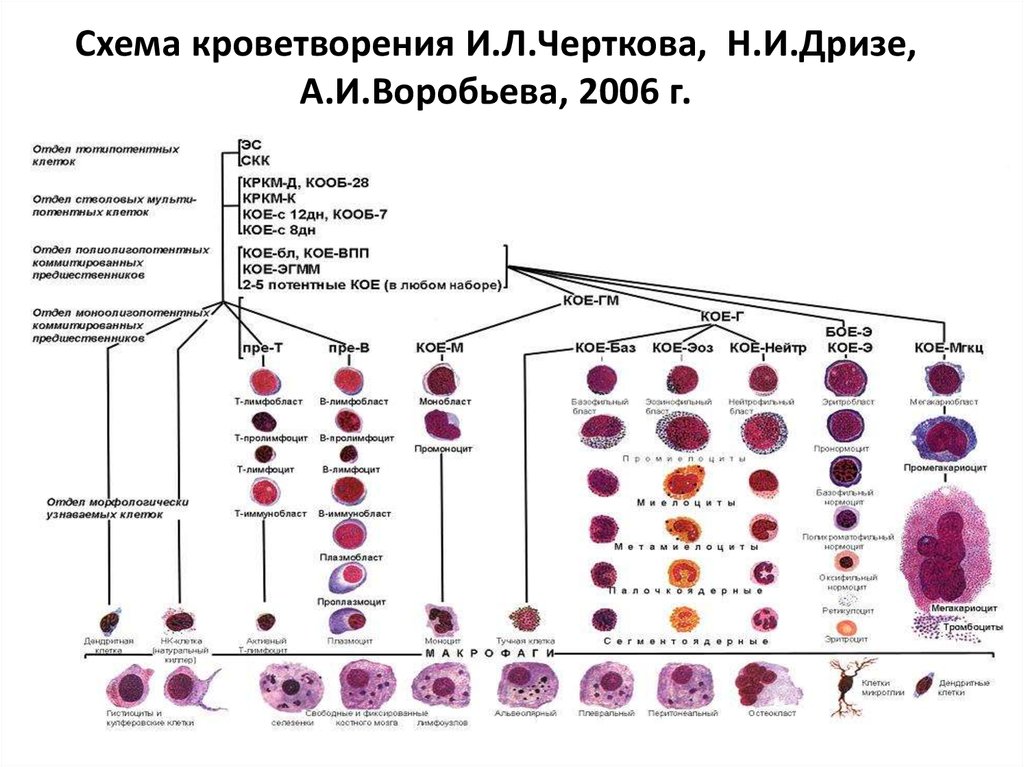
Рисунок 2. – схема кроветворения Воробьева и Черткова
Глава 3. Эмбриональный гемопоэз
Различают эмбриональный гемопоэз, который происходит в эмбриональный период и приводит к развитию крови как ткани, и постэмбриональный гемопоэз, который представляет собой процесс физиологической регенерации крови. Наиболее полно гемопоэз изучен у млекопитающих.
У млекопитающих в развитии крови как ткани в эмбриональный период выделяют 3 основных этапа, последовательно сменяющих друг друга:
мезобластический – развитие клеток крови во внезародышевых органах (мезенхиме стенки желточного мешка, хориона и стебля) протекает с 3 по 9 неделю развития зародыша человека, при этом появляется первая генерация ГСК;
печеночный – начинается с 5 – 6-й недели развития плода, основным органом гемопоэза становится печень, в ней образуется вторая генерация ГСК. Кроветворение достигает максимума через 5 месяцев и завершается перед рождением. ГСК заселяют тимус (с 7 – 8-й недели выявляются Т-лимфоциты), селезенку (с 12-й недели) и лимфатические узлы (с 10 – 12й недели);
медулярный (костномозговой) – появляется третья генерация ГСК в костном мозге, где гемопоэз начинается с 10-й недели и нарастает к рождению. После рождения костный мозг становится центральным органом гемопоэза.
Кроветворение в стенке желточного мешка. Первые этапы кроветворения происходят в желточном мешке, где найдены недифференцированные клетки – мезобласты, которые мигрируют в него из первичной полоски. Мезобласты обладают высокой митотической активностью и дифференцируются в клетки – первичные эритробласты. В течение нескольких часов после миграции происходят деление и дифференцировка мезобластов желточного мешка до первичных эритроцитов. Большинство этих клеток – ядросодержащие, но некоторые из них не содержат ядер. Другие мезобласты дифференцируются в клетки гемоцитобласты. Для эмбрионов кролика известна вторая стадия гемопоэза в желточном мешке, в ходе которой гемоцитобласты дифференцируются в эритробласты, которые становятся окончательными или вторичными нормобластами, которые теряют ядра и превращаются в нормоциты. В кровяных островках формируются сосудистые каналы, создаются первичные эритробласты и гемоцитобласты, а на более поздних этапах – зрелые эритробласты и эритроциты. Развитие эритроцитов в стенке желточного мешка происходит внутри первичных кровеносных сосудов – интраваскулярно. Одновременно экстраваскулярно из бластов, расположенных вокруг сосудистых стенок, дифференцируется небольшое количество гранулоцитов – нейтрофилов и эозинофилов. Часть ГСК остается в недифференцированном состоянии и разносится током крови по различным органам зародыша, где происходит их дальнейшая дифференцировка в клетки крови или соединительной ткани.
Кроветворение в печени. Печень у млекопитающих закладывается на 3 – 4-й неделе эмбриональной жизни и с 5-й недели становится центром кроветворения, которое происходит экстраваскулярно – по ходу капилляров, врастающих вместе с мезенхимой внутрь печеночных долек. Источником кроветворения служат ГСК, из которых образуются бласты, дифференцирующиеся во вторичные эритроциты. Одновременно с развитием эритроцитов в печени образуются зернистые лейкоциты – нейтрофилы и эозинофилы. Кроме гранулоцитов формируются гигантские клетки – мегакариоциты. К концу внутриутробного периода кроветворение в печени прекращается.
Кроветворение в тимусе. Тимус закладывается в конце первого месяца внутриутробного развития, и на 7 – 8-й неделе его эпителий начинает заселяться ГСК, которые дифференцируются в Т-лимфоциты.
Кроветворение в селезенке. Закладка селезенки происходит в конце первого месяца эмбриогенеза. Из ГСК экстраваскулярно образуются все клетки крови. Образование эритроцитов и гранулоцитов достигает максимума на пятом месяце эмбриогенеза.
Кроветворение в лимфатических узлах. Большинство лимфатических узлов закладывается на 9 – 10-й неделе; в них проникают ГСК, из которых на ранних стадиях дифференцируются эритроциты, гранулоциты и мегакариоциты. Формирование этих элементов быстро подавляется образованием лимфоцитов. Массовое «заселение» лимфатических узлов предшественниками Т- и В-лимфоцитов начинается с 16-й недели, когда образуются посткапиллярные венулы, через стенку которых осуществляется процесс миграции клеток. Из клеток-предшественников дифференцируются лимфобласты (большие лимфоциты), а затем средние и малые лимфоциты. Дифференцировка Т- и В-лимфоцитов происходит в Т- и В-зависимых зонах лимфатических узлов.
Кроветворение в костном мозге. Закладка костного мозга осуществляется на втором месяце эмбрионального развития. Первые ГСК появляются на 12-й неделе развития; в это время основную их массу составляют эритробласты и гранулоциты. Из ГСК в костном мозге формируются все клетки крови, развитие которых происходит экстраваскулярно. Часть ГСК остается в недифференцированном состоянии: они могут расселиться по другим органам и тканям и стать источником развития клеток крови и соединительной ткани. Таким образом, костный мозг становится центральным органом, осуществляющим универсальный гемопоэз, и остается им течение постнатальной жизни. Эмбриональный костный мозг отличается от центров более раннего развития гемопоэза активным образованием миелоидных клеток и доминированием этого процесса в гемопоэзе. Миелопоэз начинается в центральной части костномозговой полости и распространяется оттуда по всей полости кости. Эритропоэз в эмбриональном костном мозге развивается позже, чем в органах, рассмотренных раннее, и «смешивается» с процессом миелопоэза.