

http
.docx
http://micro.moy.su/publ/obshhaja_mikrobiologija/fototrofnye_bakterii_i_fotosintez/fototrofnye_bakterii_i_fotosintez/15-1-0-148
Использование световой энергии галобактериями
Виды, относящиеся к роду Halobacterium (Н. halobium, H. cutirubrum), со ставляют высокоспециализированную в физиологическом отношении группу бактерий. Места их естественного обитания-высококонцентрированные или насыщенные растворы солей, такие как воды Большого соленого озера в США или места, где добывают соль путем выпарива ния морской воды. Приспособленность бактерий к существованию в столь экстремальных условиях связана с тем, что концентрация соли внутри клеток так же высока, как и в окружающей среде. Оптимальный рост таких бактерий наблюдается в 3,5-5,0 М растворе NaCl. Для нор мального функционирования выделенных из клеток ферментов тоже не обходимы более чем двухмолярные концентрации соли. Галобактерии относятся к архебактериям
|
Оксигенный фотосинтез Первичные процессы фотосинтеза протекают в тилакоидах - плоских замкнутых мембранных пузырьках, содержащихся в клетках цианобак терий и в хлоропластах водорослей и высших растений. Тилакоидные мембраны и светособирающие пигменты (пигменты антенн). Тилакоидная мембрана содержит в себе пигментные молекулы (хлорофилл а, хлорофилл b и каротиноиды), переносчики электронов и ферменты. Подавляющее большинство молекул хлорофилла ( > 99,5%), а также дополнительные пигменты (каротиноиды, фикобили-протеины) ответственны за поглощение света и распределение энергии; они образуют систему антенны. Лишь незначительная часть хлорофил ла авыполняет роль фотохимического реакционного центра, в котором протекает собственно фотохимическая окислительно-восстановительная реакция. Пигменты антенн (светособирающие пигменты) улавливают свет и передают энергию хлорофиллу реакционного центра (Каротиноид -> Каротиноид*; Хлорофилл + Каротиноид* -> Хлоро филл* + Каротиноид). Каротиноиды выполняют также защитную функцию: при очень ярком солнечном освещении они отдают избыточ ную энергию в окружающую среду и тем самым защищают молекулы хлорофилла от фотоокисления. Система светособирающих пигментов и реакционный центр объединены в так называемую фотосинтетиче скую единицу. Фототрофные бактерии и фотосинтез | Просмотров: 2393 | Добавил: Wiki | Дата: 30.12.2009 |
|
Аноксигенный фотосинтез Фотосинтетический транспорт электронов у анаэробных фототрофных бактерий во многих отношениях отличается от только что описанного. В аноксигенном фотосинтезе участвует только одна световая реакция; она поддерживает циклический транспорт электронов. Электроны, поки дающие цикл для восстановления NAD, не являются продуктом разло жения воды. Фотосинтез зависит от наличия в среде восстановленных субстратов и не сопровождается выделением 02. Собственно фотореак ция хотя и аналогична первой фотореакции у зеленых растений, однако у некоторых бактерий она приводит, вероятно, лишь к созданию про тонного потенциала и тем самым к запасанию энергии (АТР), но не к восстановлению NAD. Таким образом, нециклический перенос элек тронов (от донора электронов к пиридиннуклеотиду) здесь отсутствует. По-видимому, NADH2 образуется в результате какой-то темновой реак ции в ходе обратного транспорта электронов, протекающего с затратой энергии. Следует, однако, заметить, что у фототрофных бактерий между от дельными группами существуют гораздо большие различия в составе пигментов и механизмах фотосинтеза, чем у зеленых растений. В после дующем изложении мы вначале не будем касаться зеленых бактерий. |
http://micro.moy.su/publ/obshhaja_mikrobiologija/fototrofnye_bakterii_i_fotosintez/15
Возникновение фотосинтеза, использующего видимый свет.
Другой тип бактериального фотосинтеза обнаружен у пурпурных бактерий (рис, 5, б). Здесь действует набор ферментов, отличающихся от ферментного комплекса зеленых бактерий. Это несущая хлорофилл фотосистема 2 и комплекс III. Как и в предыдущем случае, процесс начинается с поглощения кванта хлорофиллом. Первоначально перенос электронов происходит по фотосистеме 2.
Затем вступает комплекс III, способный транспортировать электроны сопряженно с откачкой ионов Н+ из бактерии. Процесс завершается возвращением электрона с комплекса III на хлорофилл. Что касается ионов Н+, то они возвращаются в клетку через Н+-АТФ-синтазу, образуя АТФ.
Отличительная черта фотосинтеза у пурпурных бактерий состоит в том, что система не нуждается во внешнем доноре электронов. Откачка ионов Н* осуществляется путем циклического переноса электронов, поддерживаемого энергией света. Данное обстоятельство можно отнести, по-видимому, на счет эволюционного усовершенствования фотосинтеза пурпурными бактериями, которые по многим признакам являются эволюционно более продвинутой группой, чем зеленые серные бактерии.

Рис. 5. Хлорофиллычый фотосинтез зеленых серных (а) и пурпурных (б) бактерий: а - хлорофилл, связанный с особым белковым комплексом - фотосистемой 1 (ФС1), возбуждается квантом света и отдает электрон по цепи электронных переносчиков на НАД*. Восстанавливаясь, НАД* связывает внутриклеточный ион Н*. Потеря электрона на хлорофилле компенсируется окислением сероводорода до серы и иона Н* снаружи бактериальной клетки. Движение Н* внутрь клетки через комплекс F0 F, дает АТФ; б - хлорофилл, связанный с белком фотосистемы 2 (ФС2), поглощает квант света и запускает циклический перенос электронов. В этом процессе участвуют переносчики электронов ФС2 и дополнительного белкового комплекса III. Перенос электронов комплексом III сопряжен с откачкой ионов Н* из клетки. Откачанные ионы Н* возвращаются через комплекс F0 F, с образованием АТФ.
Следующим шагом в эволюции фотосинтеза стали, по-видимому, цианобактерии. Цепь переноса электронов в этом случае представляет собой комбинацию: а) фотосистемы 1 зеленых бактерий, б) фотосистемы 2 и комплекса III пурпурных бактерий и в) дополнительного комплекса, расщепляющего воду на О2 и Н+ (рис. 6). Фактически донором электронов вместо сероводорода (встречающегося в достаточных количествах лишь в некоторых природных нишах) служит вездесущая вода, запасы которой практически неограничены. В результате конечный акцептор электоров — НАДФ+ восстанавливается, а вода окисляется. Образующийся НАДФН окисляется затем сложной системой восстановления углекислого газа до глюкозы. Таким образом, фотосинтез цианобактерии параллельно с образованием АТФ дает углевод — одно из главных резервных веществ современных живых клеток. Нет сомнений, что цианобактерия является эволюционным предшественником хлоропластов — органелл зеленых растений, энергетика которых устроена в основном по той же схеме, что показана на рис. 6.
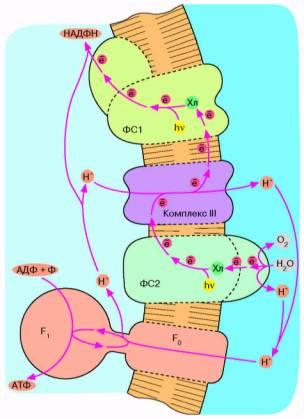
Рис. 6. Хлорофилльный фотосинтез цианобактерии. Квант света, поглощенный хлорофиллом фотосистемы 1, возбуждает перенос электронов по цепи, что завершается восстановлением НАДФ* до НАДФН. Окисленный хлорофилл фотосистемы 1 восстанавливается комплексом III, который, в свою очередь, получает электрон от фотосистемы 2. Допирование электрона фотосистемой 2 требует еще одного кванта света (поглощаемого хлорофиллом этой фотосистемы). Потеря электрона на хлорофилле фотосистемы 2 компенсируется за счет окисления молекулы воды до О2 и Н*. Ферменты, катализирующие всю цепь реакций переноса электрона от Н2О до НАДФ*, расположены в мембране таким образом, что ионы Н* откачиваются из бактериальной клетки, чтобы затем войти внутрь через F0F, и сделать АТФ. В хлоро-ппастах зеленых растений происходят те же события, но ориентация всех ферментов противоположна той, которая имеет место у цианобактерии и показана на рис. 6. Соответственно у хлоропластов фотосинтетическая цепь накачивает ионы ИГ внутрь, а комплекс F0F, переносит их наружу.
http://www.techstandard.ru/testars-423-3.html
Исследователем, который опроверг эту общепринятую теорию, был Корнелиус ван Ниль из Стамфордского университета, когда он, будучи еще студентом-дипломником, исследовал метаболизм различных фотосинтезирующих бактерий. Одна группа таких бактерий, а именно пурпурные серные бактерии, восстанавливает С до углеводов, но не выделяет О2. Пурпурным серным бактериям для фотосинтеза необходим сероводород. В результате фотосинтеза внутри бактериальных клеток накапливаются частицы серы. Ван Ниль обнаружил, что для этих бактерий уравнение фотосинтеза может быть записано как:
С О2 + 2Н2S свет = (CH2O) + Н2О + 2S
Этот факт не привлекал внимания исследователей до тех пор, пока ван Ниль не сделал смелого сообщения и не предложил следующего суммарного уравнения фотосинтеза:
С О2 + 2Н2А свет = (CH2O) + Н2О + 2А
В этом уравнении Н2А представляет собой либо воду, либо другое окисляемое вещество, например, сероводород или свободный Н2. У зеленых растений и водорослей Н2А = Н2О. То есть ван Ниль предположил, что Н2О, а не углекислота, разлагается при фотосинтезе. Эта блестящая идея, выдвинутая в тридцатые годы, экспериментально была доказана позднее, когда исследователи, использую тяжелый изотоп О2(18О2), проследили путь кислорода от воды до газообразного состояния:
С О2 + 2Н218О2 свет = (CH2O) + Н2О + 18О2
Таким образом, для водорослей или зеленых растений, у которых вода служит донором электронов, суммарное уравнение фотосинтеза записывается следующим образом:
6СО2 + 12Н2О свет = C6H12O6 + 6О2 + 6Н2О
http://xreferat.ru/66/417-1-fotosintez.html
|
Первичные процессы фотосинтеза |
|
|
|
Под фотосинтезом понимают происходящее в клетках фототрофных организмов преобразование световой энергии в биохимически доступную энергию (АТР) и восстановительную силу [NAD(P)H2], а также связанный с этим синтез клеточных компонентов. Фотосинтетическое фос-форилирование и фотосинтетическое восстановление пиридиннуклеоти-да-это процессы, ведущие к образованию первых стабильных продуктов фотосинтеза. К такому пониманию пришли в результате экспериментальных и теоретических исследований, основанных главным образом на сравнении фотосинтеза у фототрофных бактерий и у зеленых растений. После того как Виноградский (1888) установил, что для некоторых бактерий источником энергии при ассимиляции С02 отнюдь не всегда служит свет, а Энгельман (1883-1888) на основании своих физиологических исследований отнес пурпурные бактерии к фототрофным организмам, Будер (1919) показал, что эта группа бактерий обладает новым, не известным ранее типом метаболизма. Серные и несерные пурпурные бактерии ассимилируют С02 или органические вещества на свету. Однако такого рода фотосинтез существенно отличается от фотосинтеза высших растений: 1) вода здесь не может служить донором водорода, и поэтому такой фотосинтез не сопровождается выделением 02; 2) бактерии используют в качестве доноров водорода не воду, a H2S или органические вещества. Количественные исследования, проведенные на пурпурных серобактериях (ван Ниль, 1931). позволили вывести уравнение ассимиляции: С02 + 2H2S ^ <СН20> + Н20 + 2S 2С02 + H2S + 2Н20 Л 2<СН20> + H2S04 Если сравнить первое из этих уравнений с уравнением, описывающим фотосинтез у зеленых растений, можно заметить удивительную аналогию: У Chromatium С02 + 2H2S hZ <CH20> + Н20 + 2S У зеленых растений С02 + 2Н20 hZ <CH20> + Н20 + 20 Эта аналогия позволяет заключить, что в бактериальном фотосинтезе роль воды выполняет сероводород. Выведенное отсюда общее уравнение фотосинтеза hv С02 + 2Н2А - <СН20> + Н20 + 2A позволило выдвинуть гипотезу, согласно которой в основе всякого фотосинтеза лежит один и тот же первичный процесс, а разные типы фотосинтеза отличаются друг от друга только природой донора водорода (вода, сероводород или органические вещества). Этот общий первичный процесс видели сначала в фотолизе воды (Н20 + hv -» [Н] + [ОН])-в разложении ее на восстановительный и окислительный компоненты. Считали, что бактерии нуждаются в экзогенном доноре водорода Н2А, с помощью которого они восстанавливают окислительный компонент до воды, тогда как зеленые растения приобрели способность отщеплять от окислительного компонента кислород (4[ОН] -> 2Н20 + 02). Сейчас от этой концепции, рассматривающей разложение воды как первичный процесс фотосинтеза, пришлось отказаться, однако идея о переносе восстановительных эквивалентов с помощью энергии света сохраняется и в современной теории фотосинтеза. Первыми стабильными продуктами фотосинтеза являются АТР и восстановительная сила. Эти продукты можно обнаружить как в ин-тактных клетках и выделенных из них хлоропластах (у зеленых растений), так и в суспензиях фотосинтетических мембранных везикул из пурпурных бактерий. Фиксация С02 не обязательно сопряжена со световой реакцией. Она может происходить и как «темновая реакция», не зависящая от пигментсодержащих структур, при наличии АТР и NAD(P)H2. Эти два процесса разделены и в пространстве: фотосинтез происходит в мембранах и на их поверхности, а фиксация С02 - в цитоплазме или в строме хлоропластов. Как уже говорилось, оксигенный фотосинтез цианобактерий и растений отличается от аноксигенного фотосинтеза анаэробных фототрофных бактерий используемым донором водорода. Для того чтобы донором могла служить вода, необходимо последовательное осуществление двух фотореакций. Для использования же доноров с более отрицательным окислительно-восстановительным потенциалом достаточно одной-единственной фотореакции. Так как первичные процессы при оксигенном фотосинтезе изучены лучше, чем при аноксигенном, их мы и рассмотрим в первую очередь. |
http://microbiologu.ru/obschaya-mikrobiologiya/fototrofnyie-bakterii-i-fotosintez/pervichnyie-protsessyi-fotosi.html
В некоторых бактерий в процессе эволюции выработались другие биохимические пути синтеза углеводов. Бактериальный фотосинтез, в отличие от фотосинтеза высших растений, проходит в анаэробных условиях без выделения кислорода. Фотосинтез у бактерий осуществляется по типу фоторедукции, т.е. проходит восстановление диоксида углерода с поглощением лучистой энергии солнца, но без выделения кислорода.
Реакции бактериального фотосинтеза проходят с поглощением лучистой энергии бактериохлорофилл, а источником водорода для восстановления диоксида углерода является водород сульфид (сероводород), углеводы, органические кислоты и др.. Эти процессы играют важную роль в ассимиляции диоксида углерода мирового океана, а на глубинах Черного моря единственными живыми организмами является сиркобактерии, ведь для остальных живых организмов водород сульфит является сильным ядом.
http://estnauki.ru/ekologiya/7-ekologiya/11778-pervichnaja-produkcija-jekosistem-znachenie-i-osobennosti-processa-fotosinteza.html
. В этой связи чрезвычайно важным событием оказалось возникновение бактериального фотосинтеза, который привел к накоплению органических веществ уже биогенного происхождения. Однако бактериальный фотосинтез был бескислородным (аноксигенным) . И лишь значительно позднее цианобактерии (синезеленые) начали выделять при фотосинтезе кислород, который они получали в процессе фотолиза воды. Накопление в атмосфере Земли кислорода привело к появлению кислородно-озонового экрана, который защитил первичные живые организмы от губительного действия ультрафиолета и создал условия для существования аэробов.
игмент-белковые комплексы можно разделить на две функциональные группы. Основная функция первой из них - "светособирающих" или "антенных" пигмент-белковых комплексов - состоит в эффективном улавливании энергии света. Более 90% всего хлорофилла сосредоточено именно в "антенных" комплексах. Бóльшая их часть представляет собой белки с молекулярной массой от 20000 до 70000, на каждом из которых расположены 10 - 40 молекул хлорофилла. Помимо основного пигмента - хлорофилла а - в состав этих комплексов входит его аналог - хлорофилл б, а также каротиноиды - пигменты желтого или оранжевого цвета (один из них, бета-каротин, определяет окраску моркови). Цианобактерии содержат также бесхлорофилльные пигмент-белковые комплексы - фикобилины, окрашенные в синий или красный цвет. Они эффективно поглощают свет в области спектра, где хлорофилл имеет слабое поглощение, что позволяет осуществить более эффективное улавливание солнечной энергии.
Энергия возбуждения от светособирающих пигментов с очень высокой эффективностью (близкой к 100%) передается на хлорофилл другой группы пигмент-белковых комплексов - фотохимические реакционные центры. Их основная и важнейшая для всей биосферы функция состоит в том, чтобы преобразовать энергию электронного возбуждения хлорофилла (полученную в результате поглощения света в данном комплексе или в результате миграции возбуждения от светособирающих комплексов) в энергию разделенных зарядов, подобно тому, как это происходит в реакции Красновского, описанной выше.
У кислородвыделяющих организмов имеется два типа реакционных центров, функционирующих в так называемых фотосистеме 1 (ФС-1) и фотосистеме 2 (ФС-2) (рис. 5). Аналоги каждого из этих реакционных центров обнаружены у ряда фотосинте-зирующих бактерий, и молекулярная структура одного из них, выделенного из пурпурной бактерии Rhodopseudomonas viridis, впервые была установлена с помощью рентгеноструктурного анализа. Необходимо отметить, что значительная часть знаний о структурной и функциональной организации фотосинтетических реакционных центров получена при помощи изучения именно бактериальных реакционных центров. Они отличаются высокой стабильностью в изолированном из мембраны состоянии, и именно из них удалось впервые получить кристаллы, использование которых в рентгено-структурных исследованиях позволило получить данные о молекулярной структуре реакционного центра. За эту работу группа немецких исследователей в 1988 году получила Нобелевскую премию.
Бактериальный реакционный центр состоит из трех полипептидов с молекулярной массой от 24000 до 32000, на которых расположены молекулы одного из представителей хлорофиллов - бактериохло-рофилла (Бхл) и две молекулы его безмагниевого производного - бактериофеофитина (Бфф). Две из четырех молекул Бхл расположены настолько близко друг к другу (около 3 Å), что формируют компактное образование - димер (П), а четыре другие одиночные молекулы пигментов образуют две симметричные по отношению к П цепочки Бхл-Бфф. Энергия электронного возбуждения, полученная в результате поглощения света пигментами самого реакционного центра или в результате ее миграции от светособирающих комплексов, локализуется в конечном счете на П. Возбужденный П, обозначаемый как П*, за время около 10-12 сек передает свой электрон на одну из одиночных молекул пигмента, так что в результате образуется первичное состояние с разделенными зарядами П+Бфф-. Это состояние обладает очень коротким временем жизни (около 10-8 сек) вследствие возможного возврата электрона от Бфео- к П+. Однако вероятность этого процесса очень низка благодаря более быстрому (2 o 10-10 сек) "прямому" переносу от Бфео- к "стабильному" акцептору электрона, получившему название Q. При этом образуется состояние П+БфеоО , время жизни которого уже составляет около 10-3 сек. Необходимо отметить очень высокую эффективность описанных стадий фоторазделения зарядов. Так, квантовая эффективность, то есть доля поглощенных квантов, вызвавших перенос электрона, близка к 100%. Энергетическая эффективность, то есть доля энергии возбуждения, запасенной в результате фотохимического акта, составляет свыше 90% для первой стадии переноса электрона и около 50 - 70% для второй. Об изумительном совершенстве этой уникальной биологической "машины" свидетельствует, например, тот факт, что с такой же эффективностью фотоперенос электрона в реакционном центре осуществляется даже при температуре жидкого гелия (-271°С).
http://www.inventors.ru/fotosintez_i_biosfera.html