

Глава 3. Рецепторные нервные окончания
Главная функция афферентных нервных окончаний является восприятие сигналов поступающих из внешней и внутренней среды.
Рецептор - это терминальное ветвление дендрита чувствительной (рецепторной) нервной клетки.
Классификация рецепторов
В зависимости от локализации различают:
экстерорецепторы, воспринимающие раздражение внешней среды. Они расположены в наружных покровах тела, в коже и слизистых оболочках, в органах чувств;
интерорецепторы, получающие раздражение главным образом при изменении химического состава внутренней среды (хеморецепторы) и давления в тканях и органах (барорецепторы, механорецепторы);
проприорецепторы, воспринимающие раздражение в мышцах, сухожилиях, связках, фасциях, костях, суставных капсулах и т. д.
Проприорецепторы воспринимают тягу сокращения мышц, натяжения сухожилий и суставных капсул, возникающих при выполнении определенного движения или удержания частей тела в определенном положении. Это нервномышечные и нервно-сухожильные веретена, которые находятся в брюшке мышц или в их сухожилиях. Сухожильные органы (Гольджи) расположены внутри сухожилий, поблизости от мышц.
По специфичности восприятия (по модальности):
терморецепторы;
барорецепторы;
хеморецепторы;
механорецепторы;
болевые рецепторы.
В зависимости от специфичности раздражения, воспринимаемого данным видом рецептора, все чувствительные окончания делят на механорецепторы, барорецепторы, хеморецепторы, терморецепторы и некоторые другие.
Терморецепторы воспринимают изменение температуры, механорецепторы улавливают различные виды механических воздействий (прикосновение к коже, ее сдавление), ноцирецепторы воспринимают болевые раздражения. Барорецепторы представляют собой ветвящиеся свободные нервные окончания, залегающие в адвентициальном слое крупных артерий грудной полости и шеи. Наиболее важные из них - рецепторы, залегающие в стенках дуги аорты и сонного синуса. При растяжении стенки артерии под влиянием изменяющегося артериального давления эти окончания возбуждаются. Участие барорецепторов в регуляции артериального давления осуществляется по принципу обратной связи.
По строению (по морфологии) чувствительные окончания подразделяют на свободные нервные окончания, не имеющие какой-либо оболочки, и несвободные, содержащие в своем составе все компоненты нервного волокна, а именно ветвления осевого цилиндра и клетки глии.
свободные;
несвободные (инкапсулированные: пластинчатые тельца Фатера-Пачини, осязательные тельца Мейснера, концевые колбы Краузе, сухожильные органы Гольджи; неинкапсулированные).
Свободные (неинкапсулированные) нервные окончания имеются в коже. Подходя к эпидермису, нервное волокно теряет миелин, проникает через базальную мембрану в эпителиальный слой кожи. Нервные волокна разветвляются между эпителиоцитами вплоть до зернистого слоя, их веточки диаметром менее 0,2 мкм колбообразно расширяются на концах. Аналогичные концевые нервные окончания имеются в эпителии слизистой оболочки и роговицы глаза. Концевые свободные нервные окончания воспринимают боль, тепло и холод.
В эпидермисе свободные нервные окончания подходят также к измененным эпителиоцитам (клеткам Меркеля), которые имеют пальцевидные выросты. Эти окончания являются механорецепторами, воспринимающими давление.
Несвободные (инкапсулированные) нервные окончания - это осязательные тельца (Мейснера), пластинчатые (Фатера-Пачини), луковицеобразные (ГольджиМаццони). Все эти нервные окончания являются механорецепторами.
Пластинчатые тельца Фатера-Пачини - самые крупные из всех инкапсулированных нервных окончаний. Они имеют овальную форму, длину 3-4 мм и толщину 2 мм, располагаются в соединительной ткани внутренних органов и в подкожной основе Тельце снаружи покрыто соединительнотканной капсулой, имеющей пластинчатое строение. Под соединительнотканной оболочкой лежит наружная луковица, состоящая из 10-60 концентрических пластинок, образованных уплощенными периневральными эпителиоидными клетками. Войдя в тельце, нервное волокно теряет миелиновую оболочку, формирует внутреннюю луковицу и заканчивается колбообразным вздутием.
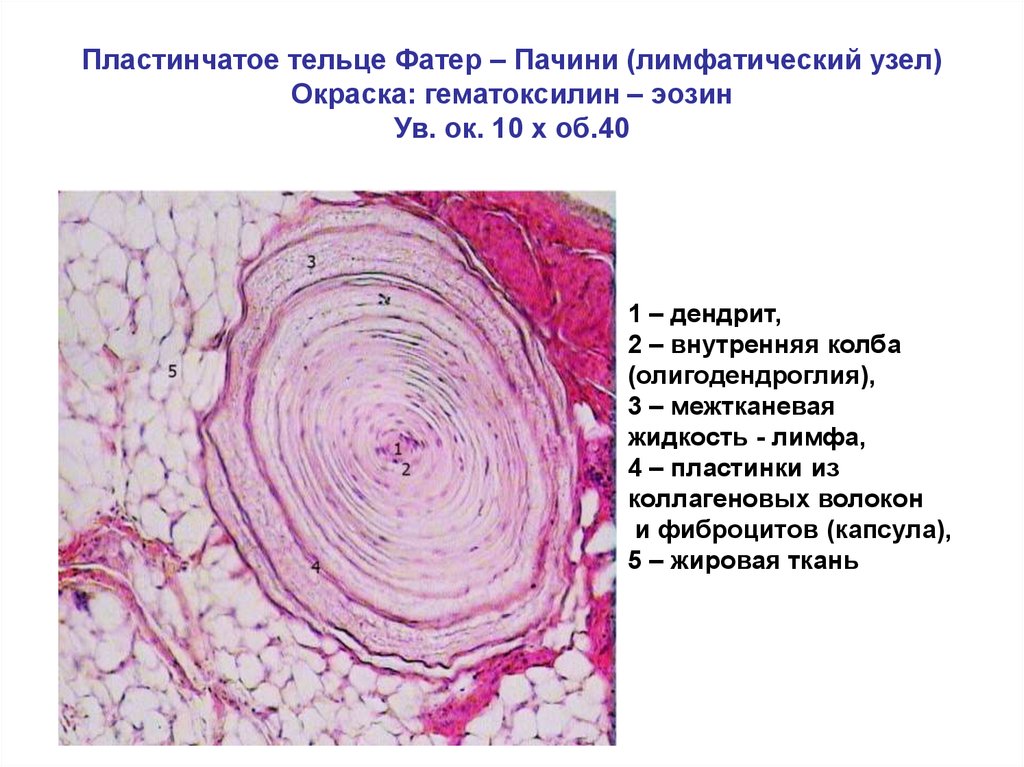
Рис. 3 – Пластинчатое тельце Фатера-Пачини
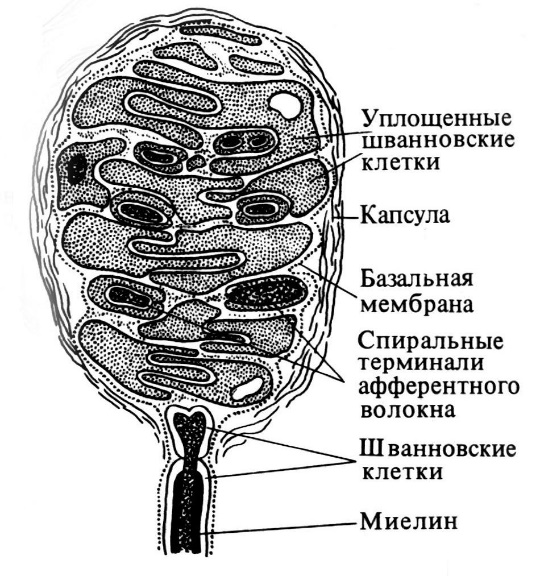 Осязательные
тельца (Мейсснера)
длиной 50-160 мкм, шириной около 60 мкм,
овальные или цилиндрические, особенно
многочисленны в сосочковом слое кожи
пальцев рук, а также в коже стопы. Тельце
образовано удлиненными, уплощенными
шванновскими клетками, лежащими одна
на другой. Нервное волокно, входя в
тельце, теряет миелин и оканчивается
колбообразным расширением. Периневрий
переходит в окружающую тельце капсулу,
образованную несколькими слоями
эпителиоидных периневральных клеток.
Сдавление этих клеток передается
осязательному тельцу, вызывая деформацию
его клеток, что приводит к возникновению
импульса в нервных волокнах. Таким
образом, мейсснеровские тельца являются
механорецепторами, воспринимающими
прикосновение, сдавление кожи.
Осязательные
тельца (Мейсснера)
длиной 50-160 мкм, шириной около 60 мкм,
овальные или цилиндрические, особенно
многочисленны в сосочковом слое кожи
пальцев рук, а также в коже стопы. Тельце
образовано удлиненными, уплощенными
шванновскими клетками, лежащими одна
на другой. Нервное волокно, входя в
тельце, теряет миелин и оканчивается
колбообразным расширением. Периневрий
переходит в окружающую тельце капсулу,
образованную несколькими слоями
эпителиоидных периневральных клеток.
Сдавление этих клеток передается
осязательному тельцу, вызывая деформацию
его клеток, что приводит к возникновению
импульса в нервных волокнах. Таким
образом, мейсснеровские тельца являются
механорецепторами, воспринимающими
прикосновение, сдавление кожи.
Рис. 4 – Осязательное тельце Мейсснера
Концевые колбы (Краузе) расположены в коже, конъюнктиве глаз, слизистой оболочке ротовой полости. Сферические тельца окружены толстой соединительнотканной капсулой, богатой коллагеновыми волокнами и фибробластами. Войдя в капсулу, нервное волокно теряет миелиновую оболочку и разветвляется в центре колбы, образуя множество веточек. Колбы Краузе воспринимают холод, возможно, они являются и механорецепторами.
Нервно-мышечное веретено крупное (длиной 3-5 мм и толщиной до 0,5 мм), окружено плотной соединительнотканной капсулой, состоящей из внутренней и наружной пластинок, в которую заключены от 2 до 12 мышечных, а также нервных волокон. Продольно расположенные веретена растягиваются вместе с мышцей и несут информацию о длине и степени растяжения мышцы.
Вставочный (ассоциативный), или кондукторный, нейрон осуществляет передачу нервного импульса с чувствительного (центростремительного) нейрона на двигательный или секреторный (центробежный).
Тела эфферентных (эффекторных, двигательных или секреторных) нейронов (от лат. Efferens - выносящий) находятся в центральной нервной системе или в симпатических и парасимпатических нервных узлах. Аксоны эфферентных нейронов несут нервные импульсы к рабочим органам (мышцам или железам), а также ко всем органам и тканям для иннервации (регуляции) обмена веществ. Выделяют также двигательные и секреторные окончания аксонов выносящих (эфферентных) нейронов. Аксоны двигательных нейронов разветвляются и каждый из них ин нервирует большое количество мышечных волокон.
Двигательные (моторные) нервные окончания располагаются на мышечных волокнах. Окончание одного двигательного нервного окончания и иннервируемое им поперечнополосатое мышечное волокно образуют двигательную единицу - мион.
Каждое мышечное волокно иннервируется веточкой аксона двигательного нейрона, который, оканчиваясь на волокне, образует нейромышечное окончание, или двигательную концевую пластинку, у которой различают пресинаптическую и постсинаптическую мембраны, разделенные синаптической щелью .
Нервный импульс, достигающий нервно-мышечного окончания, приводит к выходу в синаптическую щель ацетилхолина, который связывается со специфическими рецепторами мембраны мышечного волокна. Нервные окончания неисчерченной (гладкой) мышечной ткани располагаются между гладкомышечными клетками, где образуют расширения - синаптические пузырьки, плотно прилежащие к базальной мембране миоцитов.