

f
Рис. 74. Convoluta sagittifera (Acoela). Многоклеточная сенсилла на продольном разрезе. (По: Иванов, 1952а).
сг — реснички, сер — клетки погруженного эпидермиса, cm — кольцевые мускульные волокна, / — пучок осязательных жгутов,
1Т — продольные мускульные волокна, п — нервный ствол, с которым связана сенсилла, par — паренхима, sc — скопление чувствительных клеток.
sc
(рис. 2, А; Ах, 1956; Riedl, 1969). Существуют они и у многих Acoela, например у Oxyposthia praedator, у которой отличаются большой компактностью, состоят из сотен чувствительных клеток и характеризуются определенной локализацией и более или менее определенным числом. Они расположены преимущественно в передней трети тела, где имеются 3 брюшные пары и 60— 65 дорсальных сенсилл (Иванов, 1952а). Столь же сильно развитые сенсиллы этого типа свойственны некоторым Convoluta. Так, у С. sagittifera имеется 30—40 сенсилл (рис. 74), лежащих на спине четырьмя продольными рядами. Среди Macrostomida такими же сенсиллами, расположенными метамерно, обладает Alaurina prolifera (Graff, 1904—1908). У Otoplana intermedia (Proseriata) крупные пучки осязательных жгутов локализованы на переднем конце тела (Hallez, 1910; Hofsten, 1921).
Развитие многочисленных, диффузно рассеянных по телу сенсилл несомненно есть более примитивное состояние; оно характерно для некоторых Acoela, например для Anaperus (Luther, 1912)
.
У Oxyposthia и некоторых Convoluta происходит их концентрация, сопровождающаяся уменьшением их числа и строгой локализацией. Таким образом, осязательные сенсиллы дают хороший пример множественной закладки органов и последующей их олигомеризации в понимании В. А. Догеля (1936, 1954).
Ресничные органы
Большой
интерес представляют ресничные органы
чувств. Как известно, у Xenoturbella
позади
брюшного рта поперек тела проходит
глубокая ресничная борозда (рис. 4);
кроме того, имеется пара продольных
латеральных ресничных бороздок.
Вероятно, эти образования несут
какую-то чувствительную функцию
(Westblad,
1949а).
Похожая чувствительная ресничная
бороздка, опоясывающая переднюю
часть тела впереди рта, развита у
некоторых Catenulida
(Catenula,
Suo-
mina
— рис.
75)
и
у некоторых Prolecithophora
Combinata.
Эти
образования невольно напоминают
прототрох трохофорных личинок.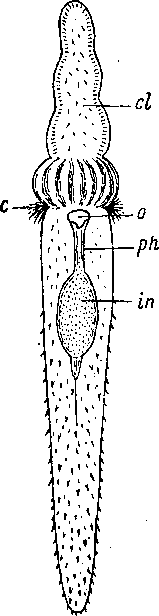
У многих турбеллярий имеются особые парные мерцательные ямки, в которых сосредоточены чувствительные клетки. В простейшем своем выражении они представлены маленькими участками чувствительного эпителия, лишенного рабдитных желез, лежащими на переднем конце тела у некоторых Typhloplanoi- da (Neorhabdocoela) и у части планарий. Очень хорошо они развиты у Catenulida; так, у Ste- nostomum это глубокие ямки (рис. 56), выстланные, помимо мерцательных, также железистыми клетками (рис. 76). Ямки вызывают небольшие водовороты, привлекающие к ним
Рис. 75. Suomina turgida (Catenulida). Общий вид с брюшной стороны. (По: Luther, I960).
с — ресничная бороздка, cl — головная лопасть со вздутым основанием, in — кишка, -ph — глотка, о — ротовое отверстие.
все новые порции воды, и выполняют функции органов химического чувства. Но наибольшего своего развития мерца- тельные ямки достигают у Prorhynchidae (Lecithoepitheliata), у которых они возникли независимо: Каждая ямка представляет собой ресничный канал, открывающийся в концевую камеру, образованную плоским эпителием, вероятно, несущим неподвижные чувствительные стереоцили, или микротрихи; в камеру открываются протоки специальных одноклеточных желез. У Хепорго
-
rhynchus обонятельные ямки особенно высоко дифференцированы (рис. 71); они способны выворачиваться и вворачиваться сокращением специальных мускулов протракторов и ретракторов. В вывернутом состоянии они функционируют как особенно чувствительные хеморецепторы (Reisinger, 1968).
К этой же категории мерцательных органов чувств относятся так называемые аурикулярные органы трикладид и другие органы «охотничьего обоняния», руководствуясь которыми турбеллярии
sep
дсо
Рис.
76. Stenostomum
sp. (Catenulida).
Разрез через чувствительную ресничную
ямку. (По: Kepner
a. Cash, 1915).
сг
— реснички на боковых стенках ямки,
fg
—
волокна ганглия, geo
—
ганглий ресничной ямки, т
— мускулы, s
—
секрет на дне ямки, sep
—
чувствительный эпителий, sp
—
чувствительные окончания сенсорных
клеток.
разыскивают свою добычу. У планарий аурикулярные органы, помимо обонятельной функции, помогают животному определять и движение воды (см.: Беклемишев, 1964).
Глаза
Глаза довольно широко распространены у турбеллярий и достигают у них довольно большого разнообразия. Они обычно парны, но Monocelis (Proseriata) имеет один непарный глаз. Число глаз варьирует от одной пары до нескольких сотен. Особенным обилием глаз отличаются Polycladida и Tricladida. У поликладид число глаз никогда не спускается ниже 12 (Aceros inconspicuus)\ у них имеются краевые и мозговые глаза. Первые лежат на края
х
тела, вторые расположены на спинной стороне над мозгом и близ него, а также возле мозговых щупалец. У планарий тоже различаются мозговые и краевые глаза. Водные трикладиды чаще всего обладают одной парой мозговых глаз (рис. 77, Б). Однако у некоторых водных планарий (Polycelis и др. — рис. 77, В), а также у крупных наземных развиваются множественные краевые
глаза.
Замечательно, что представители
самого примитивного семейства наземных
планарий — Rhynchodemi-
dae
—
обладают одной парой мозговых глаз,
которую они унаследовали от своих
водных предков, тогда как у остальных
наземных трикладид эти глаза редуцировались
и заменены независимо развившимися
многочисленными краевыми глазами
(рис. 77,^4). Так, Bipaliidae
и
Geoplanidae
имеют
множество мелких глаз, которыми усеяна
у некоторых видов вся спинная
поверхность.

Вопрос о происхождении множественных глаз у турбеллярий был поставлен и проанализирован В. А. Догелем, который по этому поводу писал: «Глаза являются одним из чрезвычайно пластичных органов во всем животном мире, многократно то появляясь, то исчезая в пределах одной и той же группы. При этом в пре-
ао тос
Рис. 77. Глаза и аурикулярные органы Tricladida.
А — наземная планария Geoplana техгсапа, Б — голова пресноводной планарии Dugesia polychroa, В — то же пресноводной планарии Polycelis tenuis (А — по: Hyman, 1939; Б ж В — по: Stei- mann, 1930). ао — аурикулярный орган, то с — краевые глаза, ос — глаз.
делах одной и той же группы могут в разное время возникать глаза, негомологичные друг другу, что затрудняет проведение правильного анализа». «У турбеллярий можно отличать глаза мозговые. . . от глаз краевых . . . Теоретически допустимо, что вторые могут возникать независимо от первых и даже после их исчезновения» (1954, стр. 34). Далее В. А. Догель приходит к выводу, что множественность глаз у Polycladida представляет собой первый этап развития этих органов, свидетельствующий о большой общей примитивности этого отряда. Поэтому ему кажется наиболее вероятным, что поликладиды с малочисленными глазами произошл
и
от многоглазых форм. У Tricladida, по В. А. Догелю, имел место аналогичный процесс множественной закладки краевых глаз, которые заменили собой исчезнувшую пару первичных мозговых глаз.
По строению и положению различают также эпителиальные и паренхимные глаза. Первые свойственны некоторым Acoela, например Otocelis rubropunctata, а также Macrostomida, например
Рис.
78. Строение глаз турбеллярий. (Из:
Беклемишев, 1964).
А
— Mesostoma
ehrenbergi (Neorhabdocoela),
В
— Platydemus
grandis (Tricladida Terricola).
cv
— стекловидное
тело, n
— ядро,
nf
— нервный
отросток зрительной клетки, рс
— пигмент, rh
—
светочувствительные палочки, sc
—
зрительная клетка.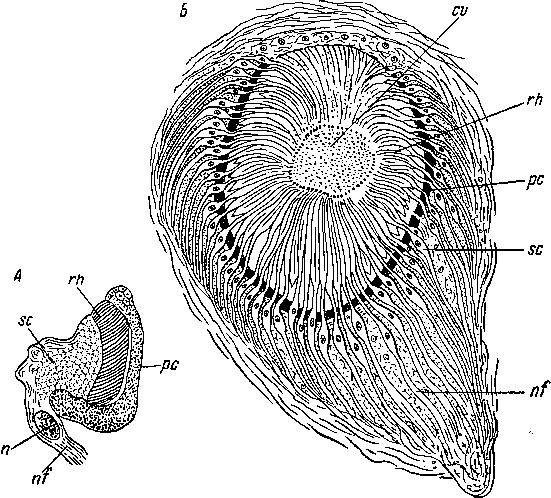
Microstomum и Alaurina. Структура этих глаз почти не изучена. По-видимому, они представляют собой участки эпидермиса с клетками, переполненными пигментом. С базальной стороны к ним прилегают светочувствительные клетки.
У остальных турбеллярий глаза лежат в толще паренхимы. Среди Neorhabdocoela известно немало таких форм, у которых каждый глаз состоит из одной световоспринимающей клетки, пигмент или отсутствует совсем, или представлен прилегающим к клетке диффузным скоплением пигмента паренхимы. Однако у большинства турбеллярий каждый глаз представлен пигментной чашечкой, или бокалом, образованным одной или несколькими клетками и содержащим от одного до нескольких десятков светочувствительных палочек, обращенных в глубь бокала и направленных от источника света (рис. 78, А). Эти глаза, следовательно, относятся к инвертированному типу. Они свойственны большинству Neoophora, исключая некоторых наземных планарий (см.: Беклемишев, 1937).
Но весьма высоко развиты глаза некоторых Polycladida. У них зрительные клетки образуют эпителиеобразную сетчатку, правда большей частью довольно несовершенную, так как в ней светочувствительные палочки еще не изолированы друг от друга пигментом. Наконец, у наземных планарий ретинальные глаза достигают наибольшего совершенства. Так, многие двуглазые Rhyn- chodemidae и Cotyloplanidae имеют мозговые глаза, состоящие из чаши пигментного эпителия, ее окружают узкие, расположенные радиально зрительные клетки (рис. 78, Б). Их центральные отростки уходят в глубь зрительного ганглия, в который погружен глаз, а периферические концы проникают в пространства между пигментными клетками. Здесь они образуют светочувствительные палочки, а продолжаясь внутрь глазной чаши, расширяются в прозрачные светопреломляющие призмы, образующие в своей совокупности стекловидное тело. На одной стороне стенка глаза прозрачна, это место представляет собой зрачок (Graff, 1889; Беклемишев, 1937). Беклемишев полагает, что строение такого глаза свидетельствует о способности животного видеть отдельные предметы.
Половой аппарат
Общие особенности полового аппарата
Половой аппарат ресничных червей представляет собой чрезвычайно пеструю картину и во многих случаях исключительно сложен. Общее представление об эволюции полового аппарата турбеллярий, по нашему мнению, наиболее верно дает В. Н. Беклемишев (1964).
При рассмотрении происхождения и филогении ресничных червей важно отметить, что все турбеллярии обладают внутренним оплодотворением и, за редкими исключениями, гермафро- дитны. Обе эти особенности обычно считаются вторичными и могут быть использованы как аргументы против утверждения о близости ресничных червей к первичным многоклеточным.
Однако внутреннее оплодотворение и гермафродитизм турбеллярий в простейших случаях весьма несовершенны и представлены самыми начальными этапами развития. Половой аппарат в таких случаях находится в состоянии, близком к тому, которое можно предполагать у самых примитивных Metazoa.
Если рассмотреть простейшие случаи внутреннего оплодотворения турбеллярий, то окажется, что они еще напоминают некоторые способы оплодотворения без копуляции. Многие бескишеч- ные ресничные черви обладают лишь мужским совокупительны
м
органом. При этом спаривание весьма несовершенно. Это либо
инъекция, либо так называемая импрегнация спермы. Инъекция
спермы может происходить в различных местах тела партнера, оНа осуществляется или копулятивным органом, снабженным кутикулярным стилетом, шипом и тому подобными образованиями, или мускулистым пенисом, который разрывает покровы партнера, функционируя как присоска. Последний способ нам удалось наблюдать у дальневосточного Plagiostomum karlingi Kulinitch, 1970 (рис. 79). Импрегнация, как полагают, состоит в приклеивании спермы к покровам партнера. Среди таких способов спаривания можно выделить крайне примитивный, когда сперма растекается по телу партнера и сперматозоиды самостоятельно проникают сквозь покровы. Тогда (как это имеет место, например, у Proantroporus murmanicus) многочисленные спермии партнера можно наблюдать в самых различных частях тела (Мамкаев, 1967). В данном случае половой процесс по сути дела недалеко ушел от таких примитивных форм осеменения, когда копуляция еще не происходит, но оплодотворение уже внутреннее: сперматозоиды самостоятельно добираются до яиц, находящихся в материнском организме (по М. С. Гилярову (1958), это наружно-внутреннее оплодотворение). Такое осеменение имеется у губок, у ряда кишечнополостных и у многих других форм. Например, у сцифо- медуз Суапеа и Aurelia яйца развиваются в карманах ротовых лопастей, но оплодотворяются еще в яичнике (Widersten, 1965). У Proantroporus murmanicus (отр. Acoela) спаривание уже происходит, но спермии все же самостоятельно проникают в организм партнера. Такое спаривание еще очень напоминает псевдокопуляцию, наблюдающуюся у многих форм с наружным оплодотворением. Таким образом, процесс копуляции развивается уже в пределах класса турбеллярий.
Что касается гермафродитизма, то это состояние также вряд ли может служить возражением против общей примитивности группы. Гермафродитизм свойствен большей части губок, в том числе всем Calcarea, которые считаются наиболее примитивными. Гермафро- дитны и гребневики, которых никто не рассматривает как вторичн
о
изменённые формы. Гермафродитны, наконец, и XenoturbeU lida.
Представление о гермафродитизме как о вторичном явлении вряд ли всегда справедливо. В большинстве случаев гермафродитизм покоится на очень низком состоянии развития сексуальности. Как правило, у гермафродитных беспозвоночных происходит смена половых фаз, нередко также имеются гермафродитные гонады или единая зачатковая зона, общая для мужских и женских половых элементов. Таким образом, один и тот же половой зачаток может давать то мужские, то женские гаметы или даже те и другие одновременно.
Все это дает основание полагать, что в таких случаях еще нет генотипического механизма определения мужских и женских половых клеток. В настоящее время общепринята гартмановская трактовка пола (Hartmann, 1956). В основе ее лежит представление о бисексуальной природе организмов. Исходя из этой теории, нетрудно представить себе, что развитие сексуальности с первых этапов может идти как по пути обособления полов, так и по пути гермафродитизма. По существу гермафродитизм — это наиболее полное выражение бисексуальной потенции организма. А поскольку бисексуальное состояние рассматривается как исходное в развитии полов, то и гермафродитизм можно считать весьма примитивным явлением.
Половые железы
В соответствии с изложенным за примитивное состояние половых желез турбеллярий естественно принять диффузную гермафродитную гонаду с нелокализованной зачатковой зоной; это представление получило широкое распространение (Steinbock, 1930—1931, 1966; An der Lan, 1936; Беклемишев, 1937, 1944, 1964; Karling, 1940; Westblad, 1949а; Hyman, 1951; Иванов, 1952а; Мамкаев, 1967). Однако Акс полагает, что диффузные гермафродитные гонады нельзя считать исходными для турбеллярий. Свое мнение он подкрепляет следующими соображениями (Ах, 1961, стр. 32).
Подавляющее большинство турбеллярий обладает обособленными, обычно парными семенниками и яичниками.
Гермафродитные гонады встречаются спорадически у отдельных представителей разных групп, для которых характерно разделение на семенники и яичники. При этом оба состояния могут встречаться даже в пределах одного рода (Haploposthia и Convoluta). Если эти изолированные, неправильно рассеянные случаи толковать как примитивные, то нельзя избежать тех методических трудностей, которые возникают и при выборе альтернативы, какая нервная система — эпителиальная или внутренняя — примитивнее.
Гермафродитная гонада очень легко может возникать вторично (например, у заднежаберных моллюсков; среди турбел-
лярий несомненная вторичная гермафродитная гонада известна у Itaspis среди Proseriata и у Puzostoma evelinae среди Prolecitho- phora).
Первый из этих аргументов, как и аналогичные соображения, рассмотренные ранее, показывает, что Акс склонен принимать за примитивное то состояние, которое широко распространено в пределах группы. Однако, как уже отмечалось, примитивные отношения далеко не всегда широко распространены. Напротив, у более или менее высокоразвитых форм они сохраняются редко, поскольку вытесняются более совершенным состоянием.
Третий аргумент Акса самостоятельного значения не имеет, так как вторичное возникновение какого-либо характерного состояния у одних видов, конечно, ни в какой мере не может служить препятствием для признания первичности подобного состояния у других, особенно у удаленных в систематическом отношении форм. Если даже у Itaspis и Puzostoma гермафродитные гонады действительно вторичны, то это еще не означает, что они вторичны всегда.
Весьма характерен второй аргумент Акса. По существу он связан с предыдущим: если за примитивное состояние принимается широко распространенное (и в том числе у представителей сравнительно низко организованных групп), то методически неправильно искать его среди изолированных, неправильно рассеянных случаев гермафродитных гонад. Нетрудно заметить, что здесь, как и при разборе других аппаратов, Акс опирается на один и тот же принцип (он сам подчеркивает, что в данном случае возникает та же ситуация, что и при рассмотрении нервной системы). Выше мы старались показать, что это слишком упрощенный подход к эволюционному процессу. И неравномерность развития различных органов, и возможность возврата тех или иных примитивных особенностей дают подчас пестрое сочетание прими- _ тивных и продвинутых признаков.
Отметим также, что гермафродитные гонады распространены далеко не так спорадически, как представляет себе Акс. Прежде всего гермафродитные гонады встречаются среди представителей сем. Haploposthiidae, которых Вестблад (Westblad, 1948), на наш взгляд, вполне убедительно трактует как самых примитивных бескишечных турбеллярий. Здесь наблюдаются гонады весьма разнообразного строения, которые демонстрируют прекрасную картину развития семенников и яичников. У Haploposthia rubra и Н. Ъгипеа имеется очень просто устроенная диффузная половая железа с нелокализованной зачатковой зоной и с перемешанными мужскими и женскими элементами, смещенными в дорсальном направлении (Westblad, 1945). У Н. rubropunctata при таком же расположении диффузной гонады наблюдается раздельнополое состояние (Westblad, 1945). Однако у женских экземпляров в яичнике, особенно в его передней области, встречаются и спермато- цитд. Последнее обстоятельство мы склонны трактовать как сви-
детельство предшествующего гермафродитного состояния, т. е. данный случай (как и аналогичные, приводимые далее) можно рассматривать как иллюстрацию перехода от исходной бисексуальности к раздельнополости. Эта раздельнополость могла развиться на основе обычного для турбеллярий обособления мужской и женской половых фаз во времени. Не исключено, однако, что у Н. rubropunctata мы имеем еще не настоящее раздельнополое состояние, а лишь резкую смену половых фаз. Н. microfoca имеет парные диффузные гермафродитные гонады (Marcus, 1950), ауЯ. viridis парные гонады начинаются впереди смешанной зачатковой зоной, несколько отступя назад они разделяются на дорсальные семенники и вентральные яичники (Westblad, 1945). Данный случай подводит нас к тому уровню развития половых желез, который наиболее распространен у Acoela и других турбеллярий. Однако гаплопостии демонстрируют нам и другие состояния: непарный дорсальный семенник и непарный вентральный яичник (Н. monogonophora, см.: Westblad, 1946), непарный дорсальный или парный дорсо-латеральный семенник и непарный вентральный яичник (Н. albiventer, см.: Marcus, 1954), у Н. opis- thorchis яичник такой же, но парный семенник смещен к заднему концу тела (Мамкаев, 1967).
Мы считаем, что приведенные случаи естественнее всего толковать как эволюцию, берущую начало от диффузной гермафродитной гонады (рис. 80). Здесь мы видим четкий морфологический ряд, наглядно демонстрирующий нам эволюцию от чрезвычайно простого состояния к более совершенному. С этих позиций нетрудно представить и формирование непарной яйцевидной гермафродитной гонады Diopisthoporus longitubus, которая разделяется в каудальном направлении на дорсальный семенник и вентральный яичник (Westblad, 1940). Представители данного вида обнаруживают и разделение полов, которое мы трактуем так же, как раздельнополость Н. rubropunctata.1 Диопистопорид проще всего связать непосредственно с гаплопостиидами, а их гонаду выводить непосредственно от диффузной гермафродитной гонады последних.
Парные
гермафродитные половые железы, кроме
Н.
micro-
phoca,
отмечены
еще у нескольких бескишечных турбеллярий:
Paratomella
unichaeta,
Convoluta
(Pseudohaplogonaria) viridipun- ctata, Mecynostomum pellitum
и
Polysolenoposthia
porsildi
(An der Lan, 1936; Westblad, 1946; Marcus, 1951; Dorjes, 1966,
1968). На
первый взгляд это те отдельные,
изолированные случаи, о каких пишет
Акс. Однако следует отметить, что у
бескишечных турбеллярий наблюдаются
семенники и яичники с общими зачат-£
.тфЦг«|#®
•■'<&. 1 ,-йг
.
‘Я
#
'й;:
Щ
,-х(©=1 @
/.оЛ
(Ге;ч>
'о-;'
ч>-/'

am
([ «Л
V ••• /

am
Д
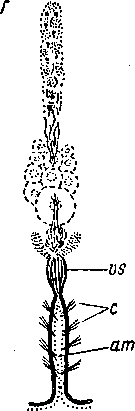
щ
{с- -
am
Рис. 80. Типы гонад и копулятивных органов у представителей родов Haplo- posthia (А, Б, Г—Е) и Paranaperus (В) (Acoela). (По: Westblad, 1948).
А — Haploposthia rubropunctata, женское состояние (яичник диффузный, в передней области также некоторое количество сперматид); Б — Н. rubropunctata, мужское состояние; В — Paranaperus pellucidus, семенники и яичники обособленные, парные (нарисованы только семенники); Г — Haploposthia monogonophora, семенники и яичники обособленные, непарные; Д — Н. rubra, диффузная гермафродитная гонада; Е — Я. viridis, парные гонады со смешанной зачатковой зоной, разделяющиеся в каудальном направлении на дорсальные семенники и вентральные яичники, am — трубковидный антрум, с — кути- кулярные палочки, р — пенис, составленный из кушкулярных палочек, vs — семенной
пузырь
.
новыми зонами, причем в ряде случаев имеет место переходное состояние, аналогичное таковому Haploposthia viridis или Diopisthoporus longitubus. Можно назвать ряд видов с общей зачатковой зоной гонад: Haploposthia opisthorchis, Haplogonaria sinubur- salia (как и у Diopisthoporus longitubus, у этого представителя гаплопостиид наряду с гермафродитным состоянием хорошо выражены мужское и женское), Nemertoderma bathycola, Actino- posthia beklemischevi (у нее также известны чисто мужское и чисто женское состояния), Proantroporus murmanicus, Antigonaria are- naria, Hofsteniola pardii, Philaetinoposthia diploposthia, Pseudacti- noposthia saltans, Pseudomecynostomum fragilis, Solenofilomorpha longissima, Oligochoerus erythrophtalmus, 0. bakuensis, 0. chlorella, Convoluta convoluta, C. bifoveolata, Archaphanostoma histobursalium (у последней хорошо выражены чисто мужское и чисто женское состояния), Bursosaphia baltalimaniaformis, у которой хорошо выраженная мужская фаза сменяется гермафродитной 1(список составлен по работам:Papi, 1957; Беклемишев, 1963а; Мамкаев, 1965, 1967, 1971; Dorjes, 1968). На рассматриваемую сторону организации турбеллярий, к сожалению, редко обращается внимание, но, насколько можно судить по рисункам Вестблада, Маркуса, Дорьеса и некоторых других авторов, у Acoela общие зачатковые зоны широко распространены. Так, по рисункам Дорьеса (Dorjes, 1968), зачатковые зоны, по-видимому, еще не полностью обособились у таких видов, как Haplogonaria syltensis, Pseudohaplogonaria vacua, Otocelis rubropunctata, Archocelis macro- rhabditis, Philocelis cellata, Anaperus tvaerminnensis, Achoerus pa- chycaudatus, Philachoerus johanni, Paraphanostoma trianguliferum, Pelophila cavernosa, P. pachymorpha, Philaetinoposthia saliens, Pseudomecynostomum granulum, Archaphanostoma macrospiriferum, Praeconvoluta karinae. Мы полагаем, что общая зачатковая зона будет обнаружена у многих Acoela, а возможно, и у ряда представителей Prolecithophora. Уже сейчас данные о гермафродитных гонадах и 'общих зачатковых зонах так многочисленны, что соображения Акса о спорадическом распространении гермафродитных гонад не соответствуют действительности. Следует подчеркнуть, что общие зачатковые зоны трудно трактовать как переход к гермафродитным гонадам. В этом случае пришлось бы говорить о широко распространенной тенденции к образованию гермафродитных желез, которая реализовалась, однако, почему-то лишь у отдельных видов. Это означало бы, кроме того, что у многих видов эволюция шла от более совершенных половых
Жблёз
к несовершенным. Но для таких фантастических
построений в настоящее время нет
уже никаких оснований, даже если принять
во внимание замечания Акса. Общие
зачатковые зоны распространены
достаточно широко, а тенденции в эволюции
гонад у турбеллярий вполне определенны.
Это обособление половых желез от
паренхимы, дифференциация семенников
и яичников, локализация у них
зачатковых зон, развитие компактных
гонад, снабженных оболочкой (tunica
propria). Из
диффузных гонады обычно становятся
парными, а затем в ряде случаев и
непарными (особенно у представителей
псаммона). Во многих случаях формы,
обладающие гермафродитной гонадой и
семенниками и яичниками с общей
зачатковой зоной, можно связать
непосредственно с кругом самых
примитивных бескишечных турбеллярий
(прежде всего это представители семейства
Haplo-
posthiidae).
От
таких Acoela,
кроме
Diopisthoporus,
выводятся
и Nemertoderma,
и
Paratomella,
и
Solenofilomorpha,
Antigonaria, Hofsteniola,
относящиеся,
по-видимому, к близким группам, а также
Actinoposthia
и
другие Childiidae
и
ряд других форм. Представителей семейства
Convolutidae
(а
также Otocelididae)
можно
выводить от гаплопостиид через анаперид.
В отдельных случаях рассматриваемое
состояние может сохраняться (или
выявляться вновь), напоминая нам об
исходном состоянии гонады. За пределами
Acoela
гермафродитные
гонады известны у Tyrrhe-
niella sigillata
(Riedl, 1959a) — единственного
морского представителя отряда
Catenulida,
у
которого довольно компактная гонада,
непарная и вентральная, и у представителя
отряда Prolecithophora,
именно
у Prolecithoplana
lutheri
с
ее диффузной гонадой со слабой
тенденцией к парности (рис. 106; Karling,
1940).
У Puzostoma
evelinae
—
две компактные гермафродитные гонады
и два фолликулярных вителлария (Marcus,
1950).
По всей вероятности, не совсем еще
разделились мужская и женская зачатковые
зоны у Plicastoma
norvegicum.
Такое
состояние можно ожидать и у некоторых
других плагиостомид. По-видимому, к
данному отряду можно отнести и Ichtyophaga
subcutanea
(Сыромятникова,
1949; Меницкий, 1963), обладающую диффузной,
парной дорсо-лате- ральной гонадой
(желточники у нее не обнаружены, но это
можно объяснить неполной половой
зрелостью). Все эти формы относятся к
двум весьма примитивным отрядам,
исходное состояние которых может
выводиться из организации Acoela.
Что
касается упоминаемой Аксом Itaspis
evelinae
(Proseriata,
Otoplanidae),
то гермафродитная гонада отмечена
здесь лишь у одного экземпляра, у
которого оба яичника на одной стороне
развились как гермафродитные железы
— овотестисы (Marcus,
1952).
Анормальное развитие гонад, и в частности
образование овотестисов, встречается
у отдельных особей и среди высших
животных, даже среди позвоночных.
Изменения в формировании половых
органов могут быть вызваны целым рядом
причин. Вряд ли такие случаи можно
использовать для решения
сравнительно-анатомических вопросов.
Напоминанием о примитивном Состоянии половых желез служат также гонады с диффузной нелокализованной зачатковой зоной. Обычно зачатковая зона располагается спереди, однако у Paratomella unichaeta она растянута по всей длине гермафродитных гонад (Dorjes, 1966), а у Oxyposthia praedator (рис. 81) — по всей длине диффузных семенников и яичников (Иванов, 1952а); очевидно, обособление семенников и яичников здесь произошло раньше, чем локализация зачатковой зоны. В семенниках нелока- лизованная зачатковая зона отмечена также у Otocelis sachalinen- sis, Convoluta sagittifera (Иванов, 1952a), Convoluta bifoveolata (Мамкаев, 1971), Proantroporus murmanicus (Мамкаев, 1967), Ackoerus pachycaudatus, Philachoerus johanni, Pelophila cavernosa, Pseudaphanostoma brevicau- ^ n 0Q
datum, P. pelophilum, Bur- , I
sosaphia
baltalimaniformis,
'
Praeaphanostoma chaetocau- datum (Dorjes, 1968) и
Рис. 82. Фолликул гермовител- лярия Prorhynchus. (По: Hyman, 1951).
с/ — желточные клетки фолликула, оо — яйцо, п — ядро яйца.
на некоторых стадиях у Actinoposthia beklemischevi (Мамкаев, 1965). Хотя это состояние найдено пока у небольшого числа видов, мы не можем рассматривать его как вторичное. Если сравнить последний список с двумя предыдущими, то можно заметить, что у некоторых форм имеются оба примитивных признака. Вряд ли это совпадение случайно. Семенники у Acoela обычно лежат дорсальнее яичников, но встречается и обратное расположение (Achoerus pachicaudatus, Pseudomecynostomum fragilis, So- lenofilomorpha longissima). Гонада в целом может занимать дорсальное (Nemertoderma) или вентральное (Convoluta convoluta, С. viridipunctata) положение, хотя обычно яичник вентральнее или вентро-латеральнее центральной паренхимы, а семенники дорсальнее или дорсо-латеральнее. Сказанное свидетельствует, что у Acoela еще не закончено формирование семенников и яичников; бескишеч- ные турбеллярии дают богатый материал для исследования эволюции половых желез начиная с самых примитивных состояний.
У высших турбеллярий (Neoophora) женские половые железы дифференцируются на собственно яичники (гермарии) и желточ- ники (вителлярии). Иногда морфологически вителлярии еще не полностью обособлены от гермариев, и тогда говорят о гермовител- ляриях. Как известно, желточные клетки — это видоизмененные ооциты, утратившие способность к оплодотворению и развитию, специализированные для образования желтка. Эти клетки не используются для питания растущей яйцеклетки, а входят в состав так называемого сложного яйца и служат для питания раз-
А. В. Иванов и Ю. В. Мамкаев
129
вивающегося зародыша. Таким образом, желточные клетки Neo- ophora резко отличаются от распространенных у других животных желточных клеток, питающих растущий ооцит (Беклемишев, 1964).
Можно
выявить два пути формирования
желточников. Один из них состоит в том,
что вокруг ооцита образуется фолликул
из желточных клеток (рис. 82). Сходные
фолликулы известны еще у Acoela
(.Hofstenia,
Oligochoerus,
Polychoerus, Am- phiscolops).
В
таких случаях женские гонады даже
называют иногда гермовител- ляриями
(Westblad,
1948;
Hyman,
1951).
Однако здесь фолликулярные клетки
служат еще для питания растущего
ооцита, яйцеклетка содержит желток и
сложного яйца не образуется. У
представителей отряда Lecithoepitheliata
такие
фолликулы (рис. 83) становятся настоящими
гермо- вителляриями, поскольку у них за
счет желточных клеток образуется сложное
яйцо. При этом у Gnosonesimidae
мы
находим четыре гермовителлярия,
состоящих из фолликулов, а у Prorhynchidae
уже
только один (см. рис. 105, Б
и Е).
Другой путь формирования желточников можно наблюдать у представителей Prolecithophora. У них, как мы видим, в примитивных случаях имеется еще диф-
Рис. 83. Gnosonesima borealis (Lecithoepitheliata). Общий вид. (Из: Westblad, 1954).
g — мозг, gvi — гермовителлярии, г — кишка, о — рот, pi — женское половое отверстие, ph — глотка, рт — мужское половое отверстие, t — семенник, v — влагалище, vg — пузырек зернистого секрета, vs — семенной пузырек.
фузная гермафродитная гонада, в которой перемешаны мужские элементы и женские, уже дифференцированные на способные к развитию яйца и служащие для питания зародыша желточные клетки (рис. 106). Далее в пределах данной группы происходят отделение семенников от гермовителляриев и разделение последних на гермарии и вителлярии; фолликулов вокруг ооцитов при этом не образуется (Karling, 1940).
Половые протоки
Рассмотрим далее вопрос о возникновении половых протоков. Акс считает, что предок турбеллярий уже обладал как семяпроводами, так и яйцеводами. Однако в простейших случаях у тур- беллярий ни тех, ни других не имеется. Свою точку зрения относительно семяпроводов Акс по существу не аргументирует (Ах, 1961, стр. 33). Что касается женских выводных протоков, то он считает, что у Acoela, которые яйцеводов не имеют, яйца первоначально выходили через вагину и лишь позднее этот способ заменился выведением через рот или стенку тела (очевидно, при этом следует принять вторичную утрату бурсы у Abursalia). Этот способ выведения яиц расценивается Аксом как напоминание о первичном наличии у турбеллярий женских выводных путей. В подтверждение своей точки зрения Акс приводит мнения Мейкснера (Meixner, 1926) и Маркуса (Marcus, 1946). Они считали, что клейкие железы, секрет которых образует студенистую кладку, открываются у многих Acoela в вагину. Сйедует отметить, однако, что у бескишечных турбеллярий хорошо развиты кожные железы, которые, скорее всего, и продуцируют студенистую субстанцию, окружающую пакет яиц. Представить выведение относительно крупных яиц через бурсу, снабженную обычно узким ductus spermaticus, часто кутикуляризированным, очень трудно; кроме того, как будет показано ниже, сами бурсы несомненно возникают в пределах Acoela. Свою точку зрения Акс подкрепляет еще двумя аргументами.
Acoela и в других своих существенных особенностях (синцитиальность, пищеварительный аппарат, способ дробления) вторично изменены по сравнению с Polycladida жМасго- stomida.
СредиNeoophora, первично обладающих женскими половыми путями, имеются прекрасные модели, демонстрирующие, как яйцо или эмбрион вторично выводятся через кишечник (Baicalellia evelinae и Bresslauilla relicta из Neorhabdocoela Dalyellioida, Ethomorhynchus anophthalmus и представители Rogneda из Neorhabdocoela Kalyptorhynchia). Принятие первичного отсутствия яйцеводов должно привести к выводу об их многократном возникновении в пределах Neoophora, что неприемлемо из методических соображений.
Что касается первого из этих положений, то оно уже разобрано в соответствующих разделах. Второй аргумент демонстрирует нам тот же логически неверный ход рассуждений, который отмечен ранее. Из примеров вторичного отсутствия яйцеводов вовсе не следует невозможность их первичного отсутствия.
У Acoela известны случаи усложнения гонад, и в частности формирование выводных путей. Так, у Amphiscolops sargassi (Marcus, 1950), A. bermudensis (Hyman, 1939) и A. gemelliphorus (Marcus, 1954) имеются два своеобразных яйцевода, которые открываются наружу парой отверстий, расположенных по бокам от бурсы. Своеобразные семяпроводы с мускулистыми стенками имеют Oligochoerus bakuensis и О. chlorella, причем у первого вида мышечные волокна образуют неправильное сплетение, а у второго — два слоя: внутренний кольцевой и наружный продольный
(Беклемишев, 1963а). Сходные простейшие семяпроводы описаны' также у Convoluta convoluta (см.: Мамкаев, 1967). Семяпроводы отмечены и у Otocelis chiridotae (Беклемишев, 1915). Все эти несовершенные, своеобразные половые пути нельзя трактовать как рудименты. Они настолько отличаются от выводных путей других турбеллярий, что их можно рассматривать только как новообразования. Таким образом, среди Acoela мы видим независимое формирование половых путей. Так же самостоятельно, параллельно они могут возникать и в других группах. Следует отметить, что вторичность отсутствия женских выводных путей у Bresslauilla далеко не так очевидна, как представляет Акс. Первичное их отсутствие в данном случае принимает не только В. Н. Беклемишев, о котором упоминает Акс, но и Рейзингер (Reisinger, 1929), и Штейнбек (Steinbock, 1924, 1966).
Женские копулятивные органы
Рассмотрим далее вопрос о происхождении женских копуля- тивных органов и о природе их связи с кишечником, которая иногда имеется в виде так называемого ductus genito-intestinalis. Этот вопрос по-разному решается различными авторами. Акс, например, считает, что прототипу турбеллярий уже были свойственны не только выводные протоки, но и женские копулятивные органы в виде семенной сумки — бурсы с влагалищем (вагиной) и женским половым отверстием, открывающимся перед мужским копулятивным органом. Это утверждение по существу не аргументируется. Отмечается лишь наличие такой бурсы у макросто- мид, части Acoela, Polycladida и предполагается гомология ее с первичным бурсальным органом в пределах Neoophora (Proseriata, Tricladida).
Разнообразие женских копулятивных органов у турбеллярий необычайно велико, и для решения вопроса о их происхождении необходимо разобрать хотя бы основные их типы. По функции различаются три типа женских половых органов, связанных с копуляцией и оплодотворением: bursa copulatrix, bursa semina- lis и receptaculum seminis. Копулятивная бурса (или только канал-влагалище) служит для приема совокупительного органа партнера. В семенной бурсе хранится сперма партнера. Иногда даже при наличии копулятивной и семенной бурс развивается еще семеприемник — особый мешочек, где сперма накапливается непосредственно близ места оплодотворения яиц. 1Эти три типа органов соответствуют разным сторонам процессов совокупления и осеменения, но часто бурсальный орган выполняет несколько функций. Так,у Acoela распространена бурса с влагалищем и жен-
г,ким копулятивным отверстием, которая служит как для копуляции, так и для хранения спермы; при этом возникает приспособление и для подведения спермы к оплодотворяемому яйцу — ductus spermaticus, иногда представляющий собой кутикулярный наконечник.
По способу возникновения можно различать паренхимные и эпителиальные бурсы (Karling, 1940, 1963). Паренхимные бурсы возникают безусловно как органы для хранения спермы партнера. В примитивных случаях это просто особый участок паренхимы (рис. 84, А, Б), в котором содержится сперма партнера (например, Haplogonaria macrobursalia, Diopisthoporus psammophilus, Archaphanostoma agile, A. histobursalium, A. macrospiriferum из Acoela; Pseudostoma histobursalium из Prolecithophora; Uncinorhynchus westbladi, U. flavidus из Neorhabdocoela Kalyptorhynchia). Этот участок паренхимы затем превращается в мешковидное образование с собственными стенками (рис. 84, В) (например, у Praeaphanostoma, Haplogonaria macrobursalia, Н. glandulifera из Acoela; у Cheliplana stylifera из Neorhabdocoela Kalyptorhynchia). Далее в органах такого типа развиваются ductus spermaticus и канал, по которому сперма попадает в бурсу (рис. 26, 46, 85). Последний может представлять собой влагалище, и тогда семенная бурса становится копулятивной. Такие бурсы широко распространены у Acoela, но известны и у других турбеллярий (например, у Cheliplana hipergina из Kalyptorhynchia). В качестве наконечников бурс у Acoela, по-видимому, могут использоваться кутикулярные наконечники грушевидных органов, развивающихся под эпидермисом; возможно, в области «бурсальной паренхимы» они превращаются в ductus spermaticus. Паренхимные семенные бурсы очень часто, но далеко не всегда становятся копулятивными; в ряде случаев канал, по которому сперма попадает в bursa seminalis, не имеет функций влагалища (например, у Polycystis dolichoce- phala, P. nageli, Parautelga bilioi из Kalyptorhynchia).
Эпителиальные бурсы (рис. 86) могут возникать как впячива- ние стенки тела (например, у Bursosaphia baltalimaniformis из Acoela, у Cheliplana stylifera из Kalyptorhynchia) или как впячи- вание эпителиев половых путей (например, у Pseudograffilla areni- cola, Halamovortex nigrifrons из Neorhabdocoela Dalyellioida; у An- nulorhynchus adriaticus из Neorhabdocoela Kalyptorhynchia). Иногда эти бурсы- функционируют только как семенные, в других случаях бурсы, образующиеся из эпителиев, служат сразу и как семенные, и как копулятивные или только как копулятивные.
У
некоторых турбеллярий имеется канал,
соединяющий женскую половую систему
с кишечником (ductus
genito-intestinalis).
Для понимания природы этого образования
следует напомнить, что половой аппарат
турбеллярий выполняет не только функции
введения спермы в организм партнера,
хранения ее и подведения к оплодотворяемым
яйцам; иногда возникает также потребность
в удалении излишка спермы. Она может
удовлетворяться либо
ooc
sp hb vs

/ -f-——\ j pm да
ooc sp hb vs
В
ooc sp
Рис.
84.
Копулятивный
аппарат Archaphanostoma
macrospiriferum (A),
A■
Mstobursalium
(Б)
и Haplogonaria
macrobursalia (B) (Acoela).
(По:
Dories.
1968).
am — мужской антрум, Ьа — семенная бурса, ga — акцессорные железы, ЛЬ — бурсальная ткань, mdv — дорсо-вентральные миофибриллы, оос — ооциты, р — циррус, pg
половое отверстие, рт — мужское половое отверстие, sg — зернистый секрет, sp — клубки спермиев партнера, vs — семенной пузырек.
Путём резорбции спермы в бурсальных органах (в связй с Этим иногда обособляется особый отдел бурсы), либо путем удаления ее в кишку через специальный проток или пору.
Относительно
происхождения бурс и генитально-интестинальных
связей имеется несколько гипотез.
Ремане (Remane,
1951,
1958) считает, что семенная бурса турбеллярий
представляет собой участок задней
кишки, наличие которой он предполагает
у предков турбеллярий. При этом
женское копулятивное отверстие го-
мологизируется анусу исходных форм, а
генито-интестинальный проток
рассматривается как первичное состояние,
наследие пред-
Рис. 85. Praeaphanostoma longum (Acoela). Копулятивный . ^ аппарат. (По: Dorjes, 1968).
bs —гсеменная бурса, ov — яичник, р — пенис, р/ — женское половое отверстие, рт — мужское половое отверстие, sp — пучки спермиев, v — влагалище, vs — семенной пузырек.
рт
ков. Этой точки зрения придерживается и Акс. По мнению Штейн- бека (Steinbock, 1924, 1966), связь бурсы с кишкой отражает первичные отношения женского полового аппарата и кишечника, через который первоначально выводились яйца. Бурсу он рассматривает как обособившийся задний участок кишки, взявший на себя функцию сохранения спермы партнера. Карлинг (Karling, 1940, 1963) считает, что бурсы турбеллярий могут возникать двумя способами: впячиванием эпителиев (эпителиальные бурсы) или преобразованием участка паренхимы, который специализируется для хранения спермы партнера (паренхимные бурсы). Генито- интестинальный нроток, по его мнению, образовался вторично. Рейзингер (Reisinger, 1961) полагает, что бурсы турбеллярий возникли несколькими путями. Он придерживается представления Штейнбека и думает, что связь бурсы с кишкой отражает первичные отношения в классе турбеллярий. Вместе с тем для ряда групп он считает возможным принять самостоятельное возникновение бурс по Карлингу. Выше мы постарались показать, что бурсы и генито-интестинальные связи имеют различную природу. Как видно из монографии Дорьеса (Dorjes, 1968), у Acoela бурсы раз
вивались многократно. Независимо и, очевидно, неоднократно возникали они также и у Prolecithophora, у Neorhabdocoela (как это хорошо показал Евдонин на Kalyptorhynchia) и у представителей других отрядов.
Акс не отрицает различной природы женских копулятивных органов турбеллярий, напротив, он отмечает, что бурсальные органы, без сомнения, не имеют единого происхождения, а генито- интестинальная связь может возникать вторично (Ах, 1961,
Рис.
86. Различные формы эпителиальных бурс
Neorhabdocoela.
(По:
Karling,
1963).
А
— Cheliplana
stylifera (Kalyptorhynchia),
Б
— Pseudograffllla
arenicola (Dalyellioida),
В—
Annulorhynchus
adriaticus (Kalyptorhynchia.), Г
—
Halammovortex
nigrifrons (Dalyellioida). af
— женский
антрум, am
— мужской
антрум, be
— копулятивная
бурса, dgi
—
ductus
genito-intestina- lis, gvd
— гермовителлодукт,
ov
— яичник,
ovd
— яйцевод,
pg
— половое
отверстие, rs
— семеприемник,
v
— влагалище.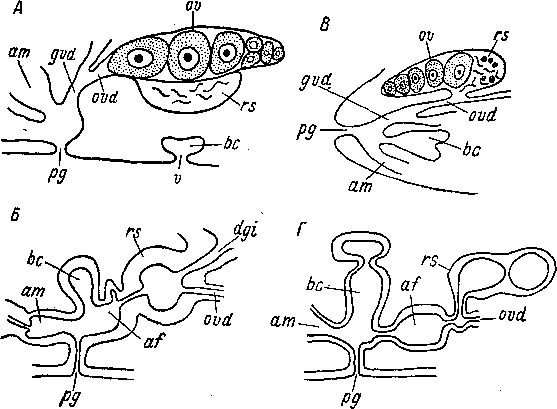
стр. 35). Однако он считает, что следует признать наличие первичного бурсального органа, который выводится от предков турбеллярий. Таким образом, ему приходится различать «первичные» бурсы, связанные друг с другом общностью происхождения, и «вторичные» бурсы, возникшие независимо. Однако, если имеется только «вторичная» бурса, а «первичная» отсутствует, то необходимо предположить довольно сложный процесс: редукцию «первичной» бурсы и замену ее «вторичной». Еще труднее представить исчезновение бурсы, когда не появляется орган, компенсирующий ее функцию. Как известно, при гермафродитизме вырабатываются механизмы, препятствующие самооплодотворению. К ним надо отнести и бурсы. Вряд ли в такой ситуации бурса могла оказаться ненужной и исчезла. Тем не менее случаи отсутствия женских копулятивных органов многочисленны, причем отнюдь не тольк
о
у Acoela. Трудно также представить, чтобы более совершенная «первичная» бурса с ductus spermaticus и влагалищем была заменена весьма несовершенной (например, «бурсальной тканью», о которой уже упоминалось). Столь же трудно объяснить морфологический ряд редукционных упрощений, который приходится строить, если выводить путем полимеризации от «первичной» бурсы Акса бурсоподобные органы представителей Oligochoerus и Anaperus. Все эти трудности отпадают, если принять возникновение и прогрессивное развитие женских копулятивных органов в пределах класса турбеллярий.
Мужские копулятивные органы
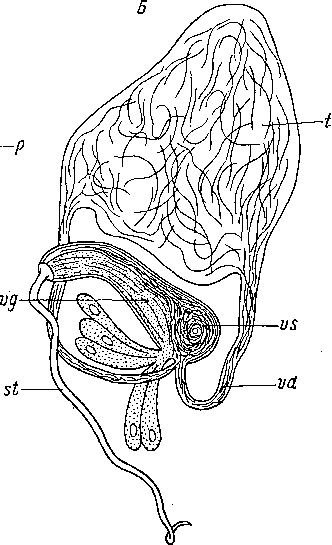
ат
Рис. 87. Совокупительный орган в виде мускулистого пениса (А) и кутикулярного стилета (.5).

Что касается мужских копулятивных органов, то ими обладают все турбеллярии. Разнообразие их чрезвычайно велико. В простейших случаях они представляют собой результат вворачивания по
-
А — Procerodes lobata (Tricladida), В—Haplovejdovshya subterranea (Neorhabdocoela).
(A — из: Beauchamp, 1961; Б — из: Luther, 1962). af — женский антрум, am — мужской антрум, be — копулятивная бурса, gc — скорлуповые железы, р — мускулистый пенис, st — стилет, t — семенник, vd — семяпровод, vg — пузырек зернистого секрета, vs — семенной пузырек.
кровов (рис. 80, Б—Е; 81). Как показал Карлинг (Karling, 1963), это простейший способ образования органов, функция которых связана с нападением на добычу или с копуляцией и сводится к захвату и прикреплению, разрыву покровов, проникновению
сквозь покровы и т. ir. Таким путем образуются и глотка, и мужской копулятивный орган, и хоботок калипторинхий. Далее перед всеми этими органами открываются три основные возможности развития: 1) образование выворачивающегося органа, 2) развитие мускулистого выдвигающегося органа, 3) кутикуля- ризация. В соответствии с этим мужские копулятивные органы

Рис.
88. Копулятивный орган Promonotus
schultzei
(Proseriata).
(По: Ах, 1956).
А — циррус в вывернутом состоянии; Б — циррус ввернут, семяизвергательный канал (dej) закручен в спираль.
развиваются как циррус (рис. 26; 84, А, Б; 85; 89, А; 90), мускулистый пенис (рис. 87, А) и кутикулярный стилет (рис. 2; 31; 33, А; 55, А; 83; 87, Б; 89, Б). Следует отметить, что во многих случаях развитие осложняется сочетанием нескольких тенденций. Например, циррус может приобретать кутикулярные шипы и крючья (рис. 88), а мускулистый пенис — стилет (рис. 25; 89, Б). Нередко в формировании копулятивного аппарата участвуют органы, уже имеющиеся у животных; к ним относятся железы и прежде всего органы кожного вооружения.
Напомним также, что процесс копуляции связан не только с переносом спермы, но и с раздражением партнера, которое,очевидно, играет существенную роль в обеспечении осеменений. Функция раздражения осуществляется самыми различными способами. В состав мужского копулятивного органа обычно входят специальные железы, секрет которых нередко изливается в особый резервуар (vesicula granulorum), который либо объединен с vesicula seminalis (рис. 25; 87, Б), либо представляет собой отдельный пузырек (рис. 33, А; 35; 48; 106). В ряде случаев образуются особые простатические органы (рис. 83; 89, А; 90). У Acoela, например, в качестве таковых используются грушевидные железы; раздражающую функцию у них осуществляют также железистые шипы, сагиттоцисты и иглы.
Сказанного достаточно, чтобы а 5
показать,
как велико разнообразие мужских
копулятивных органов турбеллярий.
Если же сравнить положение в разных
отрядах, то можно обнаружить весьма
различные картины. У представителей
некоторых отрядов мужские совокупительные
органы однотипны. Так, у Macrostomida
и
Рис. 89. Копулятивный аппарат представителей Monocelididae (Proseriata).
А — Minona evelinae, В — Monocelis fusca. (Из: Beauchamp, 1961). Ьс — копулятивная бурса, dgi — ductus genito-intestinalis, ovd — яйцевод, p — пенис, р/ — женское половое отверстие, р/с — женское кодулятивное отверстие, рт — мужское половое отверстие, vp — простатический пузырек, vs — семенной пузырек.
Lecithoepitheliata — это стилеты, у Prolecithophora — мускулистые пенисы с развитием складчатости. В других отрядах разнообразие очень велико. Особенно интересны в этом отношении Acoela. Если у Neorhabdocoela и Seriata разнообразие связано с развитием сложно устроенных копулятивных органов, то у Acoela оно состоит в наличии очень примитивных состояний и развитии от них в различных направлениях. Мы отметили три основных пути, по которым может идти эволюция мужских копулятивных органов, начиная от простейшего впячивания покровов. И подобное впячи- вание, и все три пути развития имеются у Acoela, причем такое разнообразие наблюдается только у них. Среди других турбеллярий совокупительный орган в виде простого вворачивания покровов имеется лишь у Plicastoma jenseni из Prolecithophora (Reisinger, 1934—1935; Karling, 1967).
Все основные типы совокупительных органов намечаются по существу уже у Haploposthia и близких видов. Рассмотрим для
Примера путь, ведущий к кутйкулярному стилет^. У представите* лей Haploposthia в ресничном трубковидном половом преддверии — антруме — развиваются мелкие кутикулярные палочки, очевидно представляющие собой оформленный секрет (Н. rubropunctata,
Н. monogonophora), затем они складываются в пирамидку, образующую уже простейший кутикулярный пенис (Н. rubra, Рага- proporus pellucidus). Далее кутикулярные палочки превращаются в настоящие спикулы, которые образуют стилет такого типа,
Рис.
90.
Копулятивный
аппарат Minona
cornupenis (Proseriata).
(По: Karling,
1967b).
bs
— семенная
бурса, i
— кишка,
о — рот, op
—
простатический орган, ovd
—
яйцевод, р — циррус, р/ — женское половое
отверстие, р/с — женское копулятивное
отверстие, ph—
глотка, рт
— мужское половое отверстие, st
—
стилет простатического органа, vs
—
семенной пузырек.
как у Paraproporus rubescens, Paraphanostoma submaculatum и Actinoposthia (рис. 91). Развитие кутикулярного стилета ведет к редукции антрума, в результате формируется орган, характерный для Paraphanostoma и Childia. Столь же четкие ряды развития прослеживаются и при образовании цирруса и мускулистого пениса.
Если о происхождении женских копулятивных органов можно с уверенностью сказать, что они у различных турбеллярий возникли независимо, то происхождение мужских копулятивных органов не столь ясно. Все турбеллярии уже обладают мужским совокупительным органом. Исходя из этого, можно сделать два предположения: 1) мужские копулятивные органы возникли независимо у разных турбеллярий, 2) они берут начало из одного источ- йика. Принимая во внимание огромное Многообразие в строеййй, положении и числе мужских копулятивных органов турбеллярий, мы вслед за В. Н. Беклемишевым (1964) склонны считать, что они самостоятельно появились в разных группах. Заметим, что

из этого заключения вытекает важное следствие: если мужской копулятивный аппарат действительно возникал в пределах данной группы и, стало быть, отсутствовал у первичных турбеллярий (женский, как мы уже видели, отсутствует и у многих ныне существующих форм), то это означает, что исходные способы осеменения у турбеллярий были весьма примитивными (псевдокопуйяция, наружно-внутреннее оплодотворение, возможно, уже с зачаточ-
ной копуляцией). Можно встать и на вторую точку зрения, однако разнообразие мужских копулятивных органов турбеллярий таково, что по существу нужно принять возможность полного их преобразования. Если даже первичные поликладиды и имели какой-то примитивный копулятивный орган наподобие выворачивающегося трубковидного антрума, то при наличии такого удобного материала, как грушевидные железы, могла произойти субституция функции. Аналогичные примеры у турбеллярий имеются. Так, сагиттоцисты, появившиеся у зеленых конволют, начинают использоваться при копуляции, мягкие пенисы при этом у некоторых видов редуцируются (см.: Мамкаев, 1971). Таким образом, придерживаясь второй точки зрения, также приходится признать независимое формирование мужских копулятивных органов у разных турбеллярий. В результате обе версии оказываются по существу близкими одна к другой.
В этом разделе мы представили материал, показывающий, что у турбеллярий встречаются очень примитивные способы осеменения, а бурсы и мужские совокупительные органы формируются в пределах группы. Совершенно очевидно, что спаривание при отсутствии женских копулятивных органов первично. Становление внутреннего оплодотворения идет различными путями. Как и при возникновении любой новой функции, здесь развитие идет во многих направлениях. Следует подчеркнуть, что при этом мо- , гут быть весьма разнообразные ситуации. Совокупительные органы возникают из разных источников, используются различные «подручные» материалы. Ресничные черви представляют ценный материал для исследования формирования различных систем, так как они дают многочисленные морфологические ряды, которые можно проследить от самых первоначальных состояний.
ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Особенности примитивного спирального дробления
Онтогенез турбеллярий за последние двадцать лет привлекал к себе большое внимание. Появились несколько крупных исследований и ряд обзорных работ, авторы которых пытались проанализировать эмбриональное развитие в соответствии со своими взглядами на происхождение Metazoa, в частности Тиг- bellaria.
Для нас особый интерес представляют статья О. М. Ивано- вой-Казас (1959а) «К вопросу о происхождении и эволюции спирального дробления», аналитическая глава об истории развития турбеллярий из работы Акса (Ах, 1961), высказывания В. Н. Беклемишева относительно онтогенеза турбеллярий (19636, 1964)
и экспериментальный анализ дробления у Acoela, осуществленный Бойер (Воуег, 1971).
Archoophora, лишенные еще желточников, имеют энтолеци- тальные яйца, дробление которых, естественно, сохранило еще первичные черты. Что касается Neoophora, то их экзолецитальные яйца откладываются в капсулах, также содержащих множество, иногда сотни и тысячи, желточных клеток. Поэтому эмбриональное развитие Neoophora подверглось значительным вторичным изменениям, сводящимся к выработке приспособлений к возможно раннему использованию зародышем желтка, заключенного в желточных клетках.
У Archoophora встречаются два варианта спирального дробления — квартетное с четырьмя основными бластомерами — макромерами, от которых на анимальном полюсе последовательно отделяются квартеты микромеров, и дуэтное — с двумя основными бластомерами. Первый вариант хорошо известен у Polycladida, второй очень характерен для A coela и в интересной форме проявляется также у Macrostomida.
Очень важно решить вопрос, какой из этих двух вариантов является исторически более ранней формой спирального дробления. По господствующим представлениям, общие предки Polycladida и Acoela обладали уже типичным квартетным дроблением, причем предполагается, что у первых оно сохранилось в почти неизменном виде, а у вторых дало начало дуэтному варианту (Remane, 1950; Шмидт, 1951; Hyman, 1951; Ливанов, 1955; Marcus, 1958, Богомолов, 1960; Ах, 1961; Федотов, 1966; Slewing, 1969). Это утверждение почти ни у кого не вызывает сомнений, принимается без доказательств, как нечто само собой разумеющееся, и речь идет главным образом о том, каким именно путем совершился эволюционный переход от квартетного дробления к дуэтному (см., например: Богомолов, 1960; Siewing,
1969).
Противоположное мнение, к которому присоединяемся и мы, было высказано Бресслау (Bresslau, 1909), развито О. М. Ивано- вой-Казас (1959а, 19596) и поддержано затем В. Н. Беклемишевым (19636, 1964). По этой концепции, все особенности квартетного спирального дробления в той форме, в которой они проявляются у Polycladida (не говоря уже о Polychaeta), возникли в результате длинного и сложного процесса эволюции онтогенеза из примитивных форм дуэтного спирального дробления.
В классической работе по развитию Convoluta roskoffensis Бресслау (Bresslau, 1909) пришел к выводу, что замечательное сходство между дуэтным дроблением Acoela и квартетным у Polycladida не случайно, но объясняется их гомологией и выражает близкое родство этих отрядов. По Бресслау, дуэтное дробление Acoela примитивнее. На это указывает все последующее развитие бескишечных турбеллярий, в котором нет никаких следов деградации или неотенического ускорения,
У Cnidaria дробление яйца очень разнообразно и изменчиво и нередко имеет признаки очень примитивного спирального дробления (Мечников, 1886; Wietrzykowski, 1912; Berg, 1941; N у holm,
1942—1944; Корсакова, 1949; Жинкин, 1951; Дондуа, 1952).
Эти признаки спиральности иногда проявляются начиная со второго деления и заключаются в очень характерном взаимном расположении первых четырех бластомеров двумя перекрещивающимися парами. Именно такое расположение характерно для Acoela и наблюдается, например, у гидроидной медузы Rathkea fasciculata (рис. 92; Мечников, 1886).

В г А
Рис.
92. Различные формы дробления у Cnidaria.
А
— анархическое дробление гидромедузы
Oceania,
В
— дуэтное спиральное дробление у
гидромедузы Rathhea
fasciculata,
вид
сбоку; В
— квартетное спиральное дробление у
нее же, вид с анимального полюса; Г и Д
— спиральное дробление морского пера
Funiculina,
вид
с анимального полюса (А—В
— по: Мечников, 1886; Г,
Д
—
по: Berg, 1941).
Разумеется, одно сходство в начальных стадиях дробления у таких форм, как Rathkea и Acoela, отнюдь не может свидетельствовать об их близком родстве. Вероятно, примитивное дуэтное дробление появлялось у первобытных Metazoa неоднократно,
и, скорее всего, случаи расположения дуэтами бластомеров у некоторых Cnidaria можно рассматривать только как модели, показывающие нам, как могли возникнуть особенности дуэтного дробления у предков турбеллярий. По-видимому, дуэтное дробление в той форме, как оно представлено у современных Acoela, не вполне примитивно, а имеет уже некоторые черты специализации, в частности дифференциацию бластомеров на макро- и микромеры.
Следовательно, вовсе не обязательно дуэтное дробление Acoela выводить из квартетного. Скорее, обе формы дробления произошли независимо от примитивного дуэтного дробления.Развитие Archoophora
Обычно рассмотрение эмбрионального развития турбеллярий начинают с изложения квартетного дробления Polycladida, как наиболее близкого к типичному спиральному дроблению Polychaeta. Мы отступим от этой схемы и начнем с анализа развития Macrostomida, которое отличается рядом примитивных особенностей, сочетающихся, правда, с некоторыми явно вторичными приспособлениями.
Квартетный тип спирального дробления у некоторых видов Macrostomum отмечен давно (Reisinger, 1923; Богомолов, 1949; Papi, 1953), но детали развития у Macrostomum appendiculatum 1 стали известны сравнительно недавно (Seilern-Aspang, 1957а).
По данным Зайлерн-Аспанга, первое деление яйца меридионально, а второе, слегка неравномерное, леотропно, так что образуются два макромера и два микромера (рис. 93, А). Их взаимное расположение очень характерно и полностью совпадает с дуэтным расположением четырех первых бластомеров у Acoela, а именно два микромера лежат на анимальном полюсе в промежутке между двумя вегетативными макромерами. Микромеры не соприкасаются друг с другом, тогда как макромеры прилегают друг к другу на вегетативной стороне зародыша.
Третье деление дексиотропно. После него у одних экземпляров М. appendiculatum развитие идет по дуэтному, у других — по квартетному типу в зависимости от количества желтка в яйцах; при менее обильных его запасах дробление принимает дузтный характер. В этом случае первый дуэт микромеров и его потомки оттесняются последующими дуэтами к анимальному полюсу зародыша (рис. 93, Б—Г). При квартетном типе развития первый дуэт микромеров остается рядом с двумя макромерами и вместе с ними составляет основной квартет макромеров (А, В, С, D), от которых в дальнейшем отделяются четверки микромеров (рис. 93, Д).
При квартетном варианте на стадии 16 бластомеров четыре макромера, богатые желтком и лежащие на вегетативной стороне, преобразуются в очень своеобразную эмбриональную оболочку — «желточную мантию», которая обрастает весь зародыш и в дальнейшем эмбриогенезе не участвует, так как служит лишь для питания (рис. 93, Д). При дроблении по дуэтному типу желточная мантия образуется только из двух макромеров и двух микромеров третьего дуэта.
Эти непонятные на первый взгляд явления можно объяснить только тем, что в дроблении Macrostomum проявляются еще очень примитивные отношения (Иванова-Казас, 1959а, 19596). Крайняя
неустойчивость и изменчивость дробления в данном случае весьма характерны. Другой, не менее примитивный признак заключается в недетерминированности дробления. Еще в начале дробления в яйце появляются два презумптивных зачатка — глотки и мозга и полового аппарата. Они представлены двумя скоплениями свободной от желтка, базофильной цитоплазмы, однако между расположением этих зачатков и первых плоскостей деления нет ни-
si
Рис.
93. Macrostomum
sp. (Macrostomida).
Дробление яйца. (По: Seilern-
Aspang,
1957а).
А
— дузтное дробление, стадия 4 бластомеров;
В
— то же, дексиотропное отделение второго
дуэта микромеров, стадия 6 бластомеров;
В
— то же, разделение бластомеров первого
дуэта, стадия 8 бластомеров; Г — то же,
леотропное отделение третьего дуэта
и разделение бластомеров первого и
второго дуэтов, стадия 16 бластомеров;
Д
— квартетный вариант дробления,
стадия 16 бластомеров. dt—<4——
бластомеры первого, второго и третьего
дуэтов, st
—
клетки основного дуэта (макромеры).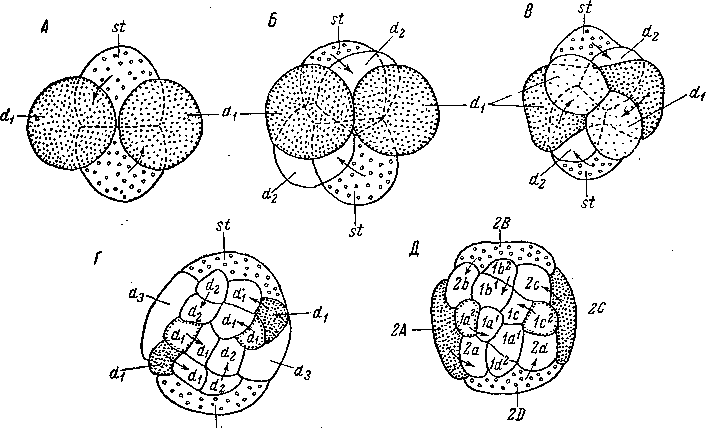
какой корреляции. После первого деления зачатки могут равномерно распределиться между двумя бластомерами или оба попадают в один из них, то в один, то в другой. На стадии четырех бластомеров у разных зародышей зачатки оказываются то в какой-либо одной клетке, то в двух, трех или во всех четырех. Другими словами, проспективное значение сходных бластомеров у разных зародышей варьирует в сильной степени, т. е. устойчивой детерминированности бластомеров еще нет. Об ее отсутствии свидетельствует и образование желточной мантии у разных зародышей из различных бластомеров (см.: Иванова-Казас, 1959а, 19596). Более того, оказывается, что при голодании Macrostomum откладывают яйца с уменьшенным запасом желтка, и в этом случае желточная оболочка вовсе не формируется.
тС другой стороны, способность зародыша Macrostomum образовывать зародышевую оболочку уникальна среди Archoophora и ее нельзя расценивать иначе, как раннюю вторичную специализацию.
Итак,
весьма неустойчивое недетерминированное
развитие Macrostomum
appendiculatum
представляет
собой очень примитивную форму
спирального дробления, из которого
легко выводится как настоящее дуэтное
дробление Acoela,
так и истинное квартетное дробление
других видов Macrostomum,
а также Polycladida
и
остальных Archoophora.
Таким образом, дуэтное дробление Acoela, по-видимому, не примитивнее квартетного, но оба возникли независимо от каких-то архаических форм дуэтного дробления. Если это так, то в развитии низших Spiralia (т. е. Plathelminthes и Nemathel-
Рис.
94. Примеры спирального дуэтного
дробления. (Из: Иванова-Ка- зас, 1959а).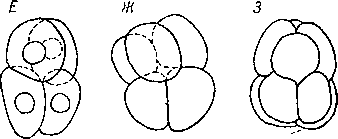
А, В — начальные стадии дробления»Соп- voluta (Acoela), В—Д — то же у Me- sostomaehrenbergi (Neorhabdocoela), Е—3— то же у Lepidodermella squamata (Gastro- tricha).
minthes) должно проявляться не только квартетное дробление, ной следы дуэтного дробления. И действительно, они имеются. О. М. Ива- новой-Казас удалось по литературным источникам обнаружить хорошие примеры дуэтного дробления у Neorhabdocoela (Mesostoma ehrenbergi — рис. 94, В—Д\ по: Bresslau, 1904), у Cestoidea (Taenia serrata, по: Janicki, 1907), у Gastrotricha {Lepidodermella squamata — рис. 94, E—3; no: Sacks, 1955) и у Monogenoidea (Gyrodactylus, no: Kathariner, 1904).
Судя по описанию Ланга (Lang, 1884), у многих Polycladida четырехклеточный зародыш в точности сходен с таковым у Macrostomum appendiculatum, Acoela и Rathkea. Два анимальных бластомера (А и С) имеют несколько меньшие размеры и вследствие лео- тропности второго деления помещаются над промежутком между двумя вегетативными макромерами- В viD (рис. 95). Позднее ани- мальные и вегетативные макромеры ложатся более или менее в одной плоскости.1Все дальнейшее развитие яйца уPolycladida,
как известно, 'носит характер Истинного Кйартетйбго спирального дробления. Впрочем, Ланг отмечает, что иногда, в виде исключения, у поликладид встречаются яйца, развивающиеся с образованием дуэтов.
Нужно сказать, что у некоторых Acoela дробление сохраняет еще большую неустойчивость, что вполне согласуется с представлением о примитивности дуэтной его формы. Так, по сообщению С. И. Богомолова (I960), некоторые яйца Convoluta convoluta испытывают квартетное дробление, тогда как ббльшая их часть
развивается обычным для Acoela способом; изредка у этого вида встречается даже отделение микромеров от одного макромера. Как показали прижизненные наблюдения М. А. Гу- реевой, во всех этих случаях мы имеем дело с нормальным развитием, в результате которого формируется подвижный червячок.
Высказывалось мнение, что дуэтное дробление Acoela произошло от квартетного развития путем «выпадения» второго меридионального деления. Как мы видели на примере развития Macrostomum, appendi- culatum и при внимательном сопоставлении четырехклеточных зародышей Acoela, Polycladida и Macrostomida, это утверждение лишено всяких оснований. Ни о каком выпадении деления не может быть и речи по той простой причине, что как при дуэтном, так и при квартетном спиральном дроблении второе деление всегда имеет более или менее ясно выраженный леотроп- ный характер. Первый дуэт микромеров у Acoela, возникающий вследствие леотропного деления, гомологичен не первому дексио- тропному квартету у Polycladida, а их макромерам А и С.
Итак, на основании изложенных выше фактов и соображений мы должны следующим образом представлять себе первоначальный тип дробления, свойственный общим предкам всех турбеллярий.
Это было очень примитивное спиральное дробление без дифференциации на макро- и микромеры. Второе клеточное деление уже было леотропным, и благодаря этому существовали явные признаки дуэтного типа. Развитие еще не было детерминированным, т. е. потенции отдельных бластомеров были очень широкими и судьба их легко менялась в зависимости от внешних и внутренних условий.
Примитивное спиральное дробление, очень близкое к этому гипотетическому типу развития, встречается, как мы видели, у некоторых Cnidaria. Зивинг (Siewing, 1969) называет его «псевдо-
епйрайьныМ», йМёй, очевидно, в виду не Только отсутствие характерных особенностей детерминированного дробления Polychaeta, но считая, что по своей природе оно вообще не имеет никакого отношения к истинному, спиральному дроблению. С нашей точки зрения, правильнее называть его вслед за О. М. Ивановой-Казас (1959а) «первично-спиральным дроблением».
Из него с одинаковой легкостью в процессе эволюции онтогенеза могли сформироваться обе формы специализированного детерминированного спирального дробления — дуэтная и квартетная.
Заслуживает внимания вопрос о природе процесса гаструляции у турбеллярий. Обычно принимается, что гаструляция осуществляется исключительно путем эпиболии, т. е. постепенного обрастания микромерами макромеров. Эпиболия была описана для Polycladida (Lang, 1884; Surface, 1907) и для Gnathostomulida (Riedl, 1969). Однако В. Н. Беклемишев (19636) совершенно прав, утверждая, что преобладающая роль принадлежит иммиграции клеток. В самом деле, на стадии, соответствующей бластуле, у Polycladida внутрь мигрируют некоторые клетки второго и предположительно третьего квартета и, наконец, все бластомеры четвертого квартета. Все эти клетки вместе с макромерами 4А—D дают начало фагоцитобласту, т. е. элементам энтодермы и мезодермы. Таким образом, у низших турбеллярий фагоцито- бласт возникает в значительной мере посредством мультиполярной иммиграции, хотя количество клеток, мигрирующих внутрь, невелико вследствие специализации и детерминированности дробления. Эти факты и соображения, естественно, приложимы и к другим Archoophora и чрезвычайно важны для укрепления гипотезы Фагоцителлы И. И. Мечникова и идеи о примитивности организации Acoela (Иванов, 1968).
Эмбриональное развитие у Acoela изучалось рядом авторов (Gardiner, 1895; Bresslau, 1909; Богомолов, 1960; Ах u. Dorjes, 1966; Apelt, 1969; Boyer, 1971; Богута, 1971, и др.). У Acoela в результате дуэтного дробления зародыш очень рано приобретает двулучевую симметрию (рис. 96). Гаструляция происходит путем погружения внутрь двух- макромеров, которые, размножаясь затем вместе с потомками четвертого дуэта микромеров, дают паренхиму. Дифференциации на энтодерму и мезодерму не происходит. Эти факты исключительно интересны, они вполне согласуются с представлением о большой примитивности Acoela и об изначальном отсутствии у них эпителизованной кишки и дают, следовательно, основание рассматривать паренхиму Acoela как малоизмененный фагоцитобласт.
Примитивность дуэтного дробления бескишечных ресничных червей недавно получила новое прекрасное подтверждение в экспериментальной работе Бойер (Boyer, 1971). Объектом ее исследования были дробящиеся яйца Childia groenlandica. Оказалось, что эмбрионы Acoela обладают очень высокой регулятивной способ
ностью, что, разумеется, свидетельствует об отсутствии детерминированности развития. В результате удаления одного из бластомеров на 2-клеточной стадии развития формируется нормальное животное, имеющее, однако, размеры, в два раза меньшие обычных. Нормальные особи формируются и при удалении части клеток или всех клеток первого, второго и третьего дуэтов. Однако
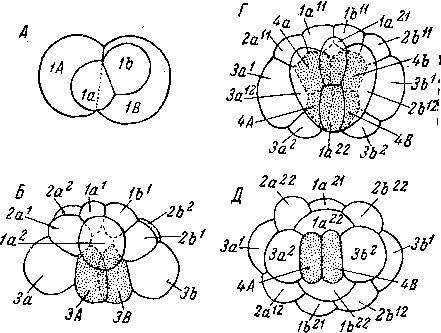
А — 4-клеточная стадия, с анимального полюса; Б — 12-клеточная стадия, сбоку; В — 16-клеточная стадия, сбоку (произошла иммиграция бластомеров ЗА и 3В); Г — 24-клеточная стадия, сбоку (отделение четвертого дуэта 4а и 46); Д — тоже, с вегетативного полюса; Е — 32-клеточная стадия, с анимального полюса.
при удалении у зародыша обоих макромеров получаются ненормальные животные в виде ресничных сферул; у них не развивается часть центральной пищеварительной паренхимы, на месте которой остается полость, и отсутствуют статоцист и рот. «Недетерминированность развития и высокая регулятивная способность у Acoela заставляют предположить, что этот отряд находится на уровне предковых форм Spiralia», — пишет Бойер (Boyer, 1971, стр. 97).
Замечательно далее, что у Acoela нет клетки, соответствующей бластомеру Ы аннелид и других «квартетных Spiralia», которая давала бы начало всей мезодерме и по крайней мере части энтодермы. Вместо этого «неясно выраженные у Acoela энтодерма и

Рис.
97. Planocera
inguilina
(Polycladida).
Некоторые стадии эмбрионального
развития. (По: Surface,
1907).
А
— 8-клеточная стадия, с анимального
полюса; В
— 16- клеточная стадия, с анимального
полюса; В
— оптический разрез, образование
эктомезрдермы; Г — гаструла, образование
стомодеума. Ы
— бластопор, end
—
энтодерма, тез
— мезодерма, г
— эктодерма, v
—
желток.
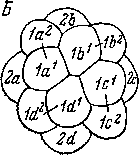
мезодерма происходят от всех клеток, мигрирующих внутрь во время гаструляции» (Воуег, 1971). В этом заключается еще одна чрезвычайно примитивная черта развития Acoela.
Некоторые авторы, особенно С. И. Богомолов (1960), настаивают на наличии у Acoela настоящей инвагинационной гаструлы с эпителизированной первичной кишкой. Собственные данные Богомолова о развитии Convoluta convoluta, однако, изложены неясно, документированы недостаточно и потому неубедительны. Как известно из очень обстоятельных работ Гардинера (Gardiner, 1895), Бресслау (Bresslau, 1909) и подтверждено самим Богомоловым, вся «энтодерма» Acoela в момент гаструляции состоит всего из двух макромеров (рис. 96, Б, В). Погружение их внутрь зародыша естественнее рассматривать как иммиграцию, а не инвагинацию. Мы уверены, что невозможно серьезно и без предвзятости
говорить о существовании первичной кишки,, состоящей из энтодермального эпителия, когда налицо всего две клетки, сохраняющие к тому же характер недифференцированных бластомеров.
У Polycladida мы находим уже вполне сложившееся квартетное дробление (рис. 97). Имеются бластомеры, гомологичные розетке и кресту аннелид, но, разумеется, не столь индивидуализированные, как у последних. Бластомеры, отвечающие трохобла- стам аннелид, однако, не участвуют в образовании ресничного шнура. Судьба бластомеров, образующих энтодерму у Polycladida, очень своеобразна. Из клеток четвертого квартета, гораздо более крупных, чем макромеры 4А—D, бластомеры 4а—с дегенерируют, распадаются и вместе с макромерами идут на питание зародыша. Возможно, впрочем, что у ряда поликладид они еще участвуют в формировании кишки (Selenka, 1881; Lang, 1884). Потомки бластомера Ы, как у других Spiralia, дают клетки средней кишки и первичные мезодермальные клетки. Эктомезенхима, представленная сначала четырьмя радиально лежащими клетками, отделяется от третьего квартета микромеров.
К примитивным чертам дробления у Polycladida следует отнести крупные размеры микромеров, отсутствие настоящих розетки, креста и трохобластов и гомоквадрантность. Кроме того, у поликладид первый соматобласт (2d), играющий такую большую роль в развитии кольчатых червей и моллюсков, не выражен: этот бластомер еще не отличается от других микромеров второго квартета ни по размерам, ни по участию в дальнейшем развитии. Впрочем, именно у Polycladida уже проявляется в зачаточной форме очень прогрессивная тенденция, характерная для высших Spiralia, а именно происходит некоторое увеличение квадранта!), связанное, очевидно, с возрастающим значением его в формировании туловища взрослого животного. За счет квадранта D развиваются ткани и органы дорсо-каудальной области тела и происходит разрастание задней части туловища во время онтогенеза. У аннелид эта роль квадранта D достигает своего максимального развития. Дегенерацию клеток четвертого квартета микромеров и макромеров следует рассматривать как раннюю вторичную специализацию развития.
У части поликладид (подотряд Cotylea), как известно, имеется характерная пелагическая личинка, открытая Иоганесом Мюллером (Miiller, 1850) и ныне называемая мюллеровской личинкой. Она имеет 8—10 выростов в средней части тела, покрытых длинными ресничками преорального ресничного шнура (рис. 98). Метаморфоз личинки носит эволютивный характер и заключается в постепенной редукции выростов и ресничного шнура и в усиленном росте в длину особенно посторальной части тела. Этот рост начинается очень рано, еще при формировании самой личинки и приводит к смещению ротового отверстия, лежащего первоначально у зародыша на заднем полюсе, на брюшную сторону тела.
Для других Archoophora, насколько теперь известно, характерен квартетный тип спирального дробления. Он отмечен у Catenulida (Stenostomum leucops, по: Bresslau, 1928—1933, и Catenula lemnae, по: Богомолов, 1949) и недавно описан у Gnathostomulida (Gnathostomula jenneri, по: Riedl, 1969). У последних дробление носит еще очень примитивные черты (рис. 99). Бластомеры А и С лежат в анимальной половине зародыша и соприкасаются; это указывает на леотропность второго деления. Бластомер D едва крупнее, чем остальные макромеры, однако после гаструляции в центральной части заро-
Рис.
98. Мюллеровские личинки поли*- кладид.
(Из: Beauchamp, 1951).
А — личинка Tysanozoon sp., сбоку; В — то же «Loboptera gargantua», с брюшной стороны. с — мозг, сг — ресничный шнур, I — лопасти, ос — глаза, sa — теменной султан, sp — задний султан.
А—3 — последовательные стадии развития.
1969).
дыша заметна пара больших клеток, которые, по мнению Ридля, представляют собой потомков бластомера Ы.
Развитие Neoophora
Среди Neoophora известны многочисленные случаи, когда развитие начинается еще как настоящее квартетное спиральное дробление. Таково начало дробления у Lecithoepitheliata, у некоторых Neorhabdocoela (Bresslauilla relicta и Phaenocora, по: Богомолов, 1949) и у ряда Proseriata (Otomesostoma auditivum, по: Reisinger, 1955; Monocelis fusca, no: Giesa u. Ax, 1964). Однако у большинства Neoophora, особенно у Tricladida, исчезают последние следы спирального расположения бластомеров, деление их с самого начала приобретает равномерный характер, дробление становится анархическим.
Ё развитии Neoophora можйо различать йесколько главных тенденций в выработке приспособлений для усвоения желтка из желточных клеток. В дальнейшем изложении мы в основном полагаемся на ту трактовку этих явлений, которую дает им О. М. Ива- нова-Казас.
У Lecithoepitheliata, представляющих большой интерес в филогенетическом аспекте, развитие изучалось у двух пресноводных форм — у Prorhynchus stagnalis (Steinbock u. Ausserhoffer, 1950) и Xenoprorhynchus steinbocki (Reisinger, 1970). У Prorhynchus stagnalis спиральное вначале дробление заканчивается образованием морулы. Замечательно, что в разное время года развитие Prorhynchus протекает различно. Летом развивающийся зародыш все время остается компактным, желточный материал отчасти поглощается его поверхностными клетками, но главная масса желтка сохраняется до конца развития и заглатывается уже вполне сформированным животным перед его вылуплением. Этот тип развития Штейнбек и Ойзерхофер вполне резонно считают более примитивным.
При низкой температуре развитие идет совсем иначе. Клетки раннего зародыша обрастают массу желточных клеток, которая при дальнейшем развитии оказывается в полости кишечника, где и переваривается.
Правда, по мнению Акса (Ах, 1961), в первом случае, т. е. при развитии плотного зародыша, происходят только модификацион- ные изменения в развитии эктодермы, а именно она формируется так рано и быстро, что под ней оказываются лишь немногие желточные клетки. Как бы то ни было, факт тот, что у Prorhynchus stagnalis имеются значительные вариации способа развития, существование которых становится понятным, если признать Leci- thoepitheliata группой, еще близко стоящей к основанию ствола Neoophora.
Недавно Рейзингер (Reisinger, 1970) сообщил о данных его ученицы Щискович (Szyskowitz), изучавшей эмбриональное развитее Xenoprorhynchus steinbocki. У этой турбеллярии яйца также окружены желточной клеточной мантией. Яйцо испытывает типичное спиральное квартетное дробление, сходное с таковым у Polycladida. Как у Archoophora, Sipunculida, у многих аннелид и моллюсков, у Xenoprorhynchus путем эпиболии образуется стерро- гаструла. Имеется небольшое преобладание квадранта D — особенность, намечающаяся уже у поликладид, но отчетливо выраженная у многих аннелид. На стадии около 40 клеток, сразу же после гаструляции, внутри зародыша выделяются два крупных бластомера — потомки 4d. Они отделяют от себя несколько мелких клеток, образуя нечто вроде коротких мезодермальных полосок, которые вскоре распадаются на мезенхиму. В отличие от Polycladida энтодермальные макромеры А—D и микромеры 4а—4с не резорбируются. Несколько мелких клеток, происходящих от второго и третьего квартетов, смещаются на периферию, где образуют эмбриональную оболочку. Вероятно, они отвечают клеткам провизорной эктодермы Tricladida, но в конце концов служат для захвата и переваривания желточных клеток.
Таким образом, для Xenoprorhynchus характерно сочетание очень примитивных черт эмбрионального развития Archoophora со вторичными провизорными приспособлениями, характерными для Neoophora. Это сочетание представляет, разумеется, исключительный интерес, так как очень важно с точки зрения установления родства Lecithoepitheliata с Acoela, именно с Hofsteniidae (Steinbock, 1966; Reisinger, 1970).
Эмбриональное развитие Paravortex gemellipara (Neorhabdocoela, Dalyellioida), судя по данным Болла (Ball, 1916), более всего напоминает примитивный вариант развития Prorhynchus stagnalis (с плотным зародышем). Сперва видны только два макромера и кучка микромеров. Позднее, на стадии морулы, различимы три сорта клеток (рис. 100). На одной стороне зародыша находятся мелкие клетки, потомки микромеров — мезо-эктодерма, на другой стороне лежат потомки одного из макромеров, дающие позднее первичную, или «желточную», энтодерму. Центр зародыша занят клетками — потомками второго макромера; из них образуется вторичная, или «кишечная», энтодерма. Энтодермальная часть зародыша соответствует будущей спинной стороне и обращена внутрь яйцевой капсулы, а будущая брюшная сторона обращена наружу. Клетки первичной энтодермы заглатывают желток и в свою очередь фагоцитируются клетками вторичной энтодермы. Значительная часть желтка, однако, остается вне зародыша. Тем временем поверхностные клетки мезо-эктодермы брюшной стороны уплощаются и разрастаются, образуя мезодерму и эктодерму спинной стороны (рис. 100, Б).
У других Neorhabdocoela, а именно у Mesostoma и Bothromeso- stoma, развитие больше похоже на развитие «полого зародыша» у Prorhynchus stagnalis. В летних яйцевых капсулах Mesostoma ehrenbergi (подотряд Typhloplanoida) желточные клетки, прилегающие со спинной стороны к зародышу, сильно вакуолизированы, тогда как периферические желточные клетки сливаются в синцитий. В массе эмбриональных клеток начинается дифференциация органов. Поверхностные клетки брюшной стороны дифференцируются в эктодерму и растут на бока и спинную сторону, обрастая при этом вакуолизированную часть желтка. Последний в конце концов попадает в полость кишки и здесь переваривается. Сходно протекает и развитие Plagiostomum girardi (Prolecithophora), но с тем, однако, отличием, что бластомеры зародыша слабо связаны друг с другом и сперва располагаются в массе желточных клеток, что напоминает дробление планарий (Bresslau, 1904).
К этому же типу в сущности относится и эмбриональное развитие Temnocephalida, изученное у Temnocephala fasciata (Haswell, 1909), Т. novae-zealandiae (Harrison, 1947) и Scutariella (=Caridi- nicola) indica (Fernando* 1934). У всех этих комменсальных турбел- лярий, близких к Neorhabdocoela, развитие протекает сходно. Желточные клетки вскоре сливаются в синцитий, а яйцо сначала испытывает полное, равномерное и беспорядочное дробление, а затем бластомеры дают зародышевую синцитиальную массу, которая отличается гомогенной и базофильной цитоплазмой. В ней рассеяны крупные и мелкие ядра (рис. 101, А).
Рис.
100. Paravortex
gemellipara
(Neorhabdocoela).
Эмбриональное развитие. (По: Ball,
1916).
А
— один зародыш, окруженный желточной
массой; В
— яйцевая капсула с двумя несколько
более старшими зародышами. ер
— эпидермальные клетки на поверхности
зародыша, та
— макромеры, mi
—
микромеры, nv
—
желточные ядра в вителло- фагах, vit
—
вителлофаги, vrn
—
желточная масса.
Постепенно в зародышевом синцитии дифференцируются зачатки мозга, глотки, кишечника, выделительного аппарата и половых органов (рис. 101, Б). Одновременно из зародышевого синцития выходят мелкие клетки, которые выползают на поверхность желточной синцитиальной массы и здесь дифференцируются в кожный эпителий, мышцы кожно-мускульного мешка и паренхиму. Желточная масса, таким образом, оказывается между стенкой тела и кишечником и здесь резорбируется.
У Seriata вторичные приспособления к усвоению желтка достигли максимального развития. Большой интерес прёдставляет развитие Bothrioplana semperi (Proseriata), изученное Рейзингером (Reisinger, 1923, 1940), так как оно еще близко стоит к типу развития, свойственному Prolecithophora и Neorhabdocoela. У Bothrioplana при дроблении обособляется часть бластомеров в виде «блуждающих клеток», которые выходят на поверхность желточной массы и здесь образуют провизорную оболочку. Под нею несколько позднее формируется дефинитивная эктодерма спинной
Рис.
101. Scutariella
indica
(Temnocephalida).
Эмбриональное развитие. (По:
Fernando,
1934).
А
— образование зародышевого синцития
с макро- и микромерами, В
—дифференциация зачатков органов
зародыша. Ы
— зародышевый синцитий, с —зачаток
мозга, end
—
зачаток кишки, ph
—
зачаток глотки, п
— ядро желточного синцития.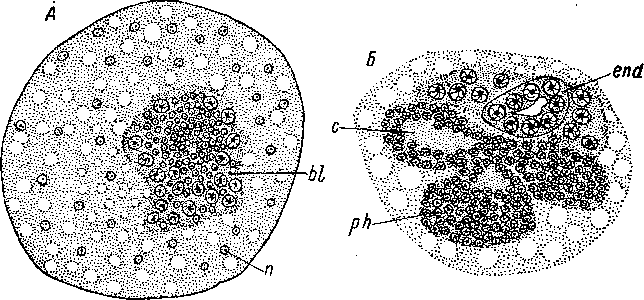
стороны тела. Из остальных клеток зародыша развиваются все внутренние органы. Таким образом, у Bothrioplana, так же как у Mesostoma, Plagiostomum и Temnocephalida, желток обрастается частью эмбриональных клеток.
Недавно стало известно и развитие Monocelis fusca (Proseriata) благодаря интересным исследованиям Гиза и Акса (Giesa и. Ах, 1964; Giesa, 1966). У Monocelis, как у большинства Proseriata, в яйцевой капсуле, кроме яйца, содержится множество (несколько сотен) желточных клеток. Деление яйца носит еще явные черты спирального дробления, на которых нельзя не остановиться (рис. 102). Второе деление леотропно, как у Acoela, и приводит к образованию двух вегетативных макромеров и двух несколько меньших клеток, сдвинутых на анимальный полюс. Позднее, как у Polycladida, Macrostomida и у многих других турбеллярий, первые четыре клетки ложатся в одной плоскости, составляя основной квартет макромеров. Первый квартет микромеров образуется при дексиотропном делении. Далее дробление утрачивает правильность. Развивается целобластула, которая скоро уплощается до степени двуслойного диска.
В конце дробления вследствие резко неравномерного деления многих бластомеров появляется множество мелких абортивных клеток, которые выталкиваются из тела зародыша и частью дегенерируют, а частью заползают в массу желточных клеток, где до времени пребывают в неактивном состоянии (рис. 103, А).
От
тела зародыша отделяется сперва 8, затем
через некоторое время еще 5 клеток,
которые образуют провизорную эмбриональную
оболочку (рис. 103, Б).
Одни ее клетки, называемые бластопораль-
ными, захватывают желточные и абортивные
клетки и проталкивают их внутрь под
оболочку, где те и другие постепенно
накапли-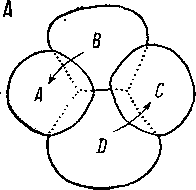

Рис. 102. Monocelis fusca (Proseriata). Дробление яйца. (По: Giesa, 1966).
А — 4-клеточная стадия, с анимального полюса (второе деление леотропное, как при дуэтном дроблении),
В — 8-клеточная стадия, с анимального полюса.
ваются в большом количестве. Другие клетки оболочки — вител- лофаги — заглатывают и переваривают желточные клетки, и, наконец, третьи образуют большую часть оболочки, часть их покрывается ресничками, с помощью которых зародыш начинает вращаться внутри яйцевой капсулы.
Оболочка не участвует в формировании тела животного, которое развивается из клеток эмбриона, или зародышевого диска, и вторично вошедших в его состав захваченных с желтком абортивных клеток. Зародышевый диск разрастается в промежутке между эмбриональной оболочкой и массой захваченных желточных клеток и постепенно обрастает всю желточную массу, образуя спинную сторону червя.
Таким образом, у Monocelis fusca в отличие от Bothrioplana semperi в начале развития полного обрастания желтка не происходит.
Сходным образом протекает развитие и у других Monocelididae (Сoelogyпорога, Archilopsis, по: Ах, 1961) и у Otomesostoma auditi- vum (Reisinger, 1955).
По развитию Tricladida опубликовано немало работ, причем первым исследователем эмбриологии планарий был И. И. Мечников (Мечников, 18776,1883; Mattiesen, 1904; Stevens, 1904; Fulinski, 1914, 1916, 1938; Seilern-Aspang, 1956, 1957b, 1957c, 1958; Moigne, 1963; Melander, 1963; Koscielski, 1966).
Для всех трикладид очень характерно наличие нескольких яиц и громадного количества желточных клеток в яйцевом коконе. Крайний пример в этом отношении представляет Dendrocoelum lacteum — в коконе содержится 20—40 яиц и 80—90 тысяч желточных клеток. Все или большая часть яиц развиваются в молодых животных.
Рис.
103. Monocelis
fusca
(Proseriata).
Эмбриональное развитие.
(По:
Giesa,
1966).
А
— выход из зародыша абортивных клеток;
В
— зародыш с эмбриональной оболочкой,
«бластопоральными» клетками и
вителлофагами. ап
— анимальный полюс зародыша, аЬ
— абортивные клетки, аЪс
— вновь образующаяся абортивная
клетка, Ъс
— «бластопоральные» клетки, се
— клетки зародыша, eh
—
эмбриональная оболочка, тз
— ядра вителлофагов, plv
—
цитоплазма вителлофагов, vc
—
заглоченные желточные' клетки.
Дробление у трикладид полное и равномерное, но беспорядочное, так как бластомеры с самого начала почти не связаны друг с другом (рис. 104, А, Б). Они лежат в желточном синцитии, образующемся в начале развития из желточных клеток, прилегающих к яйцу. Этот синцитий с заключенными в нем бластомерами представляет собой единую физиологическую систему (Seilern-Aspang, 1958).
Часть бластомеров концентрируется в середине желточного синцития, другая расползается в разные стороны и, достигнув внешних границ синцития, образует сперва сетевидный, а позднее сплошной слой провизорной эктодермы (рис. 104, В, Г). Скопление внутренних бластомеров, прилегающее в одном месте к провизорной эктодерме, развивается в провизорные глотку и кишечник. Глотка быстро достигает высокой гистологической дифференциации и характеризуется постоянством клеточного состава, что свидетельствует о значительной детерминированности всего развития. На этой стадии зародыш характеризуется четырехлучевой
симметрией. Главная его ось совпадает с осью провизорной глотки и отвечает дорсо-вентральной оси будущего червя. Скоро провизорная глотка начинает заглатывать желточные клетки, окружающие зародыш. Кишка переполняется ими, и весь зародыш сильно увеличивается в размерах. Далее между провизорной эктодермой и кишкой за счет размножения бластомеров, не принимавших
0X0
пет
![]()
Рис.
104. Эмбриональное развитие планарий
(Tricladida).
(Из
: Beauchamp,
1961).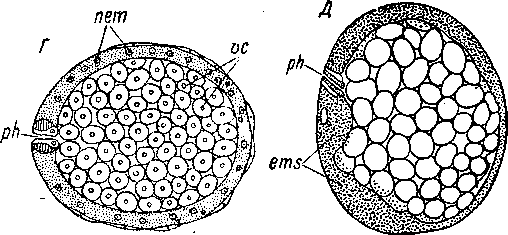
Ап В — стадии дробления Planaria torva, В — образование эмбриональной глотки у Dendrocoelum lacteum, Г — эмбрион, наглотавшийся желточных клеток; Д — резорбция желточных клеток и образование эмбриональной полоски, ес — провизорная эктодерма, ems — эмбриональная полоска, in — провизорная кишка, пет — ядра зародыша, ph — провизорная глотка, vc — желточные клетки.
пока участия в развитии, формируется зародышевая полоска, обладающая с самого начала билатеральной симметрией (рис. 104, Д). В ней дифференцируются три клеточные группы бластемы, из которых постепенно развиваются все дефинитивные органы. Провизорные органы дегенерируют, т. е. имеет место настоящий некробиотический метаморфоз.
Как справедливо подчеркивает О. М. Иванова-Казас, скрытая личинка Tricladida во многом напоминает зародыш Proseriata. Заманчиво принять, что все провизорные органы личинки планарий гомологичны эмбриональной оболочке Monocelis, поскольку большая часть ее клеток, по-видимому, соответствует провизор
ной эктодерме личинки. Гомологами провизорной глотки можно считать «бластопоральные клетки» у Monocelis, проталкивающие внутрь желточные элементы, а зачатком провизорной кишки — клетки-вителлофаги, способные к фагоцитозу. Однако все эти структуры у планарий состоят из большего количества клеток и функционируют несколько иначе.
Ранние стадии развития планарий подверглись еще большим вторичным изменениям, исчезли последние следы спирального дробления, еще очень полно представленные у Proseriata. Стадия бластулы выражена неясно. В общем, разумеется, развитие трикладид гораздо более специализировано, чем просериат. Акс выводит тип развития Tricladida (Ах, 1961) непосредственно из такового Proseriata, но правильнее считать, что оба типа сформировались в результате дивергентной эволюции от общего анце- стрального прототипа.
Подводя итоги, мы можем сказать, что у Neoophora явно намечается несколько основных тенденций в эволюции онтогенеза.
У одних форм, например у Prorhynchus из Lecithoepitheliata и у Paravortex из Neorhabdocoela, зародыш все время остается среди желточных клеток, которые тем или иным способом усваиваются всеми или только специально для этого предназначенными клетками. Зародыш прилегает к желтку своей спинной стороной, эктодерма которой образуется в последнюю очередь.
У других турбеллярий, именно у Lecithoepitheliata (при некоторых условиях развития у Prorhynchus), у Prolecithophora (Plagiostomum), у ряда Neorhabdocoela (Mesostoma, Bothromeso- stoma), у части Proseriata (Bothrioplana), зародыш! занимает в желтке периферическое положение, будучи обращен спинной поверхностью к желтку, и постепенно обрастает его клетками эктодермы.
У многих Proseriata (Monocelis, Сoelogyпорога, Archilopsis) приспособления для усвоения желтка развиваются в более своеобразной форме. Зародыш занимает в общем то же положение, но образует особую провизорную эмбриональную оболочку, одни клетки которой заглатывают желток, другие переваривают его, тогда как третьи образуют провизорные покровы.
От сходного, но более примитивного типа онтогенеза, по-ви- димому, возникло некробиотическое развитие Tricladida с характерной «несвободной личинкой».
ФИЛОГЕНИЯ TURBELLARIA Archoophora
Подробный анализ организации и развития Turbellaria, осуществленный нами, позволяет сделать некоторые заключения о родственных связях между отрядами. Из этого анализа прежде
А. В. Иванов и Ю. В. Мамкаев
161
Всего явствует, что Acoela представляют собой низшую группу среди Archoophora и что общий предок турбеллярий обладал аце- лоидным строением.
Acoela
Действительно, вся организация Acoela отличается глубокой примитивностью. Их внутренняя клеточная масса находится на уровне смешанной паренхимы с зачаточным тканеобразованием, как у самых примитивных многоклеточных (Trichoplax, Porifera). Эта паренхима с ее несовершенным разделением на периферический слой (потенциальную мезодерму) и центральную» пищеварительную часть (потенциальную энтодерму) чрезвычайно близка к первичному фагоцитобласту общих предков Metazoa. Среди турбеллярий мы находим все ступени обособления кишечника, причем начальные стадии этого филогенетического процесса свойственны Acoela, тогда как конечные стадии эпителизации кишки мы видим у Polycladida и у многих Macrostomida и Catenulida.
Покровы бескишечных турбеллярий отличаются следующими примитивными особенностями.
Слабое отграничение от паренхимы, что выражается в отсутствии базальной мембраны и резкой границы между эпидермисом и паренхимой (примитивный кожный эпителий Oligochoerus erythrophthalmus, немертиновый эпителий Nemertodermatidae, погруженный тип эпидермиса).
Присутствие эпителиально-мускульных элементов.
Образование различных органов кожного вооружения, которые в примитивных случаях отличаются своей множественной закладкой, неправильной локализацией и слабой дифференциацией, а при дальнейшей эволюции претерпевают резкую олигомеризацию и, частью меняя функцию, переходят на службу половому аппарату.
Первые этапы развития простых кожных сенсилл.
Бескишечным турбелляриям свойственны наиболее примитивные формы нервного аппарата.
Нервная система, целиком залегающая в толще эпидермиса в виде простого нервного плексуса, сравнимого с таковым у Cni- daria, иногда сочетающаяся с зачаточным эндоном.
Полупогруженная нервная система.
Целиком погрузившийся в паренхиму нервный ортогон, представленный различными ступенями дифференциации, вплоть до образования ортогонального мозга.
Большое количество одинаковых продольных нервных стволов ортогона (до 6 пар), которые у некоторых примитивных форм располагаются радиально, симметрично вокруг главной оси тела (D iopisthoporus).
Отсутствие или слабое развитие эндонального мозга.
Органы чувств также весьма примитивны. Особо следует подчеркнуть постоянное присутствие статоциста на переднем конце тела и развитие фронтального органа. Статоцист Nemertodermatidae отличается наличием не одного, а двух-трех и даже четырех лито- цитов (Sterrer, 1966), что скорее надо считать примитивной особенностью.
Очень примитивные отношения мы видим в половой системе.
Диффузно рассеянные в паренхиме, созревающие половые клетки, как мужские, так и женские (у некоторых форм), — особенность, характерная для самых примитивных Metazoa и, кроме Acoela, свойственная только Xenoturbellida и Porifera. Появление у большинства Acoela локализованных зачатковых зон половых клеток представляло следующий шаг в эволюции полового аппарата. Это состояние напоминает случаи предпочтительной локализации первичных половых клеток близ жгутиковых камер у губок.
Связанное с первой особенностью отсутствие tunica propria гонад и истинных половых протоков, как мужских, так и женских.
Отсутствие у многих Acoela дополнительных женских половых органов и женского полового отверстия.
Большое разнообразие примитивных типов мужского совокупительного органа, в частности таких его форм, которые, вероятно, гомологичны грушевидным железам и железистым шипам.
Наличие примитивных форм bursae seminales с кутикуляр- ным слоистым наконечником, напоминающих грушевидные органы.
К числу несомненно примитивных черт в организации Acoela относятся следующие.
Нестабилизированное в пределах отряда положение рта.
Отсутствие глотки у одних форм и слабое развитие pharynx simplex у большинства других.
Изначальное отсутствие протонефридиев.
Как мы видели, дуэтное дробление бескишечных турбеллярий относится к очень древним формам спирального дробления. Оно не более специализировано или вторично изменено, чем дробление Polycladida и Macrostomida.
Наконец, нельзя не отметить еще одну чрезвычайно характерную особенность Acoela. По сравнению со всеми другими отрядами они обладают наиболее неустойчивой пластичной организацией, обнаруживая необычайно широкий размах вариаций почти во всех важнейших морфологических признаках. Естественно эту особенность рассматривать как признак общей глубокой примитивности отряда.
Разумеется, Acoela не лишены и вторичных изменений организации. К их числу прежде всего надо отнести некоторую гистологическую и физиологическую специализацию паренхимы. Последняя лишь в редких случах сохраняет первоначальную способность фагоцитобласта к исключительно внутриклеточному пищеварению. Лишь у немногих форм (Convoluta distorta и Oxyposthia praedator) главная доля в пищеварении принадлежит амебообразным блуждающим фагоцитам. В большинстве случаев развивается особая лабильная форма пищеварительной паренхимы, внешне похожая на синцитиальное состояние. Пищеварение в ней, по-ви- димому, протекает не только в мелких пищеварительных вакуолях, но также и в более или менее обширных межклеточных лакунах и полостях.
Macrostomida
Macrostomida еще во многих отношениях близки к Acoela, а именно к семейству Convolutidae (Reisinger, 1933). Общие с последними признаки выражаются у них в простом строении покровов и кожно-мускульного мешка, в простом строении глотки — pharynx simplex и в общем характере полового аппарата, в частности в наличии у них двух раздельных половых отверстий — переднего женского и заднего мужского (впрочем, у некоторых форм — Acanthomacrostomum и др. — они могут быть вторично слиты). У макростомид имеется характерная семенная сумка, похожая на бурсу бескишечных турбеллярий (рис. 46). Яичники макростомид чрезвычайно сходны с таковыми Acoela. Мужской совокупительный орган макростомид, как у некоторых Acoela, построен по типу грушевидного органа. Характерно, что все эти признаки, общие лишь с Convolutidae, представляют собой самые примитивные особенности макростомид.
К прогрессивным чертам организации макростомид относятся следующие.
Стабилизация положения рта на переднем конце тела.
Простой эпителизованный кишечник с ресничным покровом и соответственно полостное пищеварение.
Значительная олигомеризация стволов нервного ортогона и концентрация нервного аппарата. От многочисленных продольных нервных стволов у макростомид осталась лишь пара боковых. Прецеребральный отдел ортогона полностью исчез. От эндональ- ного мозга вперед отходят только чувствительные нервы.
Редукция статоциста.
Развитие парных протонефридиев, снабженных примитивными циртоцитами и открывающихся наружу парой вентральных пор.
Напротив, эмбриональное развитие по крайней мере части макростомид сохранило еще очень примитивные особенности. Спиральное дробление настолько еще не стабилизировано и не детерминировано, что одинаково легко может протекать как по дуэтному, так и по квартетному типу (рис. 93). Однако образование желточной оболочки у эмбрионов Macrostomum appendiculatum представляет собой несомненно вторичную особенность.
В общем Macrostomida вопреки мнению Акса (Ах, 1961, 1963а) далеки от прототипа турбеллярий. Только некоторые особенности мх эмбрионального развития могут быть использованы для реконструкции общего прототипа.
Macrostomida — это слепая филогенетическая ветвь' Archo- 'opkora, отделившаяся от ацелоидных предков, близких к современным Convolutidae.
Gnathostomulida
Другой, близкий к Acoela, а следовательно, и к общему предку турбеллярий отряд представлен Gnathostomulida. По общему плану строения они мало чем отличаются от Acoela и Macrostomida. Очень примитивной их особенностью следует считать жгутиковый эпидермис, который резко отличает их от всех остальных турбеллярий. Однако эпителиально-мускульные элементы в нем уже отсутствуют. В числе других примитивных признаков нельзя не- упомянуть наличие, по меньшей мере у некоторых форм, интра- эпителиальной нервной системы и статоцистов в передней части тела и отсутствие протонефридиев (Riedl, 1969). Половая система гнатостомулид ничем в сущности не отличается от таковой у Acoela. Семенная бурса с наконечником в общем построена по типу грушевидного органа. Эмбриональное развитие — очень примитивное, гомоквадрантное, квартетное спиральное дробление (рис. 99).
К признакам ранней специализации и прогрессивного развития у гнатостомулид несомненно относятся следующие.
Развитие челюстного аппарата в глотке простого типа.
Эпителизация кишечника.
£ 3. Дифференциация поперечнополосатой мускулатуры в кожномускульном мешке и в глотке. Впрочем, таксономическое значение этого признака невелико, поскольку поперечнополосатая мускулатура возникает очень легко и независимо в самых различных группах Metazoa, известна она в том числе и у некоторых Acoela (Westblad, 1945; Иванов, 1952а).
Своеобразная организация спермиев у части гнатостомулид.
Нет сомнений, что Gnathostomulida представляют весьма обособленную ветвь Archoophora, очень рано отделившуюся от первобытных турбеллярий. Замечательная их особенность заключается в сочетании в их организации весьма примитивных признаков с чертами прогрессивного развития.
Catenulida
Обособленную группу Archoophora несомненно составляют и Catenulida. Главные примитивные их признаки заключаются в наличии статоциста, простой глотки, ортогональной нервной системы и фолликулярных семенников. Сюда же, кроме того, надо отнести наличие у некоторых форм эпителиально-мускульных клеток и ацелоидного синцитиального кишечника.
К ярким специфическим вторичным особенностям принадлежат эпителизованный ресничный кишечник (у большинства) и непарный медио-дорсальный протонефридий с задней или вентральной порой, а также непарные гонады и дорсальное положение мужского полового отверстия. Специализированными также следует считать циртоциты (у Stenostomum).
У катенулид, как известно, сильно развита способность к бесполому размножению путем паратомии (этот способ размножения свойствен также некоторым Acoela и Macrostomida). В связи с этим у катенулид наблюдается тенденция к редукции полового аппарата, приводящая у Rhynchoscolex к исчезновению мужских половых органов и к появлению партеногенеза (Reisinger, 1924; Marcus, 1945а; Ах, 1961, 1963а). Мы рассматриваем эти явления как вторичные изменения. Ближайшими родичами Catenulida среди Acoela предположительно считаются Nemertodermatidae (Steinbock, 1938).
Polycladida
В организации Polycladida сочетаются примитивные особенности с признаками высокого развития (Bresslau, 1928—1933). К первым относятся следующие.
Способность формировать множественные и неправильно рассеянные органы кожного вооружения и образование из них примитивных форм совокупительных и раздражающих органов мужского полового аппарата.
Сильное развитие нервного ортогона, состоящего из многочисленных стволов.
Слабое развитие протонефридиев.
Сильно расчлененные гонады. Этот признак, правда, можно связывать и с значительным увеличением размеров тела и, следовательно, рассматривать его как вторичную особенность (Ах, 1961).
К признакам вторичным и частью носящим прогрессивный характер принадлежит ряд особенностей.
Сильное развитие глотки складчатого типа.
Эпителизованный кишечник с ресничным покровом и полостное пищеварение.
Высокий уровень дифференциации тканей, сильное развитие пограничных опорных мембран, в том числе базальной мембраны эпидермиса. Резкое обособление кожного эпителия.
Сильное развитие эндонального мозга, его сравнительно высокая архитектоника. Развитие соединительнотканной капсулы вокруг мозга. Дифференциация внекапсулярных ганглиев (globuli).
Отсутствие комплекса фронтальных желез.
Редукция во взрослом состоянии статоциста и фронтального органа.
Высокое по сравнению с другими Archoophora развитие органов чувств.
Более специализированное, чем у Macrostomida и, вероятно, у Gnathostomulida, квартетное спиральное дробление.
Некоторые признаки Polycladida явно коррелируются с большими размерами тела, а именно сильное разветвление кишечника и усложнение ортогональной части нервной системы за счет развития дополнительных нервных стволов, их разветвлений и анастомозов. К этой категории признаков относятся и значительное усиление и усложнение кожно-мускульного мешка.
Поликладиды занимают довольно обособленное положение среди низших турбеллярий. Их невозможно связать с какой-либо определенной группой Archoophora и остается предположить, что линия эволюции, ведущая к Polycladida, очень рано обособилась от древнего комплекса ацельных предков. Впрочем, по мнению Штейнбека (Steinbock, 1924), имеется ряд признаков в мужском совокупительном аппарате и в яичниках, сближающих Polycladida с семейством Hofsteniidae среди Acoela.
Neoophora
Что касается Neoophora, то это искусственное объединение отрядов по одному признаку, а именно — по наличию желточников и соответственно — экзолецитальных яиц. Если бы удалось доказать, что желточники возникли в процессе эволюции один раз, то это говорило бы в пользу монофилетической природы Neoophora и давало бы повод к пересмотру статута этой группы как «Stadien- gruppe».
Однако, как мы видели, намечаются по крайней мере две независимые линии эволюции женской половой системы, приводящие к формированию желточников. Одна линия ведет от Acoela, именно от Hofsteniidae, к Lecithoepitheliata и, по мнению некоторых авторов, от примитивных представителей последних — к Neorhabdocoela. Другая линия ведет от Acoela к примитивным Prolecitho- phora.
Lecithoepitheliata
К этому отряду, кроме Prorhynchidae, Gnosonesimidae, первоначально причисляли и Hofsteniidae. Первый представитель Hofsteniidae — Hofstenia atroviridis — был описан Боком (Воск, 1923) и отнесен им к отряду A lloeocoela. Штейнбек первый обратил внимание на принципиальное сходство Нofstenia с типичными Асое- 1а, а значительно позднее Папи (Papi, 1957) на основании сравнительного изучения Hofstenia и открытого им нового рода — Hof- steniola — перенес Hofsteniidae в отряд Acoela. Это было совершенно правильное решение. Действительно, Hofsteniidae имеют погруженный эпидермис с эпителиально-мускульными клетками, интраэпителиальную нервную систему, статоцист с зачаточным эндональным мозгом, фронтальные железы с фронтальным органом, типичный ацелоидный пищеварительный синцитий, нерезко отграниченный от периферической паренхимы, диффузные семенники и яичники без оболочек и половых протоков. У Hofsteniidae отсутствуют дополнительные органы женского полового аппарата и женское половое отверстие, как у многих других Acoela. Бок рассматривал большинство этих признаков как вторичные упрощения и редукции, и за ним охотно следует Акс, но Штейнбек сразу правильно оценил их как чрезвычайно примитивные особенности, характерные для Acoela.
Оригинальными чертами Hofsteniidae следует считать переднее, почти терминальное положение рта, наличие большой мускулистой трубчатой глотки — pharynx simplex — и особенно локализацию мужского полового отверстия спереди, неподалеку от ротового (рис. 35). Впрочем, первые две особенности встречаются и у других Acoela (например, у Proporidae).
Недавно детальное сравнение организации Hofsteniidae и Prorhynchidae, осуществленное Штейнбеком (Steinbock, 1966) и дополненное затем Рейзингером (Reisinger, 1968), убедительно показало близость этих двух групп.
Они сходны прежде всего по плану строения, т. е. одинаковым расположением рта, глотки, кишечника, органов половой системы.
У Prorhynchidae рот терминальный и ведет в большую ротовую полость, которой нет еще у Hofsteniidae. Мужское половое отверстие у проринхид открывается в эту полость (рис. 33), а у гофсте- ниид рядом со ртом. Однако у наиболее примитивного представителя Lecithoepitheliata — Hofstenioplesia (=Prorhynchus) haswelli — ротовая полость еще совершенно зачаточная; несмотря на это, в нее уже впадает мужской половой аппарат.
Как мы видели выше, покровы Hofsteniidae и Prorhynchidae принадлежат к одному типу; у тех и других погруженный эпидермис с синцитиальной эпителиальной пластинкой сочетается с удвоенным кожно-мускульным мешком. У Hofstenioplesia, по многим признакам занимающей как бы промежуточное положение, покровы лишены еще базальной мембраны и обладают эпителиальномускульными клетками (см. также стр. 34).
Глотка Prorhynchidae относится к одной из разновидностей массивного типа (pharynx variabilis). Однако она настолько близка к простой трубчатой глотке Hofsteniidae, что ее происхождение от последней представляется очень правдоподобным (Steinbock, 1927, 1966; Reisinger, 1968). Свидетельством ее независимого происхождения от массивной глотки других форм служит и отсутствие настоящего глоточного кармана. В зачаточном состоянии глоточный карман появляется у Hofstenioplesia', он едва развит у Prorhynchus alpinus и Geocentrophora sphyrocephala и только у Xenoprorhynchus (высшего представителя Lecithoepitheliata) достигает заметного развития (Reisinger, 1968).
Кишечник Lecithoepitheliata обособлен и эпителизирован, но замечательно, что среди Acoela как раз у Hofsteniidae проявляются первые признаки оформления кишечника — Hofsteniola имеет уже кишечную полость, которая у видов Hofstenia приобретает более или менее постоянный характер.
Мужской совокупительный аппарат у гофстениид и проринхид сходен не только по положению среди других органов, но и в деталях строения, и это сходство при всем желании невозможно объяснить конвергенцией. В обоих случаях в состав его входят и семенной пузырек, и мешочек зернистого секрета, связанные с ко- пулятивным органом, направленным вперед и обладающим стилетом из длинных хитиноидных палочек (кроме Geocentrophora, у которой простой стилет крючкообразен).
Имеются, разумеется, и серьезные различия в организации Hofsteniidae и Prorhynchidae, но они выражаются главным образом в наличии у последних прогрессивных признаков и новообразований. Так, у них развиваются половые протоки и имеются дополнительные органы женского полового аппарата, открывающиеся женским половым отверстием вентрально (рис. 33). У проринхид появились протонефридии и исчез статоцист. Естественно, прогрессивно развита и нервная система с органами чувств, достигающими у высших представителей Lecithoepitheliata, особенно у Xenoprorhynchus, высокой дифференциации (рис. 71). Интересно, что еще у Hofsteniidae, именно у Marcusiolatinga, развивается полу- погруженный нервный ортогон (рис. 61; Du Bois-Reymond Marcus, 1957).
Особенно интересны эволюционные изменения яичников у сравниваемых форм. Для Lecithoepitheliata характерно, что созревающие в яичнике ооциты окружены фолликулярной оболочкой из желточных клеток (рис. 33, 82), причем яйца утратили способность вырабатывать желток, т. е. стали экзолецитальными, а фолликулярные клетки, напротив, потеряли способность к развитию, но накапливают желток и выделяют вещество яйцевой капсулы. У Hofsteniidae яйца энтолецитальные и желточных клеток нет. Hof- steniola pardii не имеет еще лецитоэпителиальных оболочек (Papi, 1957), но у Hofstenia atroviridis молодые ооциты окружены питающими клетками, частью заглатываемыми подрастающими ооцитами, частью образующими вокруг них фолликулярную оболочку. Желток накапливается как в ооците, так и в фолликулярных клетках; последние выделяют яйцевую оболочку (Воск, 1923). Питающие клетки, поглощаемые ооцитами и представляющие собой абортивные яйца, 'широко распространены у Acoela; они описаны, например, у Proporus venenosus (Graff, 1891), у видов Haplodiscus (Bohmig, 1895), у Oxyposthia praedator (Иванов, 1952a).
Представители второго семейства лецитоэпителиат — виды Gno- sonesima — отличаются очень интересным строением яичников и по особенностям формирования яиц занимают промежуточное место между Hofsteniidae и Prorhynchidae. По наблюдениям Рейзингера (Reisinger, 1926), у Gnosonesima antarctica ооциты образуют еще довольно много желтка, хотя в основном эта функция принадлежит уже фолликулярным клеткам.Таким образом, у Hofsteniidae и низших Lecithoepitheliata хорошо прослеживается процесс превращения питающих клеток, или абортивных яиц, в настоящие желточные клетки.
![]()



Е уЭ
Р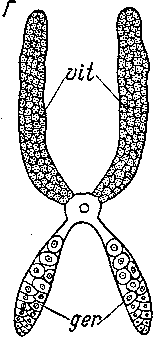

Рис. 105. Эволюция женских гонад у Neoophora по теории «тетрамерного яичника». (Из: Hyman, 1951).
Л — гипотетическое анцестральное состояние тетрамерного яичника; Б — тетрамерный гермовителлярий Gnosonesima, состоящий из желточных фолликулов, окружающих ооциты; В — обычный тип гермовителляриев с овариальной и желточной областями; Г — пара яичников и пара желточников; Д — один яичник и пара желточников; Е — гермовителлярий Prorhynchidae', Ж, 3 —другие типы гермовителляриев. ger — гермарии, gp — гонопор, f — фолликул, ov —яичники, or — овариальная область гермовителлярия’ vit — шелточники, vr — желточная область гермовителлярия.
Виды Gnosonesima очень интересны и в другом отношении. Они обладают «тетрамерной» женской гонадой (рис. 83), принимаемой Рейзингером и Бресслау за филогенетически исходное состояние, из которого развились как непарный вытянутый яичник с задней зародышевой зоной, свойственной Prorhynchidae, так и вс
еостальные типы гермовителляриев, гермариев и вителляриев турбеллярий (рис. 105). Однако, так как низшим Prolecithophora свойствен совсем иной тип примитивной женской гонады, то, очевидно, теория «тетрамерного яичника» за пределами Lecithoepitheliata -неприложима.
В последнее время монофилетическое единство Lecithoepitheliata оспаривается Карлингом (Karling, 1968). К этому отряду ныне относят только два семейства — Prorhynchidae и Gnosonesimi- dae — с совершенно различным строением глотки — первые обладают pharynx variabilis, вторые — ph. coniformis. Эти различия, по мнению Карлинга, настолько серьезны, что единое происхождение отряда исключено. Lecithoepitheliata, по Карлингу, искусственная группа.
Это заключение, однако, кажется нам преувеличенным. Несмотря на все различия в строении глотки, оба семейства следует производить от общего предка, которого можно представить себе как очень примитивного представителя Lecithoepitheliata, очень близкого к Prorhynchidae. Эта форма могла обладать еще примитивной глоткой (pharynx simplex), из которой выводится глотка современных Prorhynchidae и от которой мог произойти, как полагает и сам Карлинг, и специализированный глоточный аппарат Gnosonesimidae. Вероятно, и «тетрамерные» яичники следует приписать этому гипотетическому предку.
Prolecithophora и другие отряды
Как ни соблазнительно производить неоофорных турбеллярий от примитивных Lecithoepitheliata с «тетрамерными» яичниками, от этой концепции приходится отказаться из-за наличия среди Prolecithophora многих чрезвычайно примитивных форм с диффузными гонадами и желточниками, по организации стоящих еще на уровне примитивных Archoophora (Karling, 1940). Среди этих форм особенного внимания заслуживают Prolecithoplana lutheri (подотряд Combinata) и Plicastoma jenseni (подотряд Separata).
Prolecithoplana (семейство Cylindrostomidae) — небольшая (около 1.5 мм) морская турбеллярия, имеет антеро-дорсальную общую зачатковую зону гонад, как у многих Acoela; от нее берут начало не только мужские и женские половые клетки, но и желточные. Все эти клетки перемешаны и развиваются в паренхиме, не будучи объединены в семенники, яичники и желточники (рис. 106). Спермии пробираются через паренхиму к округлому семенному пузырьку, который открывается в совокупительный орган, состоящий только из маленького сферического мешочка зернистого секрета. Яйца и желточные клетки ползут сквозь паренхиму к железистому общему женскому протоку (ductus communis). Общее половое отверстие открывается во влагалище (карман) глотки, как у всех Combinata. Ротовое отверстие на переднем конце тела. Эпидермис Prolecithoplana представлен погруженным эпителием; рес
ничная глотка складчатого типа ведет в мешковидную кишку, лишенную еще tunica propria; мозг плохо отграничен от паренхимы; от него отходят четыре пары продольных нервных стволов; хорошо развиты фронтальные железы.
Plicastoma
jenseni,
тоже
небольшая (до 1 мм) морская турбел- лярия,
обнаруженная у берегов Гренландии, была
описана Рейзингером под родовым
названием Proporoplana
(Reisinger, 1934—
1935). Так как все изученные Рейзингером
особи были лишены желточных клеток,
то он отнес ее к Archoophora,
выделив в особый отряд, получивший
позднее название Proplicastomata.
Рейзингер
писал, что Proporoplana
по
организации настолько еще близка к
Acoela,
что
только такие прогрессивные черты,
как наличие складчатой глотки,
обособление кишечника от паренхимы
и крупный эндональный мозг заставляют
его выделять эту форму в самостоятельный
отряд.
Недавно Карлинг изучил особи этой турбеллярии, собранные Вест- бладом у побережья Исландии, и обнаружил, что в половозре-
Рис. 106. Prolecithoplana lutheri (Prolecithophora)i. Схема организации, вид слева. (По: Karling, 1940).
at — половой антрум, с — мозг, gf — фронтальные железы, in — кишка, о. — ротовое отверстие, od — общий женский проток, oes — пищевод, оо — ооциты, pg — половое отверстие, ph, — глотка, sp — сперматоциты, vc — желточные клетки, vg — пузырек зер! нистого секрета, vs — семенной пузырек.
лом состоянии они обладают желточными клетками, которые образуются с некоторым «запозданием», что нередко бывает среди Prolecithophora (Brandtner, 1934), и, судя по основным признакам, относятся к Prolecithophora Separata, именно к роду Plicastoma из семейства Plagiostomidae. Отряд Proplicastomata, таким образом, перестал существовать, а значение Plicastoma jenseni как «промежуточного звена» между низшими Archoophora и Prolecithophora значительно возросло.
Рейзингер отметил следующие черты строения Plicastoma jen- seni, общие с Acoela.
Примитивный погруженный эпидермис с эпителиальномускульными клетками.
Ацелоидное строение кишечника — отсутствие tunica propria, синцитиальная структура стенок кишки, пищеварение, протекающее исключительно в пищеварительных вакуолях.
Сильное развитие фронтальных желез и фронтального органа.
Отсутствие протонефридиев.
Ацелоидная половая система — яичник с парными зачатковыми зонами, без оболочки, яйцевода и дополнительных женских органов; отсутствие женского полового отверстия; диффузный семенник без семяпровода; мужской совокупительный орган в виде невооруженного пениса на заднем конце тела, открывающийся в терминальный мужской антрум.
Типичный нервный ортогон, состоящий из трех пар продольных стволов, залегающих еще в толще эпидермиса.
К прогрессивным особенностям Plicastoma jenseni принадлежат упомянутые выше pharynx plicatus, очень крупный эндональ- ный мозг и обособленность кишки, которая к тому же имеет уже постоянную полость.
Итак, Prolecithoplana и Plicastoma служат хорошими моделями, поясняющими, каким образом совершался эволюционный переход от древних ацелоидных форм к Prolecithophora, которых Карлинг (Karling, 1940) вполне резонно считает центральной группой среди Neoophora. Оба подотряда Prolecithophora — Combinata и Separata — произошли, по его мнению, от общих Prolecithoplana-образных предков. От них же, по-видимому, возникли Seriata и примитивные Neorhabdocoela. Эволюция гонад в этих филогенетических линиях иллюстрируется Карлингом на целом ряде модельных примеров, представление о которых дает его схема (см. рис. 107). Кроме двух форм, рассмотренных нами выше, диффузными половыми железами обладают Plicastoma norvegicum (рис. 107, Г) и Plagiostomum torquatum (рис. 107, Д). Но уже среди Combinata появляются формы с компактными семенниками и гер- мовителляриями — Enterostomula из Cylindrostomidae, Мопо- ophorum (рис. 107, Е) и Pseudostomum (рис. 107, Ж) из Pseudostomi- dae — и даже с компактными гермариями и вителляриями, например Pseudostomum quadrioculatum и Hypotrichinidae (рис. 107,
; эти формы по половой системе уже сходны с Neorhabdocoela.
Среди рабдоцелид самые примитивные формы, по мнению Карлинга, следует искать в занимающем центральное положение подотряде Typhloplanoida, именно в семействе Solenopharyngidae. Некоторые его представители, например Autopharynx vaginatus, по ряду признаков близки к Cylindrostomidae из Prolecithophora Combinata. Это мнение оспаривается Аксом, который думает, что Typhloplanoida скорее всего имели общих предков с Proseriata (Ах, 1961, 1963а). От Typhloplanoida одна ранняя боковая ветвь ведет к подотряду Dalyellioida\ от последних, вероятно, произошли комменсальные Temnocephalida (Karling, 1956с; Boettger, 1957) и Udonellida (Иванов, 19526). Подотряд хоботковых турбеллярийРис. 107. Схема происхождения и эволюции различных типов гонад. (По: Karling, 1940, с незначительными изменениями).
А — исходная единая дорсо-ростральная мужская и женская половая зачатковая зона; В — раздельные мужская и женская половые зачатковые зоны; В — тип Prolecithoplana lutheri, единая дорсо-ростральная зачатковая зона спермиев, яиц и желточных клетон; Г — тип Plicastoma norvegicum, наиболее примитивный тип среди Plagiostomidae (Рго- lecithopkora Separata) со сравнительно дорсальными зачатковыми зонами спермиев и яиц и с вытянутыми вдоль тела желточниками; Д — тип Plagiostomum torquatum — более развитый тип среди Plagiostomidae со сравнительно вентральными зачатковыми зонами семенников и яичников; Е — тип Enterostomula и Monoophorum (Cylindrostomidae из Prolecithophora Combinata), раздельные, снабженные оболочкой семенники и гермовителлярии, овариальная область еще непарна и лежит медио-дорсально; Ж — тип Pseudostomum hlostermanni (Cylindrostomidae) с парными, периферически лежащими (вентральными) вытянутыми овариальными областями гермовителляриев; 3 — тип Hypotrichina tergestina и Pseudostomum quadrioculatum (Hypotrichinidae и Cylindrostomidae из Prolecithophora Combinata) и многих Neorhabdocoela с проксимально отчлененными гермариями; И — тип многих Neorhabdocoela с полностью самостоятельными гермариями и вителляриями; К —■ тип Scleuraulophorus cephalatus (Cylindrostomidae) с овариальными областями в задних участках гермовителляриев.
(Kalyptorhynchia)
— специализированная
в некоторых отношениях группа, стоящая
довольно изолированно; ее сближают с
Typhloplanoida
(Graff, 1904—1908;
Bresslau,
1928—1933).
Классы паразитических плоских червей.
Рис. 108. Филогенетические отношения между отрядами и подотрядами турбеллярий. (Ориг.).
Porifera — -ГТ
Phagocytella
Что касается Seriata, то подотряд Proseriata содержит много примитивных форм, которые, вероятно, были филогенетическими предшественниками Tricladida (Ах, 1961, 1963а).
Филогенетические взаимоотношения между отрядами и некоторыми подотрядами турбеллярий показывает схема на рис. 108.
МОРФОЛОГИЧЕСКИЙ ТИП, ИЛИ ПРОТОТИП, TURBELLARIA
Весь фактический материал, рассмотренный нами выше, позволяет сделать попытку реконструировать общий план строения прототипа всех турбеллярий. Мы уже знаем, что ближе всего к общему предку ресничных червей стоят современные бескишечные турбеллярии. Однако среди Acoela есть немало вторично измененных в том или ином отношении форм. Так, например, текучее синцитиальное состояние пищеварительной паренхимы, как и резкое ее обособление от периферического слоя паренхимы у многих ви
дов, следует рассматривать как вторичную специализацию. То же относится и к переднему и, по-видимому, даже к вентральному положению рта, а также и к сильному развитию трубчатой глотки у некоторых форм.
По
нашему мнению, прототип Turbellaria,
который,
вероятно, был и общим предком других
низших Protostomia,
должен
быть’наделен следующими чертами
^организа- ции.
Чисто клеточным строением, т. е. [отсутствием вторичных синцитиальных структур.
Жгутиковым эпидермисом, нерезко отграниченным от паренхимы, без базальной мембраны.
Кожно-мускульным мешком, состоящим из миофибрилл эпителиально-мускульных клеток.
Массой разнородных внутренних клеток, обладающей свойствами смешанной паренхимы, с блуждающими амебообразными пищеварительными
Рис. 109. Реконструкция прототипа Turbellaria. (Ориг.).
al — пшца, ср — клетки паренхимы, dvm — дорсо-вентральное мускульное волокно, ер — эпидермис, / — жгуты, gep — кожная железа,
gf — фронтальные железы , гт — интраэпителиальные миофибриллы, п — нервный плексус, о — ротовое отверстие, оо — ооциты, oog — оогонии, ph с — пищеварительные клетки, pst — пора статоциста, sp — спермин, spg — сперматого- нии, st — статоцист, stl — лито- ЦИтЫ.
клетками — фагоцитами, играющими основную роль в пищеварении.
Простым ротовым отверстие^ на заднем конце тела, лишенным глотки.Лежащей в толще эпидермиса нервной системой в виде нервного плексуса.
Комплексом фронтальных желез.
Наличием статоциста с несколькими литоцитами на переднем конце тела.
Наличием фронтального органа.
Отсутствием протонефридиев.
Диффузным гермафродитным половым зачатком.
Диффузно рассеянными в паренхиме развивающимися мужскими и женскими половыми клетками. Отсутствием оболочек гонад и половых протоков.
Отсутствием мужского копулятивното аппарата.
Отсутствием женских дополнительных половых органов и женского полового отверстия.
Выведением зрелых оплодотворенных яиц наружу через ротовое отверстие или через разрывы стенки тела.
Наружным оплодотворением.
Примитивным, недетерминированным спиральным дроблением яйца дуэтного или квартетного типа.
Общий план строения этого гипотетического организма изображен в сагиттальном разрезе на рис. 109.
ПРОИСХОЖДЕНИЕ TURBELLARIA И ИХ ПОЛОЖЕНИЕ В СИСТЕМЕ
Представления различных авторов о происхождении турбеллярий сильно расходятся (см. таблицу). Из них прежде всего следует отбросить гипотезу целлюляризации, или полиэнергидную теорию (Steinbock, 1937, 1952, 1958; Hadzi, 1944, 1958, 1963; Hanson, 1958, 1963).
Эта концепция, по которой примитивные турбеллярии берут начало от многоядерных инфузорий, явно не подтверждается фактами; она подверглась серьезной критике со стороны Ремане, Иегерстена, Акса и Гайман (Remane, 1958, 1963; J&gersten, 1959; Ах, 1961; Hyman, 1964) и недавно подробно проанализирована
А. В. Ивановым (1968).
Все остальные гипотезы делятся на две основные группы. К одной относятся концепции, рассматривающие турбеллярий как деградированных потомков целомических сегментированных животных. Гипотезы другой группы считают ресничных червей низшим классом Bilateria, произошедшим от еще более примитивных Metazoa и давшим начало многим таксонам Protostomia.
А. В. Иванов и Ю. В. Мамк^ед
177
Классификация гипотез происхождения Turbellaria
Турбеллярии произошли от многоядерных инфузорий путем целлюляризации.
Полиэнергидная теория (Steinbock, 1937, 1952, 1958; HadZi, 1944, 1958, 1963; Hanson, 1958).
Турбеллярий произошли от трохофо- рообразных личинок олигомерных сидячих Coelomata.
Турбеллярии произошли от многоклеточных животных.
А. Предками турбеллярий были метамер- ные Coelomata. (Теории регрессивной эволюции турбеллярий).
Б. Предками турбеллярий были примитивные Meta- zoa. (Теории прогрессивной эволюции турбеллярий).
Архицеломатная теория (Alvarado, 1955, 1956, 1959).
Турбеллярии произошли от полимерных аннелид.
Аннелидная «теория (Van Beneden, 1891; Заленский, 1907; Lameer'e, 1910—1911, 1931; Boas, 1917; Naef, 1931; Brien, 1961, 1969).
Турбеллярии произошли от олигомерных Coelomata.
Теория Spiralia (Heider, 1914; Remane, 1951, 1954, 1959; Marcus, 1958; JSgersten, 1959; Ax, 1960, 1961; Slewing, 1969).
Турбеллярии произошли от Ctenophora.
Ктенофорная теория (Lang, 1884; Wilhel- mi, 1913; Догель, 1954).
Турбеллярии произошли от гипотетического Protrochozoon.
Трохофорная теория (Hatschefe, 1888).
Турбеллярии произошли от гастрееоб- разных Coelenterata.
Гастреадная теория (Karling, 1940; Ливанов, 1945, 1955; Федотов, 1966).
Турбеллярии произошли от плануло- образных предков.
Планулоидная теория (Graff, 1891, 1904; Беклемишев, 1937, 1944, 1952; Hvman, 1951; Hand, 1963).
Турбеллярии произошли от неотени- ческих планул Cnidaria.
Планулоидная теория Беклемишева (Беклемишев, 19636, 1964).
Турбеллярии произошли от фагоцител- лообразных предков Metazoa.
Ацельная теория Мечникова (Мечников, 1886; Иванов, 1968; Salvini-Plawen, 1969; Godreanu, 1970)
.
Общая характеристика гипотез происхождения турбеллярий от Coelomata
Остановимся сперва на гипотезах первой группы. Основаниями для них служат следующие предположения и соображения.
От Plathelminthes, как от животных с внутренним оплодотворением и гермафродитной половой системой, нельзя вывести остальных Bilateria с их наружным оплодотворением и раздельнополостью, тогда как обратный переход вполне возможен
.Паренхима турбеллярий представляет собой редуцированный и дезинтегрированный гомолог целома анцестральных ме- тамерных Coelomata, так как оба образования в онтогенезе развиваются из гомологичных первичных мезобластов (потомков бластомера 4 d).
Следовательно, первичными Bilateria могли быть только целомические метамерные животные с трубчатым кишечником, заканчивающимся анусом. От них произошли Plathelmlnthes и другие амерные черви, претерпев редукцию целома, исчезновение метамерии и другие значительные упрощения организации.
Объяснить происхождение первобытных целомических Bilateria — Procoelomata — непосредственно от низших Metazoa пытались многие авторы. Седжвик (Sedgwick', 1884) первый создал цикломерную энтероцельную теорию, по которой аннелиды выводятся непосредственно из организации кораллового полипа, перешедшего к ползанию на ротовой поверхности тела. Э. Ван Бенеден (Van Beneden, 1891) развил эту идею; в качестве предка аннелид он избрал обитающих в грунте Ceriantharia с их единственной зоной роста мезентериальных перегородок — между гастральными камерами, которую он сравнил с задней зоной роста сегментов у Polychaeta. Ему последовали Ламер (Lameere, 1910—1911, 1915—1916, 1931) и Бриан (Brien, 1961, 1969). Сходную концепцию — теорию Кораллулы — разработал Ремане (Remane, 1950). Наконец, Иегерстен (Jagersten, 1955) предложил выводить Procoelomata от несколько усложненной Билатерогастреи, предполагаемого им гипотетического предка всех Metazoa.
Все эти гипотезы принимают энтероцельное происхождение целома и все (кроме теории Билатерогастреи) выводят сегментацию из цикломерии Anthozoa. Кроме того, обычно их связывают с признанием теории Гастреи Геккеля, хотя непременная логическая связь с последней отсутствует.
Перечисленные аргументы в пользу первичности Coelomata чрезвычайно серьезны и требуют самого внимательного рассмотрения.
(Действительно, среди современных турбеллярий мы не имеем форм с наружным оплодотворением, если не считать за турбелля- рию Xenoturbella bocki. Однако, как отмечалось выше, выделение Xenoturbellida в качестве особого класса Plathelminthes носит условный характер. Xenoturbella во всяком случае остается формой, еще очень близкой к Acoela и к первобытным ацелоидным турбел- ляриям. Она обладает наружным оплодотворением и полностью лишена совокупительных органов. Спермии у Xenoturbella имеют еще характер обычных сперматозоидов с головкой, шейкой и хвостовым жгутом. Очевидно, все эти черты первичны и свидетельствуют о том, что общие предки Turbellaria и Xenoturbella откладывали яйца в воду, где они и оплодотворялись.
Кроме того, мы видели (стр. 120), что средипримитивных турбеллярий распространены весьма несовершенные формы спаривания, свидетельствующие о том, что внутреннее оплодотворение и копуляция действительно сформировались уже в пределах класса ресничных червей.
Таким образом, видимо, ничто не препятствует выведению билатеральных беспозвоночных с наружным оплодотворением от вымерших, очень примитивных общих предков Turbellaria и Хепо- turbellida. Выведение же Turbellaria (и других амерных Bilateria) из сегментированных Coelomata наталкивается на непреодолимые препятствия. Главное из них — невозможность объяснить ацелию бескишечных турбеллярий вторичным упрощением. Не говоря о том, что для этого нельзя привести никаких экологических обоснований, оно не подтверждается ни сравнительно-анатомическими, ни эмбриологическими фактами. Напротив, последние, как мы могли неоднократно убедиться, заставляют рассматривать всю паренхиму Acoela как фагоцитобласт, гомологичный таковому общих предков Metazoa,
Что касается предполагаемой редукции целома, то это допущение не подтверждается эмбриональным развитием турбеллярий. Спиральное дробление Archoophora значительно примитивнее такового аннелид по ряду признаков (см. стр. 148—152). С известными нам фактами гораздо лучше согласуется предположение, что первичные мезобласты у примитивных форм способны давать только элементы паренхимы (энтомезенхиму) и лишь у высших Spiralia стали источником для формирования целома. В процессе эволюции первоначально однородный фагоцитобласт с исключительно внутриклеточным пищеварением сперва разделился на две области — опорную периферическую и пищеварительную центральную. Затем и та и другая эпителизировались, хотя и не одновременно. Сначала центральная часть фагоцитобласта образовала эпителиальный кишечник (энтодерму), затем опорная паренхима дифференцировалась в виде целома. Приведенные в данной работе морфологические факты и отношения вряд ли допускают какое- либо иное толкование.
Рассмотрим, однако, особенности отдельных гипотез регрессивной эволюции Plathelminthes.
Архицеломатная теория Альварадо
Филогенетические концепции Альварадо, в частности его толкование природы Plathelminthes, были опубликованы в пятидесятых годах (Alvarado, 1955, 1956, 1959) и остались малоизвестными.1
Исходя из гипотезы Гастреи, Альварадо приходит к представлению о первично целомической природе Bilateria. Вслед за Ремане он производит их от четырехлучевых Anthozoa через гипотетического Procoelomon, причем считает энтероцельный модус образования целома первичным. За исходные формы Bilateria онПринимает гипотетические организмы, сходные с личинками современных Deuterostomia. От Procoelomon через личинкообразную Doliolaria Альварадо производит Echinodermata, а через Тогпа-
Т / Bet
Scaphopoda
Pelecypoda
Polychaeta
. pogoriophora Pterohranchia
Enteropneusta
Sipimculidea

Praerbipidoglossum
Trocbozoon-
Kamptozoa
fphylactolaemata
' Qymnolaemata Graptolitha
,Phoronidea
Brachiopoda
Echivodermata

/
/
VerteBrata
Pvocbovdata
QctodracMa
TetraBranchiata j jffaflracAiar Cestoda Nematoda
. \Acanthocepbala JTrematoda \ Nmatomorha
Siphonopoda J \ у
Gastropoda TurSellaria ^^otatoria^
polyplacophora J Hirudinea j Kinorhyncha
ЫеТгаУ OUgochaeta U-Priapulidea
/Nemertinea / \ r Artbropoda
Doliolaria
Tornaria
Procoelomon
Рис. 110. Филогенетические отношения между группами Metazoa по Альварадо (Alvarado, 1955).
ria — остальных вторичноротых. От второго ствола берет начало боковая ветвь, ведущая к Vertebrata. Продуктом дальнейшей эволюции Deuterostomia Альварадо считает совокупность олигомерных сидячих форм — Bryozoa, Phoronidea, Kamptozoa и Po
gonophora. Их общие предки, по его мнению, обладали склонностью к развитию спирального дробления и имели трохофорообразную личинку, а также испытали редукцию первичного энтероцельного целома. От этих гипотетических предков четырех упомянутых групп,1которых Альварадо характеризует как филогенетическую стадиюTrochozoon, произошли три большие ветви: Mollusca, Plathelminthes и ветвь, ведущая к полимерным аннелидам и членистоногим и к конечной регрессивной линии эволюции, приводящей к Nemathelminthes. Plathelminthes также представляют собой регрессивный ряд от Nemertini к Turbellaria — затем к Trematoda и Cestoda.
Представление об этом очень оригинальном, но чрезвычайно сложном филогенетическом древе может дать его схематическое изображение (рис. 110). Из него видно, какой запутанный и, очевидно, недостаточно оправданный филогенетический путь предполагается для плоских червей — он пролегает через последовательные стадии: Anthozoa ■— Procoelomon—Tornaria—Hemichor-
data—Archicoelomata— Trochozoon—немертинообразные формы—
Turbellaria. В построениях Альварадо несомненно много материала для критики, но мы лишь отметим, что намеченная им филогенетическая линия, приводящая в конце концов к Plathelminthes, представляется нам неприемлемой.
Теория Spiralia 2 Ремане
Теория Spiralia (Spiralier-Theorie) несомненно связана с идеями Э. Ван Бенедена, В. В. Заленского, К. Н. Давыдова, Ж. Боаса, А. Нэфа и А. Ламера, которые в той или иной форме признавали родство между аннелидами и турбелляриями и первых считали предками вторых. В. В. Заленский (1907) и Ламер (Lameere, 1910—1911, 1931) производили турбеллярий от пиявок, Боас (Boas, 1917) и Нэф (Naef, 1931) — от немертин, которых они в свою очередь считали потомками аннелид.
Развивая идеи В. В. Заленского, К. Н. Давыдов определял плоских червей как формы паренхиматозные и бесполостные, «имевшие целом, но утратившие его» (Давыдов, 1914, стр. 199). Сходные мысли одновременно с К. Н. Давыдовым высказал и Гейдер (Heider, 1914), а позднее они легли в основу теории Spiralia, развиваемой рядом выдающихся зоологов нашего времени (Remane, 1950, 1954, 1958, 1959; Ulrich, 1950, 1951; Marcus, 1958;
Jflgersten, 1959; Ax, I960, 1961, 1963a; Brien, 1961, 1969), которые пытаются найти в онтогенезе и в организации ресничных червей следы их происхождения от целомических сегментированных предков. Эти последние мыслятся как формы, стоящие гораздо ниже современных аннелид и обладающие еще тремя первичными сегментами — архимерами. Предполагается, что предки турбеллярий претерпели глубокие упрощения организации.
Сущность этого учения очень обстоятельно и последовательно изложена Аксом в его статье, посвященной эволюции и происхождению турбеллярий (Ах, 1961), и мы лучше всего поступим, если последуем за ним, расчленив его аргументацию на отдельные пункты. Итак, основы теории покоятся на следующих положениях.
Turbellaria несомненно принадлежат к Spiralia. Это доказывается тем фактом, что первичное для турбеллярий «спиральное квартетное — Ы — дробление» Polycladida полностью сходно с таковым других членов этой большой группы животных.
Spiralia представляют собой совокупность родственных групп, расположенных на линии развития Protostomia выше от- хождения от нее ветви Tentaculata (см. рис. 111).
Следовательно, какие бы то ни было предположения о прямой связи турбеллярий с Protozoa или с Coelenterata излишни.
Теория Spiralia предусматривает целый ряд изменений в организации, приведших к прототипу Turbellaria. Эти изменения были следующими:
редукция целомических полостей, или, точнее, утрата бластомером Ы потенции к образованию эпителиальных мешков между эктодермой и энтодермой и разрастание за его счет диффузной паренхимы;
связанная с этим утрата кровеносной системы;
редукция задней кишки и ануса;
переход от наружного к внутреннему оплодотворению;
замена раздельнополого состояния гермафродитным.1
Г/ Для доказательства этих предположений приводятся аналогии и «модельные случаи», почерпнутые из других групп животных.
Редукция целома и его частичное превращение в паренхиматозную ткань находит себе параллели у различных Coelomata,
а именно — у Mollusca, Hirudinea, Myzostomida и Enteropneusta.
Исчезновение кровеносной системы происходит также у Аг- thropoda (Copepoda, Ostracoda, Acari) и, кроме того, у Hirudinea.
У различных животных известны многочисленные случаи утраты задней кишки и ануса (Marcus, 1934; Remane, 1951). Поэтому, утверждает Акс, можно допустить аналогичный процесс и при становлении морфологического типа Turbellaria, даже не
зависимо от того, верна ли или нет теория Ремане о превращении задней кишки целомических предков турбеллярий в бурсальный орган (бурса + ее стебелек), вошедший в состав женского полового ^аппарата.
Articulata
Mollvsca
Дейтомерная исходная форма
Nemertini
ТигвеЫагьа
Превращение зачатт целома 6 мезенхиму. Редукция кробеносной с-мы, задней кишки и ануса.
Protostomia
/Червеобразный организй с целомом, и спиральным *кбартетным-^й-дроВлениеи^
Procoelomctta про-мезо- и метацели
Ступень Goelenterata
Рис. 111. Филогенетиче&кие отношения между основными группами Coelomata по теории Spiralia Ремане. (По: Ах, 1961).
Рассмотрим эти положения теории Spiralia.
Первое утверждение о принадлежности Plathelminthes к Spiralia ни у кого не вызывает сомнений. Однако из него никак не вытекает, что филогенетические построения рассматриваемой теории верны. Мы тоже признаем единство Spiralia, но считаем, что филогенетическое развитие спирального дробления шл
о
не от аннелидйого Варианта tc поликладйдному и адельыому, а в обратном направлении.
Второе утверждение представляет собой очень краткое определение положения Spiralia на генеалогическом древе Metazoa, предложенном Ремане, и носит догматический характер, так как не содержит никаких доказательств.
В третьем пункте предлагается признать отсутствие филогенетических связей турбеллярий с простейшими и низшими Metazoa как логический вывод из предыдущего утверждения, уже признанного нами неубедительным.
Четвертый пункт представляет собой основное содержание теории Spiralia — объяснение эволюционных процессов становления турбеллярий. Поэтому чрезвычайно важно убедиться, насколько предполагаемые изменения организации подтверждаются тем сравнительно-морфологическим и эмбриологическим материалом, который дает изучение турбеллярий. Если ресничные черви действительно произошли от сегментированных предков, обладавших кровеносной системой и задней кишкой с анусом, а также энтеро- цельным целомом (как это вытекает из существа теории Корал- лулы), то в организации низших турбеллярий должны остаться хоть какие-нибудь следы прежнего плана строения. В действительности же такие свидетельства отсутствуют.' Проктодеальная теория Ремане (Remane, 1951), рассматривающая семенную бурсу турбеллярий как остаток задней кишки, опровергается рядом авторов, в том числе самими сторонниками теории Spiralia (Bock, 1927; Meixner, 1938, 1943; Marcus, 1946; Karling, 1958; Ax, 1961). Фундаментальное сходство спирального дробления у турбеллярий и аннелид, как мы уже убедились, не таково, чтобы служить доказательством предполагаемой редукции целома.
Что касается ссылок на аналогии, то, разумеется, одного этого явно недостаточно для обоснования столь серьезных выводов, тем более что многие приведенные примеры вряд ли удовлетворяют всем требованиям, предъявляемым к хорошим «модельным случаям».
Так, редукция целома и частичное замещение его соединительной тканью у Mollusca, Hirudinea, Myzostomida и Enteropneusta не совсем то, что предполагает теория Spiralia, так как не связана с общей деградацией организма. У моллюсков, которые, вероятно, произошли от олигомерных аннелид с ларвальными сегментами (дейтомерами, по терминологии Ремане), с самого начала не было сегментированного целома (ср. с Neopilina), а в дальнейшем происходило лишь сокращение целомических полостей, но не полная их редукция. У предков Myzostomida, которые тоже относятся к олигомерным формам с ларвальными сегментами, также трудно предположить наличие сегментированного целома, а у самих мизостомид целомические полости хорошо развиты, хотя и сильно расчленены (Федотов, 1915, 1937). У Hirudinea и Enteropneusta, несмотря на частичную редукцию целомических пространств, полностью сохраняется план строения сегментированных животных. Кроме того, у рассматриваемых групп никакой связи между сокращением целомических полостей и состоянием кровеносной системы нет. Последняя не редуцируется, за исключением Hirudinea, у которых, как известно, происходит физиологическая субституция исчезающей кровеносной системы своеобразно развивающимся целомом; это совсем не то, что предполагает теория Spiralia.
Аналогию с исчезновением кровеносных сосудов у Copepoda, Ostracoda и Acari также нельзя признать удачной, так как во всех этих случаях речь идет о редукции незамкнутой кровеносной системы членистоногих, которая в известном смысле несравнима с замкнутым сосудистым аппаратом Coelomata, полностью отделенным от целома. Кровеносная система Arthropoda, как известно, отличается неполным развитием сосудов и включением полости тела — миксоцеля — в систему кровообращения. В этой ситуации при уменьшении размеров тела, естественно, легко происходит дальнейшее сокращение кровеносных сосудов вплоть до полного их исчезновения, но общая система кровообращения, поскольку в ней участвует полость тела, сохраняется. У аннелид уменьшение размеров тела никогда не приводит к заметной редукции кровеносных сосудов. Далее, у членистоногих незамк- нутость кровеносной системы коррелятивно связана с исчезновением целома (при сохранении объемистых пространств первичной полости тела), а редукция последнего — с развитием наружного хитинового скелета. Ничего подобного предполагать у предков турбеллярий не приходится.
Разумеется, теория Spiralia органически' связана с гипотезой Ремане, выводящей сегментированных Procoelomata от гипотетического кораллового полипа Corallula. Однако мы не разделяем этих идей по ряду соображений и прежде всего потому, что не видим возможностей к значительной эволюции у таких узкоспециализированных сидячих животных, как Anthozoa.
Очень убедительны замечания по поводу теории Spiralia, недавно высказанные Рейзингером (Reisinger, 1970, 1972). Он отметил, что в пользу многократной редукции целома в разных группах Scolecida нет решительно никаких свидетельств, несмотря на многочисленность и крайнее разнообразие современных Spiralia Acoelomata. Эволюция внутри Spiralia совершалась от Acoelomata к Coelomata, а не наоборот, что особенно ярко подтверждают судьбы нервного ортогона у Spiralia. Невозможно представить себе происхождение радиально симметричного по существу ортогона низших турбеллярий с его несколькими парами продольных стволов от брюшной нервной лестницы.
Против теории Spiralia высказались также Сальвини-Плавен и Кодреану. Первый автор после тщательного изучения примитивных моллюсков (из классов Solenogastres и Caudofoveata) разработал новый вариант гипотезы происхождения этих животных от турбеллярий, которых рассматривает как видоизмененных потомков первобытных фагоцителлообразных многоклеточных (Sal- vini-Plawen, 1968, 1969).
Кодреану (Codreanu, 1970) в интересной статье, посвященной критической оценке современных филогенетических концепций эволюции беспозвоночных животных, пришел к сходным выводам. Теория Spiralia, пишет Кодреану, тщетно пытается заполнить громадный разрыв, существующий между Coelenterata и Annelida; для этого она использует энтероцельную гипотезу происхождения целома, явно неприменимую к Protostomia. Теория Spiralia вынуждена предполагать исчезновение архимеров (про-, мезо- и метацелей), не имея возможности доказать, что они когда-либо существовали.
Заканчивая наш анализ теории Spiralia, мы должны еще раз подчеркнуть, что приведенный в данной работе сравнительноморфологический материал свидетельствует против этой концепции.
Ктенофорная теория Ланга
Из гипотез второй группы самая старая — ктенофорная теория. Идея о происхождении турбеллярий, именно Polycladida, от гребневиков впервые была высказана А. О. Ковалевским в 1880 г., в статье, в которой он описал первого представителя ползающих гребневиков — Coeloplana metschnikowi. Внешний вид и некоторые особенности организации Coeloplana невольно наводили на эту мысль. Несколькими годами позже Ланг детально разработал эту гипотезу в известной монографии по поликлади- дам Неаполитанского залива (Lang, 1884).
Более или менее радиальное расположение ветвей кишечника и нервных стволов у поликладид Ланг считал следами радиальной симметрии, унаследованной от Ctenophora. Ланг сравнивал с Coeloplana и с типичными гребневиками поликладид, вроде Stylochus и Planocera, обладающих широким лепешковидным телом, почти в центре которого располагается вентральный рот, а почти над ним — мозг (рис. 112).
Ланг приравнивал брюшную поверхность поликладид оральной стороне гребневиков, спинйую сторону поликладид — або- ралъной стороне гребневиков, дорсо-вентральную ось турбеллярий — главной оси тела ктенофор. Правая и левая стороны червей, по Лангу, соответствуют щупальцевым антимерам, а сагиттальная плоскость — глоточной плоскости симметрии гребневиков. Гомологичными органами у тех и других оказались мозг турбеллярий и аборальный орган ктенофор, глотки, разветвления кишки и гастроваскулярных каналов, теменные щупальца поликладид и щупальца гребневиков.
Перестройка организации гребневиков, приведшая к появлению первых турбеллярий — поликладид, была обусловлена переходом от плавающего к ползающему образу жизни и сводилась к выработке адаптаций к ползанию на ротовой стороне тела, которая стала затем брюшной поверхностью. Животные стали двигаться вперед одной из глоточных антимер, которая превратилась в передний конец тела. Мозг и связанные с ним щупальца сместились к переднему краю тела, ротовое отверстие тоже сдвинулось несколько вперед (Cotylea) или назад (Acotylea).
Со стороны эмбриологии Ланг также находил подкрепление своим взглядам. И гребневики и поликладиды имеют детерминированное дробление с образованием микро- и макромеров, в обоих
Рис.
112. Схема, иллюстрирующая происхождение
турбеллярий от гребневиков. (По: Lang,
1884).
Л
— гребневик, В
— гипотетическая промежуточная форма,
В
— поликладида. а—а
— щупальцевая плоскость симметрии, аЬ
— аборальный орган, Ь—ь
— глоточная плоскость сим-' метрии, с
— мозг, gc
—
гастральные каналы, ветви кишечника,
ph
—
глотка, t
—
щупальца.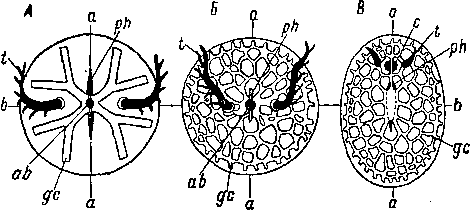
случаях глотка образуется как большое стомодеальное впячи- вание. Часть поликладид обладает мюллеровской личинкой, восемь выростов которой, окаймленных ресничным шнуром, Ланг сравнивал с меридиональными рядами гребных пластинок у Ctenophora.
Эволюцию внутри класса Turbellaria Ланг представлял себе следующим образом. Из плана строения Polycladida Acotylea он выводил трикладид путем уменьшения числа парных ветвей кишечника и редукции главной кишки — «желудка». От Tricladida посредством дальнейшего упрощения кишки он производил мелких Alloeocoela и Rhabdocoela. Что касается Acoela, то их Ланг рассматривал как потомков неотенических личинок поликладид (Lang, 1884). Кроме того, по Лангу, от турбеллярий возникли кольчатые черви (Lang, 1881). Некоторые Tricladida (вроде морской Gunda segmentata=Procerodes lobata) приобрели довольно совершенную внутреннюю метамерию вследствие вторичного упорядочения в расположении, множественных органов — гонад, ветвей кишечника, нервных комиссур. На почве этой метамерии возникла истинная сегментация аннелид. Целом последних образовался разрастанием полостей метамерно расположенных гонад (семенников). Таким образом, Ланг связывал вопрос о происхождении аннелид от турбеллярий с гоноцельной теорией целома. Эта гипотеза Ланга носит умозрительный, спекулятивный характер, логически не связана с его ктенофорной теорией происхождения поликладид и значительно уступает ей по убедительности и обоснованности.
Ктенофорная теория казалась очень хорошо обоснованной и пользовалась большим успехом до самого недавнего времени (Mortensen, 1912; Wilhelmi, 1913; Krumbach, 1925). Еще сравнительно недавно ее защищал В. А. Догель (1954), который, однако, внес в нее некоторые изменения и дополнения. Основную аргументацию Ланга Догель считал совершенно правильной. Упоминая о гомологических сходствах между Polycladida и Cteno- ph.ora, он писал: «Если мы будем объяснять все эти черты сходства одной только конвергенцией, то нам придется вообще отказаться от всяких филогенетических выводов, настолько большинство ныне принимаемых по отношению к разным животным гипотез окажется более шатким, чем защищаемая в данном случае Лангом связь между Polyclada и Platyctenea» (Догель, 1954, стр. 32). Однако происхождение других отрядов турбеллярий Догель не считал возможным связывать с ползающими гребневиками.
Таким образом, В. А. Догель принимал «двойственное происхождение турбеллярий, но вместе с тем от одной и той нее исходной группы, а именно от гребневиков». К Polycladida он применял ланговскую ктенофорную теорию, a Acoela выводил «от личиночных стадий ктенофорообразных предков, которые стали размножаться неотенически, не достигая состояния взрослой ктено- форы» (там же, стр. 32). По мнению В. А. Догеля, только такое объяснение происхождения турбеллярий делает понятным «отсутствие явственных переходов между поликладами и прочими отрядами турбеллярий».
Ктенофорная гипотеза, основанная на рассмотрении одних Polycladida, не учитывает организации и развития представителей других отрядов турбеллярий. Главное же возражение против нее заключается в том, что она неправильно приравнивает дорсо-вентральную ось турбеллярий аборально-оральной оси тела гребневиков (см.: Беклемишев, 1964). Из развития мюллеровской личинки совершенно ясно, что ее задний конец соответствует оральному полюсу, а передний конец — аборальному полюсу гребневика. Главная ось тела турбеллярий не перпендикулярна главной оси тела гребневиков, цо совпадает с ней. Иначе говоря, Turbellaria — такие же протаксонные животные, как и все Coelenterata. Основное проморфологическое различие между ними заключается в том, что у турбеллярий развилась билатеральная симметрия, а ротовое отверстие сместилось более или менее вперед по брюшной стороне тела.
По справедливому замечанию Бресслау и Рейзингера (Bresslau
u. Reisinger, 1928—1933), ктенофорная теория характеризуется преимущественно описательно-морфологической трактовкой вопроса и пренебрегает эмбриологическими фактами, с которыми она не согласуется. Но даже со сравнительно-анатомической точки зрения она далеко не безупречна. Ланг делает подчас весьма сомнительные сопоставления; например, гомологизирует щупальца ктенофор с чувствительными щупальцевидными придатками поликладид, или сравнивает орган равновесия гребневиков, в области которого нет нервного центра, с мозгом турбеллярий, или, наконец, гомологизирует восемь рядов гребных пластинок Ctenophora восьми лопастям мюллеровской личинки.
Хорошо известные эмбриологические факты особенно противоречат ктенофорной теории. Принципиальные различия между Ctenophora и Polycladida начинаются с самого начала их развития. Дробление их совершенно различно. Личинки Polycladida никогда не обнаруживают никаких ктенофорных признаков и вообще очень далеки от цидиппоидных личинок гребневиков. В онтогенезе поликладид имеется ацелоидная стадия с плотным зачатком кишки (центральной паренхимой), из которого гораздо позднее путем рассасывания части энтодермальных бластомеров формируется эпителиальный кишечник с полостью. У Acoela, напротив, в развитии нет ничего напоминающего инвагинацион- ную гаструлу1(Bresslau, 1909; Bresslau u. Reisinger, 1928— 1933). He оправдались и попытки Ланга привлечь ползающих гребневиков — Platyctenida — в качестве модельных форм, иллюстрирующих переход от Ctenophora к Polycladida. Весь онтогенез ползающих гребневиков резко отличен от такового турбеллярий. В начале жизненного цикла они проходят стадию типичной цидиппоидной личинки, как все Ctenophora. Все же особенности взрослых платиктенид, «сближающие» их с турбелляриями, проявляются в конце жизненного цикла; они носят поверхностный характер и представляют собой не что иное, как результат специализации — приспособления к ползанию по дну. Сходство Platyctenida и Polycladida объясняется конвергенцией. «Тот, кто хочет принять ктенофорную теорию на основе внешних сходств между высокоспециализированными ползающими ктенофорами и поликладами, должен закрыть глаза перед этими совершенно ясными данными сравнительной эмбриологии», — замечают Бресслау и Рейзингер (Bresslau u. Reisinger, 1928—1933, стр. 50).
Наконец, основные пути эволюции главнейших групп турбеллярий, выясняющиеся ныне в результате сравнительно-морфологического анализа, совершенно не согласуются с представлениями о первичной примитивности Polycladida.
Трохофорная теория Гатчека
Трохофорную теорию Гатчека (Hatschek, 1888) иногда рассматривают как вариант ктенофорной гипотезы на том основании, что она тоже выводит турбеллярий от гребневиков (см.: Hyman, 1951). Общим предком всех трохофорных животных (Annelida, Mollusca, Tentaculata) Гатчек считал гипотетического Trochozoon, организация которого рекапитулируется личинкой трохофорой. Trochozoon выводится от гребневиков; о строении его ближайшего предка Protrochozoon дает представление личинка протрохула, свойственная ресничным червям и немертинам (мюллеровская личинка, пилидий). Turbellaria, по Гатчеку, представляют собой боковую эволюционную ветвь, развившуюся от форм, близких к Protrochozoon.
Трохофорная теория в значительной мере основана на про- морфологических сходствах между Ctenophora и личинкой трохофорой. Для обоснования этих сходств особенно много было сделано Вольтереком (Woltereck, 1904, 1905) и В. Н. Беклемишевым (1944, 1964). В симметрии гребневиков и трохофоры были обнаружены интересные общие особенности, а именно элементы двулучевой симметрии в комбинации с четырех лучевой и дихотомической восьмилучевой. Однако, с нашей точки зрения, этот тип симметрии в обоих случаях возник независимо и есть следствие симметрии дробящегося яйца при детерминированности развития
и, следовательно, не может иметь филогенетического значения.
Гастреадная теория Карлинга
Карлинга (Karling, 1940) можно считать основателем гастреад- ной теории происхождения турбеллярий. Самыми примитивными среди ресничных червей он считает формы, наиболее приближающиеся к Acoela й особенно к Catenulida и обладающие эпителиальным кишечником (рис. 113). Карлинг рассматривает отсутствие кишки у Acoela как явление вторичное на том основании, что ацелоидный кишечник имеется и у многих Neorhabdocoela и у Р1а- giostomidae (Prolecithophora), где он якобы явно вторично упрощен. В данном случае первичное состояние у «высших форм» принимается ошибочно за вторичное, а затем этот вывод переносится и на низшие формы, т. е. допускается та же методическая ошибка, которую мы отмечали у Акса.
Н. А. Ливанов (1945, 1955) и Д. М. Федотов (1966) по существу солидаризируются с Карлингом. Ливанов, подвергнув резкой критике концепцию первичного отсутствия кишечника у Acoela (его аргументы рассмотрены на стр. 79—82), утверждает, что начало турбелляриям дала одна из ветвей гастрееобразных Coelenterata, развившая мезенхиму с «мезенхимогенными мышцами». По Ливанову, это были общие предки гребневиков и турбеллярий. Приведенные в нашей работе сравнительно-морфологические данные и их анализ говорят против этих представлений.

А — первичная форма с кишкой и простым вентральным ртом или pharynx simplex, без поповых отверстий, с дорсальным общим половым зачатком; Б — следующая стадия с вентральным мужским половым отверстием; В — образование женского выводного полового протока и ductus communis; Г — образование pharynx plicatus, общего орально-генитального отверстия; из общего полового зачатка развиваются слермии, яйца и желточные клетки; Д — развитие компактных, одетых tunica propria гонад: семенников и гермовителляриев, образование гонодуктов; Е — обособление ротового и полового отверстий, образование самостоятельных гонад без оболочка и гонодуктов, развитие pharynx variabilis из ph. plicatus.Планулоидная, или планулоидно-ацельная, теория Граффа
Особую примитивность Acoela, наличие у них аморфного фагоцитобласта и близость их к гипотетической Фагоцителле отметил еще И. И. Мечников (1877а, 1886). О происхождении ресничных червей от планулоидного организма писали многие авторы (Steinmann u. Bresslau, 1913; Bresslau и. Reisinger, 1928— 1933; Беклемишев, 1937, 1952; Hyman, 1951). Обычно самую идею происхождения от лланулообразного предка приписывают Граффу, ссылаясь на его работы (Graff, 1882, 1891, 1904— 1908, 1904), но в действительности у Граффа нигде нет высказываний в этом роде. В большой монографии 1891 г. «Die Organisation der Turbellaria Acoela» на стр. 50—55 Графф сравнивает бескишечных турбеллярий с Trichoplax adhaerens, найденным Ф. Шульце (Schulze, 1883) в морском аквариуме в Триесте. Тело этого организма состоит из наружного эктодермального жгутикового эпителия и паренхиматозной внутренней массы. Вместе с другой близкой формой — Treptoplax reptans, описанной Монти- челли (Monticelli, 1896), Trichoplaxв конце XIX в. был причислен к Mesozoa как особый класс — Mesenchymia (Delage et Herouard, 1899). Позднее было высказано предположение, что оба организма представляют собой модифицированные планулы гидроидной медузы Eleutheria (Krumbach, 1907). Это мнение было принято без критики и проникло на страницы многих учебников и руководств (Шимкевич, 1923; Hyman, 1940; Grasse, 1961; Иванов, 1968). Недавно, однако, Грелль обнаружил у Trichoplax половое размножение и описал первые стадии эмбрионального развития, показав, таким образом, что этот организм не личинка, а вполне закончившее развитие многоклеточное животное (Grell, 1971, 1972). Ныне строение и даже ультраструктура Trichoplax adhaerens хорошо известны (Grell u. Benwitz, 1971; Иванов, 1973).
Изучив Trichoplax на срезах, Графф пришел к выводу, что это животное очень близко к предкам Acoela. В работах этого автора нет непосредственных сравнений Acoela или Trichoplax с планулами книдарий. Тем не менее никто не сделал так много для обоснования теории происхождения турбеллярий от плануло- образных предков, как Графф, который своими фундаментальными исследованиями показал глубокую примитивность Acoela. Неудивительно поэтому, что эта концепция связывается с именем Граффа. Об организации предков ресничных червей Графф писал: «Поскольку все турбеллярии проходят стадию стерро- гаструлы с паренхимой, содержащей еще необособленные элементы энто- и мезодермы, нет сомнения в том, что предкам Turbellaria должна быть приписана ацелия, как примитивный признак, и что современные Acoela ближе всего стоят к этим предкам» (Graff, 1904, стр. 206). Второй существенной особенностью анцестральных форм Графф считал радиальную симметрию нерв-
193
А. В. Иванов и Ю. В. Мамкаев
ной системы. «Наличие от трех до шести пар одинаковых и радиально расположенных вокруг главной оси тела продольных нервных стволов . . . есть первичный признак, который частично или полностью утрачивается с приспособлением к ползающему образу жизни» (там же, стр. 206). И наконец, третья важная черта предковых форм турбеллярий, по Граффу, — первичное отсутствие выделительных органов.
Сравнивая Acoela с личинками Cnidaria — планулами, мы вслед за В. Н. Беклемишевым (1944) и другими авторами должны приравнивать передний конец турбеллярий аборальному, а задний конец — оральному полюсу ппанулы. Главная ось тела пла- нулы и паренхимулы гомологична продольной оси турбеллярий. Первичное положение рта у Turbellaria, очевидно, должно быть задним. Оно сохранилось лишь у немногих, очень примитивных форм. Граффу были известны только Urostomatidae (из Prolecithophora) с таким первично локализованным ртом. Теперь нам известны еще три вида Diopisthoporus (из Acoela).
Гомология осей турбеллярий и личинок книдарий подтверждается проморфологией эмбриональных стадий развития у тех и других. Acoela, будучи уже настоящими Bilateria, тем не менее сохранили еще элементы радиальной симметрии, которая проявляется очень ясно в расположении вокруг главной оси тела продольных нервных стволов ортогона.
Серьезным подтверждением теории Граффа следует считать также развитие у Acoela фронтального чувствительно-железистого комплекса органов и зачаточного мозга около статоциста на переднем конце тела. Подобные образования на первично-анималь- ном полюсе — чрезвычайно характерная особенность примитивных Metazoa и их личинок. Глубокая примитивность Acoela проявляется и в таких чертах строения, как эпителиально-мускульные клетки, отсутствие базальной мембраны эпидермиса и вообще системы пограничных перепонок, наличие неэпителизованного фагоцитобласта, эпидермальная нервная система, диффузные гонады, отсутствие у многих форм дополнительных женских половых органов, дуэтный тип спирального дробления и многие другие особенности.
Главные возражения против теории Граффа, которые делались разными авторами, сводятся к следующему.
Acoela — вторично упрощенные организмы, что прежде всего выражается в утрате у них кишечника.
Выведение турбеллярий из планулообразного предка наталкивается на серьезные методические трудности.
Признание Acoela наиболее примитивными турбелляриями нарушает представление о гомологии кишечника у всех Metazoa, установленной теорией Гастреи.
Рассмотрим внимательно эти возражения.
Многие зоологи вслед за Карлингом и Ремане отрицают примитивность Acoela, находя у них многочисленные, большей частью не связанные друг с другом вторичные упрощения строения.
Однако, по справедливому замечанию Вестблада (Westblad, 1948), для этого заключения нет никаких экологических оснований. Acoela — свободноживущие, очень подвижные животные, многие из них быстрые хищники, активно разыскивающие добычу и нападающие на нее. Между тем хорошо известно, что значительные упрощения организации обычно происходят только у животных паразитических или у малоподвижных, сидячих или полусидячих форм.
Свои возражения Ремане (Remane, 1958) начинает с заявления, что он вполне сознает всю неблагодарность задачи доказать регрессивную эволюцию у свободных, не паразитических и не сидячих животных. После такого вступления читатель, естественно, ожидает получить серьезный разбор всей проблемы, но Ремане ограничивается лишь немногими сомнительными аналогиями. Он приводит несколько примеров вторичного упрощения организации, которое, по его мнению, не находит себе никаких экологических оправданий, как в случае Acoela. Такой характер, по Ремане, носит редукция кишечного тракта у погонофор, с чем нельзя согласиться, ибо эти животные — типичные седентарные существа, обитающие в длинной защитной трубке. Перед нами как раз тот случай пассивного, полусидячего образа жизни, который имел в виду Вестблад как одну из причин редукции. Погонофоры питаются пассивно, подобно настоящим сидячим животным, воспринимая пищу через покровы и щупальца (Иванов, 1960, 1963). Это целомические метамерные животные,
находящиеся на гораздо более высоком уровне развития, чем низшие турбеллярии, и сопоставления одних с другими не очень уместны.
Далее Ремане ссылается на случаи необъяснимой редукций кишки «в десяти явно установленных линиях». К сожалению, он не называет этих примеров, почему их и невозможно обсуждать.
Наконец, Ремане приводит уже разобранные нами (стр. 186) случаи упрощения организации у мелких членистоногих — Ostracoda, Copepoda и Acari. Таким образом, заключает Ремане, для обоснования регрессивной эволюции Acoela имеются хорошо установленные модельные случаи. Мы с этим согласиться не можем.
Доводы Н. А. Ливанова, говорящие, по его мнению, в пользу вторичного упрощения Acoela, мы рассмотрели выше (стр. 79—82, 110—112). В последней своей статье Ливанов (1970) выдвинул новый аргумент. Он утверждает, что наличие у Acoela «архитектоники турбеллярного типа», при развитой у них смешанной паренхиме, свидетельствует о их происхождении от турбеллярий с эпителиальным кишечником. В основе этого утверждения лежит недоказуемое убеждение, что турбеллярный план строения мог выработаться лишь у ресничных червей с вполне развитыми тканями и органами. Однако это убеждение ошибочно.
Действительно, как показывает все, что мы знаем о строении живых существ, рабочие структуры любого организма, находящегося на любом уровне организации, будь то элементы доклеточ- ного существа, органеллы одноклеточного организма или органы и ткани метазоона, всегда располагаются вполне целесообразно по отношению к целому организму. Иными словами, у организмов всех уровней всегда вырабатывается своя определенная архитектоника, или план строения. Это вполне естественно, поскольку никакая целостная система без архитектонического расположения своих частей существовать не может. В самом деле, даже вирусы состоят из вполне определенным образом локализованных частей — в центре РНК или ДНК, снаружи белковая оболочка, нередко, кроме того, имеется хвостик, или стебелек, а иногда и другие прикрепительные образования. Любое моноэнергидное простейшее, как и любая клетка метазоона, обладает определенным набором органелл — ядром, клеточной мембраной, эндоплазмати- ческими канальцами, митохондриями, рибосомами и т. д., причем все эти рабочие структуры расположены целесообразно и вполне определенным образом по отношению ко всей клетке, т. е. в клетке имеется своя характерная архитектоника, которая стала особенно явной после цитологических электронномикроскопических исследований. То же можно сказать и о более сложных организмах. Сомателлы инфузорий, например, тоже обладают своей архитектоникой, которая, кстати сказать, конвергентно сходна с таковой некоторых Metazoa — обстоятельство, позволившее А. А. Заварзину (1932) сделать известное сопоставление организации паразитической инфузории и коловратки. Различные группы Metazoa в процессе эволюции приобрели свои характерные планы строения. Acoela, находящиеся еще, подобно губкам, на нетканевом уровне организации Metazoa, тоже выработали свою архитектонику, которую от них унаследовали другие турбеллярии. Наконец, в колониях Metazoa — кормусах, при достаточно высоком их развитии (у сифонофор, мшанок, сальп), также вырабатывается своя собственная архитектоника (Беклемишев, 1950, 1964).
Весьма поучительный пример развития архитектонических особенностей представляют Porifera. Между тем губки, подобно Acoela, отличаются развитием типичной смешанной паренхимы, прекрасно охарактеризованной В. Н. Беклемишевым (1964). Если архитектоника организма, обладающего смешанной паренхимой, может считаться только наследием тканевых предков, как утверждает Ливанов, то позволительно спросить: от каких высокоорганизованных анцестральных форм следует производить губок?
Гораздо естественнее трактовать Acoela как турбеллярий, находящихся на более низком, чем остальные ресничные черви, нетканевом уровне организации, Турбеллярный план строения впервые стал складываться у непосредственных предков Acoela, когда они перешли от свободноплавающего к ползающему образу жизни. Такие проморфологические особенности, как билатеральная симметрия, вырабатывались с одинаковым успехом у организмов, стоящих на самых различных уровнях эволюции, разумеется, при соответствующих образе жизни и способе передвижения, а именно у различных Protozoa, у личинок-планул некоторых Anthozoa и при формировании первых Bilateria. Впрочем, у Acoela, как отмечалось уже выше, мы находим еще значительные следы первобытной радиальной симметрии в расположении элементов нервного ортогона. Нередко и эндональный мозг Acoela имеет ясно выраженную радиальную симметрию, например у Diopisthoporus longitubus (Westblad, 1940). Нельзя не вспомнить здесь также и положение рта на заднем конце тела у этой бескишечной турбеллярии. В состоянии женской половой зрелости, пока нет еще мужского совокупительного органа, эта форма не имеет никаких признаков билатеральной симметрии (Беклемишев, 1964).
Заднее положение рта нетипично для других турбеллярий, в виде исключения оно встречается только у некоторых, очень примитивных Prolecithophora (из семейства Urostomatidae). Это отклонение от плана строения типичной турбеллярии носит явно первичный характер, как и радиальное расположение нервных стволов. Далее, половая система Acoela явно не укладывается в план строения типичных турбеллярий. У Acoela нет еще обособленных гонад, а энтолецитальные яйца и спермии в сущности рассеяны в паренхиме. Гонодукты отсутствуют также. Это крайне примитивное состояние сравнимо лишь с той ступенью развития полового аппарата, которую мы видим у губок. К сожалению, Н. А. Ливанов не рассматривает половую систему Acoela, между тем ее сравнение с таковой остальных турбеллярий очень поучительно и дает много ценного материала для суждения о природе бескишечных ресничных червей.
При обсуждении природы Acoela и их архитектоники Н. А. Ливанов (1970, стр. 529) смешивает две разные проблемы: вопрос о происхождении архитектонических правильностей в организации бескишечных турбеллярий и вопрос об эволюционном формировании у них тканей. Н. А. Ливанов ошибочно приписывает В. Н. Беклемишеву мнение о том, что архитектоника Acoela выводится из смешанной паренхимы путем сегрегации клеток. В действительности ничего подобного нигде у В. Н. Беклемишева мы не находим. Вопрос о том, как возникла архитектоника турбеллярий, для него ясен, поскольку он видит в Acoela самых примитивных Bilateria. Естественно, что приспособление их предков к ползающему образу жизни и привело к выработке у них билатеральной симметрии и турбеллярного плана строения (Беклемишев, 1964, I, стр. 105—116).
Большая заслуга В. Н. Беклемишева в том, что он первый подметил эволюционные пути формирования первичных тканей у низ-
ших Metazoa. Эти процессы можно наблюдать как раз у форм, находящихся еще на нетканевом уровне организации, т. е. у губок и бескишечных ресничных червей. У последних «уже проявляется некоторая наклонность к сегрегации, т. е. обособлению и собиранию воедино клеток одной системы, зачаточному тканеобразова- нию». «В понятии ткани, — пишет далее Беклемишев (1964, II, стр. 9), — не мыслится никакой определенной формы» и, добавим мы, никакой определенной архитектонической локализации.
Совершенно ясно, что Беклемишев в цитируемом месте своего руководства делает попытку лишь объяснить, как формируются ткани и органы. Вопрос же о том, почему ткани и органы располагаются в теле животного так, а не иначе, составляет другую самостоятельную проблему.
Итак, при формировании плана строения организма архитектоническому упорядочению подвергаются рабочие структуры, или элементы эргатома, любого организационного уровня. С одинаковым успехом это могут быть и клеточные органеллы простей-' шего, и отдельные клетки смешанной паренхимы Porifera и Acoela, и комплексы одинаковых клеток — ткани и органы высших животных. Именно поэтому и вопреки мнению Н. А. Ливанова между тканеобразованием и становлением архитектонических правильностей нет никакой обязательной связи.
Что касается ссылок на методические трудности, якобы связанные с планулоидной теорией, то, по мнению Ремане, предположение о «происхождении турбеллярий от планулы Cnidaria, которая отличается от Гастреи отсутствием рта и полости кишки в энтодерме, находит мало приверженцев по той причине, что прыжок от планулы к Turbellaria слишком велик для методического обоснования» (Remane, 1958, стр. 189). Это утверждение можно понять только в одном смысле. Очевидно, прыжком здесь назван переход от организма, лишенного еще рта и кишечника, к организму с эпителиальной кишкой и ротовым отверстием. Не считая ацелию бескишечных турбеллярий первичной, Ремане сопоставляет планулу не с Acoela, а с турбелляриями, имеющими уже кишку. При таком сравнении для него проще и естественнее сопоставлять турбеллярий с геккелевской Гастреей, чем с паренхимулой или планулой Cnidaria. Однако, даже став на эту точку зрения, было бы натяжкой характеризовать переход к эпителизации внутренней клеточной массы как методически необоснованный прыжок. Во всяком случае «прыжок» здесь гораздо меньше, чем при переходе от организации сидячего полипа к подвижному метамерному представителю Bilateria (по теории Корал- лулы).
Наконец, противники теории Граффа подчеркивают, что Acoela нельзя признать самой примитивной группой турбеллярий, потому что в этом случае пришлось бы допустить независимую эпителизацию кишечника у Coelenterata (Cnidaria, Ctenophora) и у низших Bilateria, По этим же соображениям, наиболее вероят- йой гипотезой происхождения Metazoa должна быть геккелевская теория Гастреи.
Однако вряд ли можно считать эпителизацию первично аморфного кишечника (фагоцитобласта) настолько сложным процессом, чтобы исключить возможность ее многократного повторения в филогенезе. Мы видели выше (стр. 71—74), с какой легкостью совершается этот процесс у громадного большинства турбеллярий, у которых он носит характер обратимого перехода от одного гисто-физиоло- гического состояния кишечника к другому.
Отказ от признания независимого появления эпителизованной кишки в разных филогенетических ветвях не мешает, однако, противникам первичности ацелии у бескишечных турбеллярий принимать конвергентное развитие куда более сложных структур. Так, Акс признает многократное формирование сложных типов глотки у Neorhabdocoela и Prolecithophora, а также у Polycladida и Proporoplana, не считая, что это противоречит принципиальным методическим установкам. По его собственным словам, имело место и конвергентное развитие некоторых органов женского полового аппарата. «При обработке семейства Otoplanidae, — пишет он, — я получил свидетельства тому, что функционально одинаковые и эквивалентные бурсальные органы ни в коем случае не могут обладать единым происхождением и что генито-интести- нальные сообщения могут возникать вторично» (Ах, 1961, стр. 35).
Концепцию происхождения Acoela от планулообразных предков особенно поддерживали и развивали Бресслау и Рейзингер (Bresslau u. Reisinger, 1928—1933) и В. Н. Беклемишев (1937, 1952). Однако планулоидная теория существует в нескольких, серьезно различающихся вариантах. Сначала Гайман внесла в нее некоторые собственные толкования, затем Беклемишев предложил новую интерпретацию, существенно меняющую ее филогенетический смысл.
Планулоидная теория в трактовке Гайман
Защищая представление о первичной примитивности организации Acoela, Гайман писала во введении второго тома своего из* вестного руководства по зоологии беспозвоночных: «Мы принимаем теорию происхождения Bilateria от планулоидного предка через ацелоидную форму. Главными изменениями были — замена радиальной симметрии билатеральной и смещение нервного центра вперед. Так как промежуточные формы неизвестны, то путь, по которому происходили эти изменения, может быть выведен ив эмбриологии низших Turbellaria. Так как рот у Acoela занимает бластопоральную сторону, то его можно принять за постоянную точку. Далее, как постулировал Ланг, очевидно, планулоидная форма должна была уплощаться по орально-аборальной оси, и одновременно должна была произойти миграция вперед чувствительного центра, как это происходит при развитии поликладид в процессе дифференциального роста. . . В результате получйлСй примитивный плоский червь с билатеральной симметрией, вентральным ртом и локализованными впереди органами чувств. Дальнейшая стадия, представленная высшими Turbellaria, была достигнута путем образования полости в центральной части внутренней массы, т. е. формирования архентерона и большей дифференциации передней области — головы» (Hyman, 1951, стр. 9).
п о
Рис.
114. Схема, иллюстрирующая планулоидную
теорию происхождения Bilateria.
(По:
Hyman,
1951).
А — личинка планула, без рта, с апикальным нервным центром; Б — образование рта и укорочение орально-аборальной оси тела; В — тело вытягивается в сагиттальной плоскости, нервный центр сдвигается вперед; Г — ацелоидная стадия, билатерально симметричный, ползающий червь с передним нервным центром; Д — одна из поздних стадий эмбрионального развития поликладид. ес — эктодерма, еп — энтодерма, in — первичная кишка, тез — мезодерма, п — нервный центр, ns — нервные стволы, о — ротовое отверстие, ph — стомодеум.
Итак, Гайман использует проморфологические положения ктенофорной теории Ланга. Она считает оральную сторону книда- рий соответствующей брюшной поверхности турбеллярий и полагает, что дорсо-вентральная ось последних, сперва отвечавшая аборально-оральной оси Coelenterata, затем наклонилась (или изогнулась) верхним концом в сторону вновь образующегося переднего конца тела, вызвав при этом смещение мозга вперед (рис. 114).
Однако проморфологический анализ Acoela не позволяет сделать этих выводов; он показывает ясно, что ни о каком наклоне или искривлении дорсо-вентральной оси тела у предков турбелля- рий не может быть речи. В самом деле, оральный полюс тела сохраняет у Acoela свое прежнее место — на заднем конце тела и только ротовое отверстие смещается вперед на брюшную поверхность. Это особенно подчеркивается тем, что продольные нервные стволы у Acoela тянутся меридионально, до заднего конца тела, сохраняя везде радиально симметричное расположение вокруг передне-задней оси тела. Кроме того, кожно-мускульный мешок у бескишечных турбеллярий представлен правильной решеткой перекрещивающихся продольных и поперечных волокон; эта решетка, не изгибаясь и не деформируясь, достигает заднего конца тела. Как и нервные стволы, она характеризуется вполне правильным радиальным расположением вокруг главной оси тела. Следовательно, положение брюшного рта большинства современных Acoela явно вторично, тогда как нервный аппарат и кожная мускулатура сохранили еще свою первоначальную локализацию (Беклемишев, 1964).
Планулоидная теория в трактовке В. Н. Беклемишева
В. Н. Беклемишев в последние годы своей жизни изменил свои представления о происхождении Acoela (Беклемишев, 19636).
Беклемишев подчеркивал, что особенно важные заключения о гомологии органов и о генетическом родстве животных дают изучение и сравнение онтогенеза и всего жизненного цикла. Сравнивая жизненные циклы Cnidaria и Turbellaria, Беклемишев пришел к заключению, что у последних он укорочен. Из яйца Hydrozoa развивается бластула, которая превращается после образования зародышевых листков в паренхимулу, а потом в пла- нулу. Если же развитие протекает под защитой гонофора или споросака, то стадии бластулы и паренхимулы эмбрионизируются. Подобная же эмбрионизация бластулы характерна для всех турбеллярий, у которых яйца развиваются под защитой оболочек. Фаза, гомологичная паренхимуле или плануле, есть конечная стадия в развитии турбеллярий, и дальнейший их онтогенез сводится в основном к гистологической и органологической дифференциации, к выработке билатеральной симметрии и к смещению рта на брюшную сторону.
Таким образом, по В. Н. Беклемишеву, примитивная турбел- лярия не только соответствует паренхимуле Cnidaria, но и утратила конечные фазы жизненного цикла, характерные для Hydrozoa. Отсюда Беклемишев делает вывод, что Turbellaria произошли от личинок Cnidaria, ставших неотеническими.
Эти соображения В. Н. Беклемишева несомненно связаны с его взглядами на происхождение Metazoa. Следуя А. А. Захваткину (1949), он представляет себе общих предков Metazoa в виде сидячих колониальных организмов, в жизненном цикле которых была лишь одна подвижная расселительная стадия бластулообразной личинки, близкой по организации к плануле современных Сиг-
А — колония Craspedomonadina типа Sphaero'eca; Б — колония Craspedomonadina типа Proterospongia; В —■ ранняя Фагоцителла; Г — образование рта на заднем конце тела; Д — первичная ацелоидная турбеллярия (переход к ползающему образу жизни, усиление дифференциации клеток, выработка билатеральной симметрии); Е — Turbellaria Асоela, смещение рта на брюшную сторону; Ж — примитивная губка (переход к сидячему образу жизни, замена локомоторной функции киноцйтов гидрокинетической, погружение кино- цитов внутрь тела — филогенетическая инверсия зародышевых листков); 3 — первичное свободноплавающее кишечнополостное типа Гастреи, родоначальник Cnidaria и Cteno- phora; И — фагоцителлообразный организм типа Trichcrplax.
daria. С этой точки зрения ацелоидные первобытные турбеллярий могли произойти только от свободной личинки неких книдарие- образных организмов.
Однако основания теории Синзооспоры А. А. Захваткина чрезвычайно шатки, как и всяких иных попыток производить Metazoa от сидячих предков (Lameere, 1929; Larsson, 1963). Несостоятельность этих концепций, по нашему мнению, доказана (см.: Иванова- Казас и Иванов, 1967; Иванов, 1968).
Наиболее вероятный вариант планулоидной теории
Итак, очень мало вероятия, чтобы первичные ацелоидные турбеллярии были непосредственными потомками неотенических личинок каких-то книдариеобразных организмов. Вряд ли родоначальниками турбеллярий могли быть и общие предки Cnidaria и Ctenophora, т. е. первичные свободноплавающие Coelenterata. Об организации последних мы можем только догадываться, и тем не менее мы можем утверждать, что это были протаксонные, вероятно гастрееобразные организмы, плававшие при помощи жгутов, с полностью эпителизованным целентерическим кишечником (см.: Иванов, 1968, стр. 263), т. е. формы, уже утратившие аморфный фагоцитобласт (рис. 115, 3). Между тем один из существенных выводов нашей работы заключается в том, что вслед за Граффом мы должны признать ацелию существенной первичной особенностью предков турбеллярий. Поэтому мы полагаем, что эволюционная ветвь турбеллярий и остальных Bilateria отделилась от общего ствола первобытных Metazoa очень рано, еще до обособления линии первичных Coelenterata. Иначе говоря, предком ацелоидных турбеллярий могла быть только Фагоци- телла — мечниковский гипотетический предок всех Metazoa.
Мы полагаем, следовательно, что несомненное принципиальное сходство Acoela с паренхимулой и с планулой Cnidaria существует лишь потому, что эти личинки рекапитулируют организацию общего предка книдарий и турбеллярий, т. е. Фагоцителлу, точно так же, как рекапитулируют ее и паренхимулы губок (Иванов, 19716).
По-видимому, между Фагоцителлой и первичными ацелоид- ными турбелляриями существовала промежуточная филогенетическая стадия. Последнюю, следуя идеям творца теории Фагоци- теллы, И. И. Мечникова, можно представить себе как паренхимулообразное существо, плававшее с помощью жгутов и обладавшее уже ротовым отверстием на заднем конце тела (рис. 115, Г). Это были уже поляризованные организмы с передним, анимальным, и задним, вегетативным (оральным), полюсами, с жгутиковым кинобластом и аморфным фагоцитобластом, богатым амебообразными пищеварительными клетками.
От них, вероятно, и произошли турбеллярии. Это заключение неизбежно вытекает из всего сказанного выше.(Беклемишев В. Н.) Beklemischev W. 1914. Uber einige acoele Turbellarien des Kaspischen Meeres. Zool. Anz., 45 : 3—7.
Беклемишев В. H. 1915. О паразитных турбелляриях Мурманского моря. 1. Тр. Петрогр. общ. естествоисп., 43, 4 : 103—172.
Беклемишев В. Н. 1916. О паразитных турбелляриях Мурманского моря. 2. Тр. Петрогр. общ. естествоисп., 45, 4 : 3—56.
Беклемишев В. Н. 1925. Морфологическая проблема животных структур. (К критике некоторых из основных понятий гистологии). Изв. Биол. н.-иссл. инст. Пермск. унив., 3, прилож. 1 : 1—74.
Беклемишев В. Н. 1927а. К фауне турбеллярий Одесского залива и впадающих в него ключей. Изв. Биол. н.-иссл. инст. Пермск. унив.,
5 : 177—207.
(Беклемишев В. Н.) Beklemischev W. 19276. Uber die Turbellarienfauna des Aralsees. Zool. Jahrb., Abt. Syst.,„54: 87—138.
(Беклемишев В. H.) Beklemischev W. 1929. Uber den Bau der Driisenstachel der Anaperiden (Turbellaria, Acoela). Zool. Anz., 80 : 232—235.
Беклемишев В. H. 1937. Класс ресничных червей (Turbellaria). В кн.: Руководство по зоологии. I. М.—Л. : 386—457.
Беклемишев В. Н. 1944. Основы сравнительной анатомии беспозвоночных. Изд. «Советская наука», М. : 1—492.
Беклемишев В. Н. 1950. К проблеме индивидуальности в биологии. Колонии у двустороннесимметричных животных. Усп. совр. биол., 29, 1 : 91—120.
Беклемишев В. Н. 1951. О видах рода Macrostomum (Turbellaria, Rhabdocoela) СССР. Бюлл. Моск. общ. исп. природы, отд. биол., 4 : 31—40.
Беклемишев В. Н. 1952. Основы сравнительной анатомии беспозвоночных. Изд. 2-е. М. : 1—698.
(Беклемишев В. Н.) Beklemischev W. 1960. Grundlagen der vergleichenden Anatomie der Wirbellosen. II. Berlin. VEB Deutsch. Verlag. Wiss. : 1—403.
Беклемишев В. H. 1963a. Ресничные черви (Turbellaria) Каспийского моря. III. Род Oligochoerus n. gen. (Acoela, Convolutidae). Бюлл. Моск. общ. исп. природы, отд. биол., 68, 1 : 29—40.
(Беклемишев В. Н.) Beklemischev V. 19636. On the relationship of the turbellaria to the other groups of the animal kingdom. In: The lower Metazoa. Ed. E. C. Dougherty. Univ. Calif. Press : 234— 244.
Беклемишев В. H. 1964. Основы сравнительной анатомии беспозвоночных. Изд. 3-е. Изд. «Наука», М. I (Проморфология) : 1—432; II (Органология) : 1—446
.Богомолов С. Й. 1949. К вопросу о типе дробления Rhabdocoela. Уч. зап. ЛГУ, сер. биол., 113, 20 : 128—142.
Богомолов С. И. 1960. История развития Convoluta в связи с морфологией ресничных червей. Тр. Общ. естествоисп., 63; Уч. зап. Каз. унив., 120, 6 : 155—208.
Б о г у т а К. К. 1970. Новая турбеллярия Anaperus biaculeatus sp. n. (Turbellaria, Acoela) из сублиторали Белого моря. Зоол. журн., 49,
: 198—209.
Б о гут а К. К. 1971. Ранний онтогенез Anaperus biaculeatus (Turbellaria, Acoela). Зоол. журн., 51, 3 : 332—340.
Быховский Б. Е. 1937. Онтогенез и филогенетические взаимоотношения плоских паразитических червей. Изв. АН СССР, сер. биол., 4 : 1353—1383.
Быховский Б. Е. 1957. Моногенетические сосальщики, их система и филогения. Изд. АН СССР, М.—JI. : 1—509.
Воронцов Н. Н. 1963. Неравномерность-темпов преобразования органов и принцип компенсации функций. Зоол. журн., 42, 9 : 1289—1305.
Гиляров М. С. 1958. Эволюция характера оплодотворения наземных членистоногих. Зоол. журн., 37, 5 : 707—735.
Г и н е ц и некая Т. А. 1968. Трематоды, их жизненные циклы, биология и эволюция. Изд. «Наука», Л. : 1—411.
Давыдов К. Н. 1914. Курс эмбриологии беспозвоночных. Пб.—Киев : 1-502.
Д е п е р е Ш. 1915. Превращения животного мира. Пг. : 1—267.
Догель В. А. 1936. Олигомеризация гомологичных органов как один
из процессов эволюции животных организмов. Арх. анат., гист. и эмбриол., 15, 3 : 101—114.
Догель В. А. 1938. Сравнительная анатомия беспозвоночных. I. Л. :
1-599.
Догель В. А. 1940. Сравнительная анатомия беспозвоночных. II. Л. : 1-495.
Догель В. А. 1947. Явление полимеризации и олигомеризации гомологичных органов в животном царстве и их эволюционное значение. Изв. АН СССР, сер. биол., 4 : 471—486.
Догель В. А. 1952. Процесс олигомеризации и полимеризации числа
органов во время эволюции животных. Ч. I. Уч. зап. ЛГУ, 141, сер. биол., 28 : 3—44.
Догель В. А. 1954. Олигомеризация гомологичных органов как один из главных путей эволюции животных. Изд. ЛГУ : 1—368.
Догель В. А. 1959. Зоология беспозвоночных. Изд. 5-е (первое посмертное). Изд. «Советская наука», М. : 1—511.
Д о н д у а А. К. 1952. О развитии Rathkea octopunctata. Уч. зап. ЛГУ,
145, сер. биол. : 166—171.
Евдонин Л. А. 1969. Новый представитель интерстициальных калипто- ринхий (Turbellaria, Neorhabdocoela, Kalyptorhynchia) острова Куна- шир. Вестн. ЛГУ, биол., 3 : 7—14.
Ж и нк и н Л. Н. 1951. Особенности дробления яйцеклеток у низших беспозвоночных. Природа, 2 : 70—73.
Заварзин А. А. 1932. Курс общей гистологии. Гос. изд., Л. : 1—376.
(Заленский В. В.) S а 1 е n s.k у W. 1907. Morphologische Studien an Wiirmer. Zweiter Teil. III. Uber die Metamorphose des Polygordius. Teil IV. Zur Theorie des Mesoderms. Mem. Acad. Sci. SPb., Ser. VIII, 19, 14 : 183—264, 265-349.
Захваткин A. A. 1949. Сравнительная эмбриология низших беспозвоночных. (Источники и пути формирования индивидуального развития многоклеточных). Изд. «Советская наука», М. : 1—395.
Иванов А. В. 1952а. Бескишечные турбеллярии (Acoela) южного побережья Сахалина. Тр. Зоол. инст. АН СССР, 12 : 40—132.
Иванов А. В. 19526. Строение Udoneila caiigorum Johnston, 1835 и положение Udonellidae в системе плоских червей. Паразитол. сб.' ЗИН АН СССР, 14 : 112—163.
Иванов А. В. 1960. Погонофоры. Фауна СССР, новая сер., 75, М.—Л. : 1—271.
(Иванов А. В.) Ivanov А. 1963. Pogonophora. Academic Press. London : 1—479.
Иванов А. В. 1967. О монофилетической природе Metazoa. Зоол. журн., 46, 10 : 1446—1455.
Иванов А. В. 1968. Происхождение многоклеточных животных. (Филогенетические очерки). Изд. «Наука», JI. : 1—287.
Иванов А. В. 1971а. К вопросу о примитивности бескишечных ресничных червей Acoela. Зоол. журн., 50, 5 : 621—632.
Иванов А. В. 19716. Эмбриональное развитие губок (Porifera) и положение их в системе животного мира. Журн. общ. биол., 32, 5 : 557—572.
Иванов А. В. 1973. Trichoplax adhaerens — фагоцителлообразное животное. Зоол. журн., 52, 8:1117—1131.
Иванова-Казас О. М. 1959а. К вопросу о происхождении и эволюции спирального дробления. Вестн. ЛГУ, сер. биол., 9, 2 : 56—67.
(Иванова-Казас О. М.) Ivanova-Kasas О. М. 19596. Entstehung und Evolution der Spiralfurchung. Naturwiss. Beitr., 12 :
: 1267—1279.
Иванова-Казас О. М. и А. В. Иванов. 1967. О происхождении Metazoa и их онтогенеза. (Критическая оценка гипотезы синзоо- споры А. А. Захваткина). Тр. Зоол. инст. АН СССР, 44 : 5—25.
Кана ев И. И. 1963. Очерки из истории сравнительной анатомиидо Дарвина. Развитие проблемы морфологического типа в зоологии. Изд. АН СССР, М.—Л. : 1—299.
К а н а е в И. И. 1966. Очерки из истории проблемы морфологического типа от Дарвина до наших дней. Изд. «Наука», М.—Л. : 1—210.
Ковалевский А. О. 1865. История развития Amphioxus lanceolatus, или Branchiostoma lubricum. Дисс. для получ. степени магистра зоологии. СПб. : 1—47.
(Ковалевский А. О.) Kowalevsky А. 1880. Ueber Coeloplana metschnikowi. Zool. Anz., 3.
Корсакова Г. Ф. 1949. Ранние стадии дробления у Tiaropsis. ДАН СССР, 65, 3 : 413—415.
Кулинич Л. Я. 1970. Новый вид рода Plicastoma (Turbellaria, Prolecithophora) из Японского моря. Зоол. журн., 49, 3 : 339—348.
Ливанов Н. А. 1937. Тип немертин (Nemertini). В кн.: Руководство по зоологии. 1. М.—Л. : 656—705.
Ливанов Н. А. 1941. Основные этапы эволюции нервной системы. Уч. зап. Каз. гос. унив., 101, 1 : 79—142.
Ливанов Н. А. 1945. Пути эволюции животного мира. (Анализ организации типов). Уч. зап. Каз. гос. унив., 105, 3, Зоология, 9 : 5—199.
Ливанов Н. А. 1955. Пути эволюции животного мира. (Анализ организации главнейших типов многоклеточных животных). Изд. «Советская наука», М. : 1—400.
Ливанов Н. А. 1970. О происхождении Metazoa. Зоол. журн., 49,
: 517—532.
Логачев Е. Д. 1957. Материалы к вопросу о взаимоотношениях тканей внутренней среды и пограничных тканей у плоских червей в онто- и филогенезе. Кемерово : 1—22.
Логачев Е. Д. 1970. О филогенетическом положении цестод. Класс или тип? Тез. докл. чтений памяти акад. Е. Н. Павловского в АН КазССР "5 марта 1970 г., Алма-Ата—Кемерово : 3—12.
Лукин Е. И. 1964. Различия в скорости эволюции разных систем органов и приспособлений к размножению и развитию животных. Зоол. журн., 43, 8 : 1105—1120
.Мамкаев Ю. В. 1961. Новый представитель гнатостомулид — Gnatho- stomula murmanica sp. n. ДАН СССР, 141, 6 : 1501—1504.
(Мамкаев Ю. В.) Mamkaev Yu. V. 1965. Etude morphologique d’Actinoposthia beklemischevi n. sp. (Turbellaria, Acoela). CahiersBjol. Mar., 6 : 23—50.
Мамкаев Ю. В. 1967. Очерки по морфологии бескишечных турбелля
рий. Тр. Зоол. инст. АН СССР, 44 : 26—108.
Мамкаев Ю. В. 1968. Сравнение морфологических различий в низших
и высших группах филогенетического ствола. Журн. общ. биол., 29,
: 48—56.
Мамкаев Ю. В. 1971.0 конволютах (Turbellaria, Acoela) южного Приморья. Фауна и флора зал. Посьет Японского моря. Иссл. фауны морей, VIII (XVI) : 31-54.
Мамкаев Ю. В. иЕ. А. Котиков а. 1972. О морфологических
особенностях нервного аппарата бескишечных турбеллярий. Зоол. журн., 51, 4 : 477—489.
Мамкаев Ю. В. и JI. Н. Серавин. 1963. Питание бескишечной
турбеллярии Convoluia convoluta (Abildgaard). Зоол. журн., 42,
: 197-205.
Мейер Э. 1897. Исследование над развитием кольчатых червей. Тр. Общ. естествоисп. Каз. унив., 31, 5 : 1—356.
Меницкий Ю. JI. 1963. Строение и систематическое положение паразитирующей в рыбах турбеллярии Ichtyophaga subcutanea Syromjatni- kowa, 1949. Паразитол. сб. ЗИН АН СССР, 21 : 245-258.
Мечников И. И. 1877а. О пищеварительных органах пресноводных турбеллярий. Зап. Новоросс. общ. естествоисп., 5, 1 : 1—12. (Мечников И. И. 1955. Акад. собр. соч. Т. 1. Медгиз, М. : 252—259).
Мечников И. И. 18776. Исследования о развитии планарий. Зап. Новоросс. общ. естествоисп., 5, 1 :1—16. (Мечников И. И.
Акад. собр. соч. Т. 3. Медгиз, М. : 51—60).
(Мечников И. И.) Metschnikoff Е. 1883. Untersuchungen iiber die mesodermalen Phagocyten einiger Wirbeltiere. Biol. Zentralblatt,
18 : 560—565. (Мечников И. И. 1954. Акад. собр. соч. Т. 5. Медгиз, М. : 16—20).
(Мечников И. И.) Metschnikoff Е. 1886. Embryologische Stu- dien an Medusen. Ein Beitrag zur Genealogie der Primitiv-Organe. Wien: 1—159. (Мечников И. И. 1950. Избр. биол. произв. Изд. АН СССР, М.—Л. : 271-471).
(Насонов Н. В.) Nassonov N. V. 1932. Zur Morphologie der Turbellaria Rhabdocoelida des Japanischen Meeres. I, II. Тр. Лаб. эксп. зоол. морфол. животных, 2, 6 : 6—115,
(Переяславцева С.) Perejaslawzewa S. 1892. Monographie des Turbellaries de la mer Noire. Зап. Новоросс. общ. естествоисп., 17, 3 : 1-303.
Робертсон Д. 1964. Мембрана живой клетки. Структура и функция клетки. М. : 159—172.
Северцов А. Н. 1916. Исследование об эволюции низших позвоночных.
Русск. арх. анат., гист. эмбр., 1, 1 : 1—114.
Северцов А. Н. 1934. Морфологические закономерности эволюционного процесса. Соц. реконстр. и наука, 3 : 21—37.
Северцов А. Н. 1967. Главные направления эволюционного процесса. (Морфобиологическая теория эволюции). Изд. 3-е. Изд. МГУ : 1—202.
Сыромятникова И. П. 1949. Новая, паразитирующая в рыбах турбеллярия Ichtyophaga subcutanea nov. gen., nov. sp. ДАН СССР, 68, 4 : 805-808.
Ульянин В. Н. 1870. Ресничные черви (Turbellaria) Севастопольской бухты. Тр. II съезда русск. естествоисп. в Москве в 1869 г., отд. зоол.,
: 1—96.
Федотов Д. М. 1915. Protomyzostomum polynephris Fedotov и его отношение к Gorgonocephalus eucnemis М. et. Тг. Пг. : 1—240.
Федотов Д. М. 1937. Специализация и деградация в строении мезосто- мид под влиянием образа жизни. В сб.: Акад. Н. В. Насонову к 80-ле- тию со дня рождения и 60-летию научн. деят. Изд. АН СССР, М. : 52— 81.
Федотов Д. М. 1966. Эволюция и филогения беспозвоночных животных. Изд. «Наука», М. : 1—404.
Фридман Г. М. 1933. Анатомическое строение Baicalarctia gulo Fr. и положение ее в системе Turbellaria. Тр. Байкальской лимнол. ст.,
: 179—256.
Хлопин Н. Г. 1946. Общебиологические и экспериментальные основы гистологии. Изд. АН СССР, JI. : 1—492.
Шимкевич В. М. 1923. Биологические основы зоологии. Т. 1. Изд. 5-е. Гос. изд., М.—JI. : 1—384.
Шмидт Г. А. 1951. Эмбриология животных. I. Общая эмбриология. Изд. «Советская наука», М. : 1—355.
Шмальгаузен И. И. 1938. Организм как целое в индивидуальном и историческом развитии. М.—Л. : 1—144.
Шмальгаузен И. И. 1939. Пути и закономерности эволюционного процесса. М.—Л. : 1—231.
Шмальгаузен И. И. 1969. Проблемы дарвинизма. Изд. 2-е, перераб. и доп. Изд. «Наука», Л. : 1—493.
Шульц Р. С. и Е. В. Гвоздев. 1970. Основы общей гельминтологии, I. Изд. «Наука», М. : 1—492.
Alvarado R. 1955. Sobre el origin у ia evolution de los metazoos celo- mados. Bol. R. Soc. Espan. Hist. Nat. (Biol.), 52 : 147—154.
Alvarado R. 1956. Palingenesis у cenogenesis en las formas larvarias de
invertebrados. An. Acoc. Espan. Progr. Ci., 21 : 413—436.
Alvarado R. 1959. La teoria de la evolucian a los cien anos de la obra de
Darwin. Rev. Univ. Madrid, 8 : 29—31, 9—14.
An der Lan H. 1936. Ergebnisse einer von E. Reisinger und 0. Steinbock mit Hilfe des Rask-0rsted Fonds durchgefiihrten zoologischen Reise in Gronland 1926. VII. Acoela I. Vidensk. Medd. fra Dansk. naturh. Foren, 99 : 289—330.
Antonius A. 1968. NeueConvolutidaeundeineBearbeitung des Verwandt- schaftskreises Convoluta (Turbellaria, Acoela). (Faunistische Studien am Roten Meer im Winter 1961/62, Teil IV). Zool. Jahrb., Syst., 95 : 297—394.
A p e 1 t G. 1969. Fortpflanzungsbiologie, Entwicklungszyklen und verglei- chende Friihentwicklung acoelen Turbellarien. Mar. Biology, 4, 4 : 267— 325.
A x P. 1952. Ciliopharingiella intermedia nov. gen., nov. spec., Reprasentant einer neuen Turbellarien-Familie des marinen Mesopsammon. Zool. Jahrb., Syst., 81 : 286—312;
A x P. 1956. Die Gnathostomulidae, eine ratselhafte Wurmgruppe aus dem Meeressand. Akad. Wiss. Lit., Mainz, Abh. math.-naturwiss. Kl., Jahrg.
8 : 1—32.
A x P. 1957. Vervielfachung des mannlichen Kopulationsapparates bei Turbellarien. Verh. Deutsch... Zool. Ges. Graz : 227—249.
A x P. 1959. Zur Systematik, Okologie und Tiergeographie der Turbellarien- fauna in den ponto-kaspischen Brackwassermeeren. Zool. Jahrb., Syst., 87, 1/2 : 43-184.
Ax P. 1960. Die Entdeckung neuer Organisationstypen im Tierreich. Die
neue Brehm-Biicherei, 258 : 1—116.
A x P. 1961. Verwandtschaftsbeziehungen und Phylogenie der Turbellarien. Ergebn. Biol., 24 : 1—68.
A x P. 1963a. Relationship and phylogeny of the Turbellaria. In: The lower
Metazoa. Ed. E. C. Dougherty. Univ. Calif. Press ; 191—224.
A x P. 1963b. Das Hautgeisselepithel der Gnathostomulida. Verh. Deutsch.
Zool. Ges. Miinchen (1963). Leipzig : 452—461.
А х P. 1966а. Das chordoide Gewebe als histologisches Lebensformmerkmal der Sandliickenfauna des Meeres. Naturwiss. Rundschau, 19 : 282—289.
Ax P. 1966b. Eine neue Tierklasse aus dem Litoral des Meeres — Gnathosto- mulida. Umschau in Wiss. Techn., 1 : 17—23.
A x P. und G. A p e 1 t. 1971. Archaphanostoma agile (Turbellaria). Embryo- nalentwicklung. Encyclop. cinematogr., Film E 1138/1967. Gottingen : : 3-7.
A x P. und R. A x. 1969. Eine Chorda intestinalis bei Turbellarien. Akad.
Wiss. Lit., Mainz, Abh. math.-naturwiss. Kl., 5 : 135—158.
A x P. und J. Dorjes. 1966. Oligochoerus limnophilus nov. spec., ein
kaspisches Faunenelement als erster Siisswasservertreter der Turbellarie Acoela in Fliissen Mitteleuropas. Int. Rev. Ges. Hydrobiol., 51, 1 : 15— 44.
Baer J. G. 1929. Contribution a l’etude des Temnocephales Dactyloce-
phala madagascariensis (Vayssiere, 1892). Bull. Biol. France Belgique, 63 : 539—561.
Baer J. G. 1931. Etude monographique du groupe des Temnocephales.
Bull. Biol. France Belgique, 65, 1 : 1—57.
Baer J. G. et L. E u s e t. 1961. Classe des Monogenes. Traite de Zool.,
publ. P. Grasse, 4, 1 : 243—325.
Ball S. C. 1916. The development of Paravortex gemellipara (Graffilla
gemellipara Linton). Journ. Morphol., 27 : 453—558.
Bargmann W. und А. К n о о p. 1959. Elektronenmikroskopische Untersuchungen an Plazentarzotten des Menschen. (Bemerkungen zum Syncytiumproblem). Zeitschr. Zellforsch., 50 : 472—493.
Beauchamp P. 1949. Biospeleologica, 69. Turbellaries 3e serie. Arch. Zool. exp. gen., 86 : 50—64.
Beauchamp P. 1951. Turbellariec Polyclades du Maroc et Mauritanie. Nouvelles stations des Planaires rheophiles dans 1’Atlas. Bull. Soc. Sci. Nat. Maroc., 29 : 239—252.
Beauchamp P. 1961. Classe des Turbellaries. Traite de Zool., publ. P. Grasse, 4, 1 : 35-212.
Berg S. E. 1941. Die Entwicklung und Kolonienbildung bei Funiculina quadrangularis. Zool. Bidr. Uppsala, 22 : 1—1000.
Bergendal D. 1892—1893. Polyposthia similis n. g., n. sp. en Acotyl Polyclad med mSnga hanglia Parningsapparater. Kgl. Fysiogr. Salsk. Handl., 4 : 1—29.
Boaden P. J. S. 1963. The interstitial Turbellaria Kalyptorhynchia from some North Wales beaches. Proc. Zool. Soc. London, 141, 1 : 173—205.
Boas J. E. V. 1917. Zur Auffassung der Verwandschaftsverhaltnisse der Tiere. I. Kopenhagen : 1—62.
Bock S. 1913. Studien iiber Polycladen. Zool. Bidr. Uppsala, 2 : 31—344.
Bock S. 1923. Eine neue marine Turbellariengattung aus Japan (Hofstenia). Uppsala Univ. Arsskrift, Math.-naturvetensk., 1 : 3—55,
Bock S. 1927. Apidioplana, eine Polycladengattung mit muskulosen Driisen- organen. Goteb. Vetensk. Handl., 4, 30, 1 : 1—116.
Boettger C. R. 1957. Stammesgeschichte und Verbreitung der Turbella- riengruppe Temnocephala. Abh. Braunschw. wiss. Ges., 9 : 26—35.
Bohmig L. 1886. Untersuchungen iiber rhabdocole Turbellarien. I. Das Genus Craffilla v. Jhering. Zeitschr. wiss. Zool., 43 : 290—328.
Bolimig L. 1895. Die Turbellaria Acoela der Plankton-Expedition. Ergeb. Plankton-Expedition Humboldt-Stiftung, 2 : 1—48.
Bohmig L. 1905. Turbellarien. Result. Voyage S. Y. Belgica en 1897— 1899. Rapp. Sci. Zool : 1—22.
В о у d e n A. A. 1953. Comparative evolution with special reference to primitive mechanisms. Evolution, 7 : 21—30.
Boyer В. C. 1971. Regulative development in a spiralian Embryo as shown by cell detetion experiments on the Acoel, Childia. Journ. Exp. Zool., 176, 1 : 97—105.
А. В. Иванов ц Ю. В. Мамкаев
209
Branderburg J. 1966. Die Reusenformen der Cyrtocyten. Zool. Beitrage,
3 : 345—417.
Brandtner P. 1934. Ergebnisse einer von E. Reisinger and 0. Steinbock mit Hilfe des Rask-0rsted Fonds durchgefiihrten zoologischen Reise in Gronland 1926. 5. Plagiostomidae. Videnskab. Medd., 97 : 87—150.
Bresslau E. 1904. Beitrage zur Entwicklungsgeschichte der Turbella- rien. I. Die Entwicklung der Rhabdocoelen und Alloeocoelen. Zeitschr. wiss. Zool., 67 : 213—332.
Bresslau E. 1906. Eine neue Art der marinen Turbellariengattung Poly- cystis (Macrorhynchus) aus dem Siisswasser. Zool. Anz., 30, 13—14 : 415—422.
Bresslau E. 1909. Entwicklung der Acoelen. Verh. Deutsch. Zool.Ges., 19 : 314—324.
Bresslau E. 1928—1933. Turbellaria. Handb. Zool. Hrsg. W. Kiikenthal und Th. Krumbach. 2. Lief. 1, 9, 16 : 52—320.
Bresslau E. und E. Reisinger. 1928—1933. Plathelminthes. All- gemeine Einleitung zur Naturgeschichte der Plathelminthes. Handb. Zool. Hrsg. W. Kiikenthal und Th. Krumbach. 2. Lief. 1 : 34—51.
Bresslau E. und H. Voss. 1913. Das Nervensystem von Mesostoma. Zool. Anz., 43 : 260—263.
Brien P. 1961. Elements de Zoologie. Notions d’anatomie comparee. II. Les Coelomates Hyponeuriens. Liege : 1—715.
В r i e n P. 1969. Principes de Phylogenese animale appliques a l’interpreta- tion des Metazoaires diploblastiques et a leur evolution en Metazoaires triploblastiques. Mem. Acad. roy. Belgique. Cl. Sci., 38, 7 : 3—110.
Bullock Т.Н. and G. A. H о r r i d g e. 1965. Structure and function in the nervous systems of Invertebrates. I, II. San Francisco—London : 1—1719.
Burger O. 1897—1907. Nemertini. Bronn’s Klass. Ordn. Tierreiches, IV,
Suppl., Leipzig : 1—542.
Codreanu R. 1970. Grands problemes controverses de revolution phylo- genetique des Metazoaires. Annee Biol., Paris, 9, 11/12 : 671—709.
С о n с 1 i n E. 1932. The embryology of Amphioxus. Journ. Morph., 54 : 69—151.
Cope E. D. 1887. The origin of the fittest. Essays on evolution. London— N. Y. : 1—467,.
D e 1 a g e Y. 1886. Etudes histologiques sur les Planaires Rhabdocoeles Acoe- les. Arch. Zool. exp. gen., Ser. 11, 4 : 109—160.
Delage Y. etE. Herouard. 1899. Traite de zoologie concrete, 2. Paris : 1—244.
Dorey A. E. 1965. The organization and replacement of the epidermis in Acoelous Turbellarians. Quart. Journ. Micr. Sci., 106 : 147—172.
Dorjes J. 1966. Paratomella unichaeta nov. gen., nov. spec., Vertreter einer neuen Familie der Turbellaria Acoela mit asexueller Fortpflanzung durch Paratomie. Veroff. Inst. Meeresforsch. in Bremerhaven, Sonderbd.
: 187—200.
Dorjes J. 1968. Die Acoela (Turbellaria) der Deutschen Nordseekuste
und ein neues System der Ordnung. Zeitschr. zool. Syst. Evolutions-
forsch., 6 : 56—452.
Du Bois-Reymond Marcus E. 1957. On Turbellaria. An. Acad. Brasil. Ci., 29, 1 : 153—191.
Ehrenberg Chr. G. 1831. Animalia evertebrata exclusis insectis recensuit Dr. C. G. Ehrenberg. Series prima cum tubularum decade prima. Bero- lini. In: Hemprich et Ehrenberg. Symbole Physicae. Folia A—D.
Fernando W. 1934. The embryology of Caridinicola indica. Proc. Zool. Soc., London : 827—850.
F о r e 1 F. A. et G. Du P 1 e s s i s. 1874. Esquisse generale de la faune pro-
fonde du Lac Leman. Bull. Soc. Vaund. Sc. Nat., 13 : 46—57.Fuhrmann О. 1930—1931. Cestoidea. Handb. Zool. Hrsg. W. Kiikenthal und Th. Krumbach. 2, 2 : 141—416.
Fulinski B. 1914. Die Entwicklungsgeschichte von Dendrocoelum lacteum Oerst. 1. Die erste Entwicklungsphase von Ei bis zur Embryonalpharynx- bildung. Bull. Acad. Sci., Crac., Cl. math.-nat., Ser. В : 147—190.
Fulinski B. 1916. Die Keimblatterbildung bei Dendrocoelum lacteum Oerst. Zool. Anz., 47 : 380—400.
Fulinski B. 1938. Zur Embryonalpharynxfrage der Trikladen. Zool. Polon., 2 : 185—207.
Gardiner E. 1895. Early development of Polychoerus caudatus Mark. Journ. Morph., II : 155—176.
G e 1 e i J. 1930. «Echte» freie Nerveneinigungen. (Bemerkungen zu den Recep- toren der Turbellarien). Zeitschr. Morphol. Okol. Tiere, 18 : 786—798.
Gemmill J. and P. T. L e i p e r. 1907. Turbellaria of the Scottish National Antarctic Expedition. Transact, roy. Soc. Edinburgh, 45: 819—827.
G i e s a S. 1966. Die Embryonalentwicklung von Monocelis fusca Oersted (Turbellaria, Proseriata). Zeitschr. Morphol". Okol. Tiere, 57 : 137—230.
G i e s a S. und P. A x. 1964. Die Gastrulation der Proseriata als ein ursprun- glicher Entwicklungsmodus der Turbellaria Neoophora. Verh. Deutsch. Zool. Ges. Kiel. (1964). Leipzig : 109—222.
Gontcharoff M. 1957. Etude des rhabdites de la trompe de Lineus ruber en microscopie electronique. C. r. Acad. Sci., Paris, 244 : 1539—1541.
Graebner I. 1969. Vergleichende elektronenmikroskopische Untersuchung der Spermienmorphologie und Spermiogenese einiger Gnatostomula Arten : Gnatostomula paradoxa (Ax, 1958), G. axi (Kirsteuer, 1964), G. jenneri (Riedl, 1960). Microskopie, 24 : 131—160.
Graff L. 1874. Zur Kenntnis der Turbellarien. Zeitschr. wiss. Zool., 24 : 123—160.
Graff L. 1882. Monographie der Turbellarien. I. Rhabdocoelida. Leipzig : 1—441.
Graff L. 1889. Monographie der Turbellarien. II. Triclada Terricola. Leipzig : 1—574.
Graff L. 1890. Enantia spinifera, der Reprasentant einer neuen Polycladen- Familie. Mitth. naturwiss. Ver. Steiermark. Jahrg. 1889, Graz : 1—16.
Graff L. 1891. Die Organisation der Turbellaria Acoela. Leipzig : 1—90.
Graff L. 1904. Marine Turbellarien Orotavas und der Kiisten Europas. 1. Einleitung und Acoela. Zeitschr. wiss. Zool., 78, 2 : 190—244.
Graff L. 1904—1908. Acoela und Rhabdocoela. Bronn’s Klass. Ordn. Thier- reichs, 4, lc : 1753—2599.
Graff L. 1911a. Vorlaufiger Bericht iiber seine mit Unterstiitzung der kai- serlichen Akademie ausgefiihrten Studien iiber die nordamerikanischen Tubellarien. 1. Acoela. Anz. math.-naturwiss. Kl. Kais. Akad. Wiss., Wien, 7 : 111—113.
Graff L. 1911b. Acoela, Rhabdocoela und Alloeocoela des Ostens der Ve- reinigten Staaten von Amerika. Mit Nachtragen zu «Marinen Turbellarien Orotavas und der Kiisten Europas». Zeitschr. wiss. Zool., 99, 1 : 1—108.
Graff L. 1912—1917. Tricladida. Bronn’s Klass. u. Ordn. Tierreichs.,
IG, Turbellaria 2 : 2601—3369.
Graff L. 1913. Turbellaria. II. Rhabdocoelida. Das Tierreich, Lief. 35 : 1—184.
Grasse P. P. 1961. Embranchement des Mesozoaires Generalites. Traite de Zool., publ. P. Grasse, 4, 1 : 1—694.
Greenberg M. J. 1959. Ancestors, embryos and symmetry. Syst. Zool., 8, 4 : 212—221.
Grell K. G. 1971. Embryonalentwicklung bei Trichoplax adhaerens F. E. Schulze. Naturwiss., 58, 11 : 570.
Grell K. G. 1972. Eibildung und Furchung von Trichoplax adhaerens F. E. Schulze (Placozoa). Zeitschr. Morphol. Tiere, 73 : 297—314.
G г е 11 К. G. und G. Benwitz. 1971. Die Ultras truktur von Trichopiax adhaerens F. E. Schulze. Cytobiologie, 4, 2 : 216—240.
Giinther K. 1962. Systematik und Stammesgeschichte. Fortschr. Zool., 14 : 269—547.
Hadzi J. 1944. Turbelarijska teorija Knidarijev. Razpr. Mat. Prir. Acad., Ljubljana, 3, 1 : 1—239.
Hadzi J. 1958. Zur Diskussion uber die Abstammung der Eumetazoen. Verh. Deutsch. Zool. Ges. Graz : 169—179.
Hadzi J. 1963. The evolution of Metazoa. Pergamon Press., London : 1—499.
H a 11 e z P. 1910. Un nouveau type d’Alloeocoele (Bothriomolus constrictus, n. g., n. s.). Arch. Zool. exp. gen., Ser. 5, 3, 48 : 611—664.
H a m a n n 0. 1885. Beitrage zur Histologie der Echinodermen. II. Die Aste- riden. Jena : 1—126.
Hand C. 1959. On the origin and phylogeny of the Coelenterates. Syst. Zool.,
4 : 191—202.
Hand C. 1963. The early worm : A Planula. In: The lower Metazoa. Ed. E. C. Dougherty. Univ. Calif. Press : 33—39.
Hanson E.D. 1958. On the origin of the Metazoa. Syst. Zool., 7, 1 : 16—47.
Hanson E. D. 1961. Convoluta sutcliffei, a new species of acoelous Turbellaria. Trans. American. Micr. Soc., 80, 4 : 423—433.
Hanson E.D. 1963. Homologies and the ciliate origin of the Eumetazoa. In: The lower Metazoa. Ed. E. C. Dougherty. Univ. Calif. Press : 7—22.
Hanstrom B. 1926. Uber den feineren Bau des Nervensystems der tricla- den Turbellarien auf Grund von Untersuchungen an Bdeloura Candida. Acta Zool., 7 : 101—105.
Hanstrom B. 1928. Vergleichende Anatomie des Nervensystems der wirbellosen Tiere, unter Beriicksichtigung seiner Funktion. Berlin : 1—628.
Harrison R. A. 1947. The embryology of Temnocephala novaezelandiae Haswell. Trans. R. Soc. New Zealand, 76 : 524—536.
Hartmann M. 1956. Die Sexualitet. Auil. 2. Stuttgart: 1—463.
Haswell W. A. 1893. A monograph of Temnocephalae. The Macleay Memorial Volum : 93—153.
Haswell W. A. 1909. The devolopment of the Temnocephalae. Part 1. Quart. Journ. Micr. Sci., 54 : 415—441.
Hatschek B. 1888—1891. Lehrbuch der Zoologie. 1. Jena : 1—144.
H e i d e r K. 1914. Phylogenie der Wirbellosen. Kultur der Gegenwart, Teil 3, Abt. 4 : 453-521.
Hendelberg J. 1967. On different types of spermatozoa in Polycladida, Turbellaria. Ark. Zool., 18 : 267—304.
Hofsten N. V. 1921. Anatomie, Histologie und systematische Stellung von Otoplana intermedia du Plessis. Zool. Bidr. Uppsala, 7 : 1—74.
Hyman L. H. 1939. Acoela and Polyclad Turbellaria from Bermuda and the Sargassum. Bull. Bingham Oceanogr. Coll. Mus. Nat. Hist. Jale Univ., 7, 1 : 1—26.
Hyman L. H. 1940. The invertebrates : Protozoa through Ctenophora. N. Y., London : 1—726.
Hyman L. H. 1942. The transition from the unicellular to the multicellular individual. In: Levels of integration in biological and social system. Biol. Symposia, 8 : 27—42.
Hyman L. H. 1951. The invertebrates : Plathyhelminthes and Rhynchocoela. The acoelomate Bilateria, II. N. Y.—Toronto—London : 1—550.
Hyman L. H. 1953. The polyclad flatworms of the Pacific Coast of North America. Bull. Amer. Mus. Hist., 100, 2 : 269—392.
Hyman L. H. 1959. The invertebrates: smaller coelomate groups. 5. N. Y.— London—Toronto : 1—783.
Hyman L. H. 1964. The phylogeny of the lower Metazoa. Quart. Rev. Biol., 39, 3 : 276-281.
Janicki С. 1907. (Jber die Embryonalentwicklung von Taenia serrata Golze. Zeitschr. wiss. Zool., 87 : 685—724.
Janicki C. 1921. Grundlinien einer «Cercomer»-Theorie zur Morphologie der Trematoden und Cestoden. Festschr. Zschokke, Basel, 30 : 1—22.
J a g e r s t e n G. 1955. On the early phylogeny of the Metazoa. The bilate- rogastraea theory. Zool. Bidr. Uppsala, 30 : 321—354.
Jagersten G. 1959. Further remarks on the early phylogeny of the Metazoa. Zool. Bidr. Uppsala, 33 : 79—108.
Jennings J. B. 1957. Studies on feeding, digestion and food storage in free-living flatworms (Platyhelminthes: Turbellaria). Biol. Bull.,
112, 1 : 63—80.
Jennings J. B. 1962. Further studies on feeding and digestion in triclad Turbellaria. Biol. Bull., 123, 3 : 571—581.
Jennings J. B. 1963. Some aspects of nutrition in Turbellaria, Trematoda and Rhynchocoela. In: The lower Metazoa. Ed. E. C. Dougherty. Univ. Calif Press : 345 353.
Jennings J. B. 1968. Nutrition and digestion. Chem. Zool., 2 : 303—326.
Jensen O. S. 1878. Turbellaria ad litora Norvegia occidentalia. Turbella- rier ved Norgesvestkyst. Bergen : 1—97.
Johnston S. I. 1913. On some Qneensland Trematodes with anatomical observations and descriptions of new species and genera. Quart. Journ. Micr. Sci., 59 : 361-400.
Karling T. G. 1940. Zur Morphologie und Systematik der Alloeocoela Cumulate uad Rhabdocoela Lecithophora (Turbellaria). Acta Zool. Fennica, 26 : 1—260.
Karling T. G. 1956a. Morphologisch-histologische Untersuchungen an den mannlichen Atrialorganen der Kalyptorhynchia (Turbellaria). Ark. Zool., 9 : 187—279.
Karling T. G. 1956b. Zur Kenntnis einiger Gnathorhynchiden nebst Beschreibung einer neuen Gattung. Ark. Zool., 9 : 343—353.
Karling T. G. 1956c. Alexlutheria acrosiphoniae n. gen., n. sp., ein be- merkenswerter marinen Vertreter der Familie Dalyelliidae (Turbellaria). Ark. Zool., Ser. 2, 19, 4 : 331—345.
Karling T. G. 1958. Zur Kenntnis der Gattung Coelogynopora Steinbock (Turbellaria Proseriata). Ark. Zool., 11 : 559—568.
Karling T. G. 1963. Some evolutionary trends in turbellarian morphology. In: The lower Metazoa. Ed. E. C. Dougherty. Univ. Calif. Press : 225—233.
Karling T. G. 1965. Haplopharynx rostratus Meixner (Turbellaria) mit der Nemertinen verglichen. Zeitschr. zool. Syst. Evolutionforsch., 3, 1—2 : 1—18.
Karling T. G. 1966a. On nematocysts and similar structures in Turbella- rians. Acta Zool. Fennica, 116 : 3—28.
Karlin g T. G. 1966b. On the defecation apparatus in the genus Archimono- celis (Turbellaria, Monocelididae). Sarsia, 24 : 37—44.
Karling T. G. 1967a. Zur Frage von dem systematischen Wert der Katego- rien Archoophora and Neoophora (Turbellaria). Commentationes Biol. Soc. Sci. Fennica, 30, 3 : 1—11.
Karling T. G. 1967b. Marine Turbellaria from the Pacific Coast of North America. 4. Coelogynoporidae and Monocelididae. Ark. Zool., 18, 6 : 493—528.
Karling T. G. 1968. On the genus Gnosonesima Reisinger (Turbellaria). Sarsia, 33 : 81—108.
Kathariner L. 1904. Ueber die Entwicklung von Gyrodactylus elegans v. Nordm. Zool. Jahrb., Suppl. 7 : 519—550.
Kelbetz S. 1962. Zum Feinbau der Rhabditen. Naturwiss., 49 : 548.
К e p n e r W. A. and J. R. С a s h. 1915. Ciliated pits of Stenostomum. Journ. Morph., 26, 2 : 235—245.
Kerkut G. A. 1960. Implication of evolution. Pergamon Press : 1—174.
Koscielski B. 1966. Cytological and cytochemical investigations on the embryonic development of Dendrocoelum lacteuin. Zool. poioil., 16 : 83-96.
Krumbach T. 1907. Trichoplax, die umgewandelte Planula einer Hydro- meduse. Zool. Anz., 31 : 450—454.
Krumbach T. 1925. Acnidaria, Ctenophora. Handb. Zool. Hrsg. W.
Kiikenthal und Th. Krumbach, 1 : 902—995.
Kiimme 1 G. 1958. Das Terminalorgan der Protonephridien. Feinstruktur und Deutung der Funktion. Zeitschr. Naturforsch., 13 : 677—679. Kiimmel G. 1962. Zwei neuen Formen von Cyrtocyten. Vergleich der bisher bekannten Cyrtocyten und Erorterung des Begriffes «Zelltyp». Zeitschr. Zellforsch., 57 : 172—201.
Kiimmel G. 1964. Morphologischer Hinweis auf einen Filtrationsvorgang in der Antennendriise von Cambarus afinis Soy. Naturwiss., 51 : 200—201. Kiimmel G. und J. Brandenburg. 1961. Die Reusengeisselzellen (Cyrtocyten). Zeitschr. Naturforsch., 16 : 692—697.
Laidlaw F. F. 1903. On the marine fauna of Zanzibar and British east Africa, from collections made by Cyril Crossland in years 1901 and 1902. Turbellaria Polycladida, I. The Acotylea. Proc. Zool. Soc. London,
: 99—113.
Lameere A. 1910—1911. Sommaire du cours d’elements de zoologie.
Ann. Soc. roy. Zool. et Malacol. Belgique, 45 : 57—152.
Lameere A. 1915—1916. Une theorie zoologique. Bull. Sci. France Belgique, 49 : 378—431.
Lameere A. 1929. Precis de zoologie. I. Bruxelles : 1—394.
Lameere A. 1931. Precis de zoologie. II. Bruxelles : 1—456.
Lang A. 1881. Der Bau von Gunda segmentata und die Verwandtschaft der Plathelminthen mit Coelenteraten und Hirudineen. Mitt. Stat. Neap., 3 : 187—251.
Lang A. 1884. Die Polycladen (Seeplanarien). Fauna u. Flora d. Golfes v. Neapel, 11 Monogr., Leipzig : 1—688.
Larsson G. 1963. Reflectons on the system of the Deuterostomia. Skrift.
Univ. Zool. Mus. Kabenhavn, 20 : 5—128.
Lentz T. 1967. Rhabdite Formation in Planaria: The role of microtubule.
Journ. Ultrastr. Research, 17, 1/2 : 114—126.
Luther A. 1912. Studien iiber acole Turbellarien aus dem Finnischen Meer- busen. Acta Soc. Fauna et Flora Fennica, 36 : 1—60.
Luther A. 1960. Die Turbellarien Ostfennoskandiens. 1. Acoela, Catenulida, Macrostomida, Lecithoepitheliata, Prolecithophora und Proseriata. Soc. pro Fauna et Flora Fennica, 7 : 3—155.
Luther A. 1962. Die Turbellarien Ostenfennoskandiens. III. Neorhabdocoela. 1. Dalyellioida, Typhloplanoida: Byrsophlebidae und Trigonosto- midae. Soc. pro.Fauna et Flora Fennica, 12 : 1—71.
Marcus E. 1934. Uber Lophopus crystallinus (Pall). Zool. Jahrb. Anat., 58 : 501—606.
Marcus E. 1945a. Sobre Catenulida brasileiros. Bol. Fac. Fil. Ci. Letr., Univ. Sao Paulo, Zool., 10 : 3—3133.
Marcus E. 1945b. Sobre Microturbellarios do Brasil. Comm. Zool. Mus.
Montevideo, 1 : 1—74.
Marcus E. 1946. Sobre Turbellaria brasileiros. Bol. Fac. Fil. Ci. Letr., Univ. Sao Paulo, Zool., 11 : 5—254.
Marcus E. 1948. Turbellarien do Brasil. Bol. Fac. Fil. Ci. Letr., Univ.
Sao Paulo, Zool., 13 : 111—243.
Marcus E. 1950. Turbellaria Brasileiros (8). Bol. Fac. Fil. Ci. Letr., Univ.
Sao Paulo, Zool., 15 : 5—191.
Marcus E. 1951. Turbellaria Brasileiros (9). Bol. Fac. Fil. Ci. Letr., Univ.
Sao Paulo, Zool., 1,6 : 5—215.
Marcus E. 1952. Turbellaria Brasileiros (10). Bol. Fac. Fil. Ci. Letr., Univ.
Sao Paulo, Zool., 17 : 5—187.
Marcus E. 1954. Turbellaria Brasileiros (11). Pap. Avuls. Dep. Zool. Seer. Agricult., Sao Paulo, 11, 24 : 419—489.
Marcus E. 1958. On the evolution of the animal phyla. Quart. Rev. Biol., 33 : 24—58.
Marcus E. and E. M a г с u s. 1966. Systematische Ubersicht der Polykla- den. Zool. Beitrage, N. F., 12, 3 : 319—343.
Mark E. L. 1892. Polychoerus caudatus nov. gen., nov. sp., Festschr. 2. 70 Geburtstage R. Leuckarts, Leipzig : 298—309.
Mattiesen E. 1904. Beitrag zur Embryologie der Sii pwasserdendrocoelen. Zeitschr. wiss. Zool., 77 : 274—361.
Meixner J. 1926. Beitrag zur Morphologie und zum System der Turbellarien—Rhabdocoela. II. Uber Typhlorhynchus nanus Laidlaw und die parasitischen Rhabdocoelen nebst Nachtragen zu den Caliptorhynchia. Zeitschr. Morphol. Okol. Tiere, 5 : 577—624.
Meixner J. 1938. Turbellaria (Strudelwiirmer). I. Allgemeiner Teil. Die Tierwelt Nord-Ostsee. Lief. 33, 4 : 3—146.
Meixner J. 1943. Turbellaria (Strudelwiirmer). II. System. Die Tierwelt Nord-Ostsee. Lief. 33, 4 : 147—238.
Melander Y. 1963. Cytogenetic aspects of embryogenesis in Paludicola Tricladida. Hereditas (Lund), 49 : 119—166.
Merton H. 1914. Beitrage zur Anatomie und Histologie von Temnocephala. Abh. Senckenberg. Ges., 35 : 1—58.
M о i g n e, Le A. 1963. Etude de developpement embryonnaire de Polycelis nigra (Turbellaria—Triclada). Bull. Soc. Zool. France, 88, 4 : 403—422.
Monticelli F. S. 1896. A delotacta zoologica. Mitt. Stat. Neapel, 12 : 432—462.
Mortensen T. 1912. Uber eine sessile Ctenophore, Tjalfiella tristoma Mrst. Verh. Deutsch. Zool. Ges. Leipzig, 22 : 367—372.
Muller J. 1850. Ueber eine eigentiimliche Wurmlarva aus der Klasse der Turbellarien und aus der Familie der Planarien. Muller’s Archiv Anat Phys. (1850): 485-500.
N a e f A. 1931. Phylogenie der Tiere. Handbuch der Vererbungswissenschaft. Berlin, 5, 3, 1 : 1—200.
Noirot-Timothee C. 1959. Recherches sur l’ultrastructure d’Opalina ranarum. Ann. Sci. Nat. Zool., 1 : 265—287.
Nursall J. R. 1959. The origin of the Metazoa. Trans. Roy. Soc. Canada, 53, 3, 5 : 1—5.
Nursall J. R. 1962. On the origins of the major groups of Animals. Evolution, 16, 7 : 118—123.
N у h о 1 m K. G. 1942—1944. Zur Entwicklung und Entwicklungsbiologie der Ceriantharien und Aktinien. Zool. Bidr. Uppsala, 22 : 87—248.
Palombi A. 1923. Diagnosi du nuove specie di Policladi della R. N. «Liguria». Boll. Soc. Nat. Napoli, 35 : 33—35.
P a n t i n C. F. A. 1951. Organic design. Advanc. Sci., 8 : 138—150.
P a n t i n C. F. A. 1960. Diploblastic animals. Proc. Linn. Soc. London, 171 : 1—14.
Papi F. 1950. Sulle affinita morphologiche nella Macrostomidae. Boll. Zool., Suppl. 17 : 461—468.
Papi F. 1951. Richerche sui Turbellari Macrostomidae. Arch. Zool. Ital., 36 : 289—340.
Papi F. 1953. Beitrage zur Kenntnis der Macrostomiden. Acta Zool. Fenn., 78 : 1—32.
Papi F. 1957. Sopra un nuovo Turbellario arcooforo di particolare significato filetico e sulla posizione della fam. Hofsteniidae nel sistema dei Turbellari. Publ. Staz. Zool. Napoli, 30 : 132—148.
Papi F. e B. Swedmark. 1959. Un Turbellario con lo scheletro: Acan- thomacrostomum spiculiferum n. gen., n. sp. Monit. Zool. Ital., 66, 4 : 229—250.
Pedersen K. J. 1961. Studies over ferskvands tricladernes cytology Kabenhavn : 1—94.
Pedersen K. J. 1963. Slime-sekreting cells in Planarians. Ann. N. Y. Acad. Sci., 106, 2, 4 : 424—443.
Pedersen К. J. 1964. The cellular organisation of Convoluta convoluta, on Acoel Turbellarian : a cytological, histochemical and fine structural study. Zeitschr. Zellforsch., 64, 5 : 655—687.
Pelseneer P. 1906. Molluska. In: A treatise on zoology. Ed. Ray Lanke- ster. London, 5 : 1—355.
Prudhoe G. 1945. On the species of the polyclad genus Paraplanocera. An. Mag. Nat. Hist., Ser. 11, 12 : 195—202.
Pullen E. W. 1953. The nature of the rhabdites in Stenostomum virginia- num. Virginia Journ. Sci., N. S., 4, 4 : 231.
RattenburyJ. C. 1954. The embryology of Phoronopsis viridis. Journ. Morphol., 95, 2 : 289—349.
Reisinger E. 1922—1923. Untersuchungen uber Bau und Funktion des Excretionsapparates bei rhabdocoelen Turbellaiien. Zool. Anz., 54 : 200—209; 56 : 205—224.
Reisinger E. 1923. Turbellaria. Schulze’s Biol. Tiere Deutschl., 6, 4 : 1-64.
Reisinger E. 1924. Die Gattung Rhynchoscolex. Zeitschr. Morphol. Okol. Tiere, 1 : 1—37.
Reisinger E. 1925. Untersuchungen am Nervensystem der Bothrioplana semperi Braun. Zeitschr. Morphol. Okol. Tiere, 5 : 119—149.
Reisinger E. 1926. Zur Turbellarienfauna der Antarktis. Deutsch. Siidpo- lar-Exp. 1901—1903, 18 Zool. : 415—462.<
Reisinger E. 1928—1933. Allgemeine Einleitung zur Naturgeschichte der Vermes Amera. Handb. Zool. Hrsg. W. Kiikenthal u. Th. Krumbach,
(1), 1 : 19—33.
Reisinger E. 1929. Zum Ductus Genito-Intestinalis-Problem. I. Uber primare Geschlechtstrakt-Darmverbindungen bei rhabdocoelen Turbellarien. Zeitschr. Morphol. Okol. Tiere, 16, 1, 2 : 49—73.
Reisinger E. 1933. Turbellaria der Deutschen limnologischen Sunda- Expedition. Arch. Hydrob., Stuttgart, Suppl. 12, 1 : 239—262.
Reisinger E. 1934—1935. Ergebnisse einer von E. Reisinger und O. Steinbock mit Hilfe des Rask-0rsted Fonds durchgefiihrten Reise in Gronland 1926. 6. Proporoplana jenseni n. gen., n. spec., ein morphologisch be- deutsamer Turbellarien typ. Vidensk. Medd. Dansk. naturh. Foren, 98 : 243—259.
Reisinger E. 1940. Die cytologischen Grundlagen der parthenogeneti- schen Dioogonie. Chromosoma, 1, 5 : 531—553.
Reisinger E. 1955. Karntens Hochgebirgsturbellarien. Carinthia, 2, 65 : 112-151.
Reisinger E. 1959. Anormogenetische und parasitogene Syncytienbil- dung bei Turbellarien. Protoplasma, 50 : 627—643.
Reisinger E. 1960. Was ist Xenoturbella. Zeitschr. wiss. Zool., 164: 188—198.
Reisinger E. 1961. Allgemeine Morphologie der Metazoen. Morphologie der Coelenteraten, acoelomaten und pseudocoelomaten Wiirmer. Fortschr. Zool., 13 : 1—82.
Reisinger E. 1968. Xenoprorhynchus ein Modellfall fur progressiven Funk- tionswechsel. Zeitschr. zool. Syst. Evolutionsforsch., 6 : 1—55.
Reisinger E. 1969. Ultrastrukturforschung und Evolution. Ber. Phys.- Med. Geselsch. Wurzburg. N. F., 77 : 5—47.
Reisinger E. 1970. Zur Problematik der Evolution der Coelomaten. Zeitschr. zool. Syst. Evolutionsforsch., 8, 2 : 81—109.
Reisinger E. 1972. Die Evolution des Orthogons der Spiralier und das Archicolomatenproblem. Zeitschr. zool. Syst. Evolutionsforsch., 10,
: 1-43.
Reisinger E. und S. Kelbetz. 1964. Feinbau und Entladungsmecha- nismus der Rhabditen. Zeitschr. wiss. Mikroskop., 65, 8 : 472—508.
R e m a n e A. 1950. Die Entstehung der Metamerie der Wirbellosen. Verh. Deutsch. Zool. Ges. (1949) : 16—23.
R e ni а п ё A. 1951. Die Bursa-Darmverbindung und das Problem des End- darmes bei Turbellarien. Zool. Anz., 146 : 275—291.
R e m a n e A. 1954. Die Geschichte der Tiere. In: Die Evolution der Organis- men, 2. Hrsg. G. Heberer : 340—422.
R e m a n e A. 1956. Die Grundlagen des natiirlichen Systems, der vergleichen- den Anatomie und der Phylogenetik. Aufl. 2. Leipzig : 1—364.
R e m a n e A. 1958. Zur Verwandtschaft und Ableitung der niederen Metazoen. (Stellungnahme zu den Hypothesen Steinbock’s und Hadzi’s). Verh. Deutsch. Zool. Ges. (1957) : 179—196.
R e m a n e A. 1959. Die Geschichte der Tiere. In: Die Evolution der Organis- men, 1. Hrsg. G. Heberer : 340—422.
R e m a n e A. 1963. The evolution of the Metazoa from colonial Flagellates vs. Plasmodial Ciliates. In: The lower Metazoa. Ed. E. C. Dougherty. Univ. Calif. Press : 23—32.
R e m a n e A. 1967. Die Geschichte der Tiere. In: Die Evolution der Organis- men, 1. Hrsg. G. Heberer : 589—677.
R i e d 1 R. 1954. Neue Turbellarien aus dem mediterranen Felslitoral. Zool. Jahrb., Syst., 82 : 157—244.
R i e d 1 R. 1959a. Turbellarien aus submarinen Hohlen, 1. Archoophora. Pubbl. Staz. Zool. Napoli, 30 Suppl. : 178—208.
R i e d I R. 1959b. Turbellarien aus submarinen Hohlen, 2. Prolecithophora. Pubbl. Staz. Zool. Napoli, 30 Suppl. : 209—304.
R i e d 1 R. 1969. Gnathostomulida from America. Science, 163 : 445—452.
R i x e n J. 1961. Kleinturbellarien aus Litoral der Binnengewasser Schleswig- Holsteins. Arch. Hydrobiol., 57, 4 : 464—538.
Sacks M. 1955. Observation on the embryology of Gastrotrich, Lepidodermella squamata (Dujardin, 1841). Journ. Morphol., 96, 3 : 473—495.
Salvini-Plawen L. 1968. Die Funktions-Coelomtheorie in der Evolution der Mollusken. Syst. Zool., 17, 2 : 192—208.
Salvini-Plawen L. 1969. Solenogastres und Caudofoveata (Mollusca Aculifera): Organisation und Phylogenetische Bedeutung. Malacologia,
1 : 191-216.
Schultze F. 1883. Trichoplax adhaerens nov. gen., nov. spec. Zool. Anz.,
: 92—97.*
Sedgwick A. 1884. On the origin of metameric segmentation and some other morphological questions. Quart. Journ. Micr. Sci., N. S., 24 : 43—82.
Seilern-Aspang F. 1956. Friihentwicklung eines marinen Triclade (Procerodes lobatus O. Schmidt). Arch. Entw.-Mech., 148 : 589—595.
Seilern-Aspang F. 1957a. Die Entwicklung von Macrostomum appendiculatum (Fabricius). Zool. Jahrb., Anat., 76, 3 : 311—330.
Seilern-Aspang F. 1957b. Polyembryonie als abnorme Entwicklung bei Procerodes lobata O. Schmidt (Turbellaria). Zool. Anz., 159 : 187—193.
Seilern-Aspang F. 1957c. Polyembryonie in der Entwicklung von Planaria torva (M. Schultz) auf Deckglaskultur. Zool. Anz., 159 : 193—202.
Seilern-Aspang F. 1958. Entwicklungsgeschichtliche Studien an paludicolen Tricladen. Arch. Entw.-Mech., 150 : 425—480.
S e 1 e n к a E. 1881. Zur Entwicklungsgeschichte der Seeplanarien. Biol. Centralbl., 1 : 229—239.
Slewing R. 1969. Lehrbuch der vergleichenden Entwicklungsgoschichte der Tiere. Verl. P. Parey, Hamburg u. Berlin : 1—531.
Sproston N. G. 1946. A synopsis of the monogenetic Trematodes. Transact. Zool. Soc. London, 25, 4 : 185—600.
Steinbock O. 1924. Untersuchungen iiber die GeschlechtstralU-Darm- verbindung bei Turbellarien etc., nebst einem Beitrag zur Morphologie des Tricladendarmes. Zeitschr. Morphol. Okol. Tiere, 2 : 461—504.
Steinbock O. 1927. Monographie der Prorhynchidae (Turbollaria). Zeitschr. Morphol. Okol. Tiere, 8 : 538—662.
Steinbock О. 1930—1931. Ergebnisse einef von Ё. Reisinger und 0. Steinbock mit Hilfe des Rask-0rsted Fonds durchgefiihrten Reise in Gron-
_ land 1926. 2. Nemertoderma bathycola. Vidensk. Medd. Dansk. nat. Foren, 90 :47—84.
Steinbock 0. 1937. Eine Theorie uber den plasmodialen Ursprung der Vielzeller (Metazoa). Arch. exp. Zellforsch., 19 : 343.
Steinbock 0. 1938. Uber die Stellung der Gattung Nemertoderma Steinbock im System der Turbellarien. Acta Soc. Fauna et Flora Fennica, 62, 1 : 1—28.
Steinbock 0. 1952. Keimblatterlehre und Gastraea-Theorie. Pyramide,
: 13-15, 26—31.
Steinbock 0. 1958. Schlusswort zur Diskussion Remane—Steinbock.
Verh. Deutsch. Zool. Ges. (1957) : 196—218.
Steinbock 0. 1963. Origin and affinities of the lower Metazoa: The «Acoe- loid» ancestry of the Eumetazoa. In: The lower Metazoa. Ed. E. C. Dougherty. Univ. Calif. Press : 40—54.
Steinbock 0. 1966. Die Hofsteniiden (Turbellaria Acoela). Grundsatzli- ches zur Evolution der Turbellarien. Zeitschr. zool. Syst. Evolutionsforsch., 4, 1—2 : 58—145.
Steinbock 0. und B. Ausserhoffer. 1950. Zwei grundverschie- dene Entwicklungsablaufe bei einer Art (Prorhynchus stagnalis M. Scb., Turbellaria). Arch. Entw.-Mech., 144 : 155—177.
Steinbock 0. und E. Reisinger. 1924. On Prorhynchus putealis Haswell, with a description of a new species of the genus. Quart. Journ. Micr. Sci., 68 : 443—451.
Steinmann P. 1930. Zur Orientierungssinn der Tricladen. Zeitschr. vergl. Physiol., 11, 1 : 161—172.
Steinmann P. und E. Bresslau. 1913. Die Strudelwiirmer (Turbellaria). Monogr. einheim. Tiere, 5. Leipzig : 1—380.
Sterrer W. 1966. New Polylithophorous marine Turbellaria. Nature, 210, 5034 : 436.
Sterrer W. 1969. Beitrage zur Kenntnis der Gnathostomulida. I. Anatomie und Morphologie des Genus Pterognathia Sterrer. Ark. Zool., 22, 1 : 1—125.
Stevens M. 1904. On the germ cells and the embryology of Planaria simpli- cissima. Proc. Acad., Nat. Sci. Philadelphia, 56 : 208—220.
Surface F. 1907. The early development of a Polyclad, Planocera inqui- lina. Proc. Acad. Nat. Sci., Philadelphia, 59, 3 : 514—559.
Ulrich W. 1950. Ueber die systematische Stellung einer neuen Tierklasse (Pogonofora E. K. Johansson). Sitzungsber. Deutsch. Akad. Wiss. Berlin math.-natur. Kl. (1949) : 1—25.
Ulrich W. 1951. Vorschlage zu einer Revision der Grosseinteilung des Tierreichs. Verh. Deutsch. Zool. Ges. Marburg (1950) : 215—271.
Van Beneden E. 1891. Recherches sur le developpement des Arachnac- tis. Contribution a la morphologie de Cerianthides. Arch. Biol., Paris,
: 115—146.
Westblad E. 1922. Zur Physiologie der Turbellarien. I. Die Verdauung.
Die Excretion. Acta Univ. Lund., 2, 18 : 1—212.
Westblad E. 1935. Pentacoelum fucoideum, ein neuer Typ der Turbella- ria metamerata. Zool. Anz., Ill, 3—4 : 65—82.
Westblad E. 1937. Die Turbellariengattung Nemertoderma Steinbock. Acta Soc. Fauna et Flora Fennica, 60 : 45—89.
Westblad E. 1940. Studien uber skandinavische Acoela. I. Ark. Zool., 32A (20) : 1—28.
Westblad E. 1942. Idem. II. Ark. Zool., 33A, 14 : 1—48.
Westblad E. 1945. Idem. III. Ark. Zool., 36.4, 1 : 1—56.
Westblad E. 1946. Idem. IV. Ark. Zool., 38A, 1 : 1—56.
Westblad E. 1948. Idem. V. Ark. Zool., 41A, 7 : 1—82.
Westblad E. 1949a. Xenoturbella bocki n. g., n. sp., a peculiar, primitive Turbellarian type. Ark. Zool. (ser. 2), 1 : 11—29.Westblad E. 1949b. On Meara stichopi (Bock) Westblad, a new representative of Turbellaria Archoophora. Ark. Zool. (ser. 2), 1 : 43—56.
Westblad E. 1952. Some new «Alloeocoels» (Turbellaria) from the Scandinavian West Coast. Univ. Bergen Arb., Natur. rekke, 7 : 3—27.
Westblad E. 1954. Some Hydroidea and Turbellaria from Western Nor* way with description of three new species of Turbellaria. Univ. Bergen Arb., Natur. rekke, 10 : 1—22.
Widersten B. 1965. Genital organs and fertilization in some Scyphozoa. Zool. Bidr. Uppsala, 37, 1 : 45—58.
Wietrzykowski W. 1912. Recherches sur le developpement des Lucer- naires. Arch. Zool. exp. gen., ser. 5, 10, 1 : 1—95.
Wilhelmi J. 1906. Untersuchungen iiber die Excretionsorgane der Siiss- wassertrikladen. Zeitschr. wiss. Zool., 80 : 268—272.
Wilhelmi J. 1909. Tricladen. Fauna u. Flora Golfes Neapel, 32 : 1—405.
Wilhelmi J. 1913. Platodaria, Plattiere. Еп: A. Lang’s Handb. Morphol. wirbell. Tiere. Auf. 3. Jena, 1 : 1—146.
Willi er В. H., Hyman L. H. and S. A. Rifenburgh. 1925. A histochemical study of intracellular digestion in Triclad Flatworms. Journ. Morphol., 40 : 299—340.
Wohlfarth-Bottermann К. E. 1961. Cytologische Studien. VIII. Zum Mechanismus der Cytoplasmastromung in diinnen Faden. Protoplasma, 54 : 1—26.
Wohlfarth-Bottermann К. E. 1964. Cell structures and their significance for amoeboid movement. Int. Rev. Cytol. Acad. Press, N. Y.—London, 16 : 61—131.
Woltereck R. 1904. Wurmkopf, Wurmrumpf und Trochophora. Bemer- kung zur Entwicklung und Ableitung der Anneliden. Zool. Anz., 28 : 273—322.
Woltereck R. 1905. Zur Kopffrage der Anneliden. Verh. Deutsch. Zool. Ges. (1905) : 154—186.Стр.
РЕСНИЧНЫЕ ЧЕРВИ 1
РЕСНИЧНЫЕ ЧЕРВИ (TURBELLARIA) 2
ИХ ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ 2
Органы чувств 136
/ -f-——\ j pm да 159
0X0 186
РЕСНИЧНЫЕ ЧЕРВИ 1
РЕСНИЧНЫЕ ЧЕРВИ (TURBELLARIA) 2
ИХ ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ 2
Органы чувств 136
/ -f-——\ j pm да 159
0X0 186
ЮриЁ Викторович Мамкаев
РЕСНИЧНЫЕ ЧЕРВИ (TURBELLARIA) их происхождение и эволюция Филогенетические очерки
Утверждено к печати Зоологическим институтом Академии наук СССР
Редактор издательства Л. Ф, Пухальская Художник Д. А. Андреев Технический редактор А. П. Чистякова Корректоры Л. М. Агаджанова и Л. В. Субботина
Сдано в набор 30/V 1973 г. Подписано к печати 18/IX 1973 г. Формат бумаги 60 X 90*/i«- Бумага W« 2. Печ. л. 14=14 уел. печ. л. Уч.-изд. л. 16.02. Изд. JV5 5279. Тип. зак. № 362. М-05751. Тираж 1100. Цена 1 р. 84 к.
Ленинградское отделение издательства «Наука» 199164, Ленинград, Менделеевская линия, д. 1
|
1-я тип. издательства «Наука» 199034, Ленинград, 9 линия, д. 12СтЛраница |
Строка |
Напечатано |
Должно быть |
|
45 |
22 сверху |
Gryptocelides |
Cryptocelides |
|
75 |
3 снизу |
Cyclophorus |
Cycloporus |
|
125 |
Подпись под рис. 80, 6 снизу |
monogonophora, |
monogonopora, |
|
131 |
19 » |
Ethomorhynchus |
Ethmorhynchus |
|
131 |
7 » |
A. gemelliphorus |
A. gemelliporus |
|
140 |
5 сверху |
H. monogonophora), |
H. monogonopora), |
|
188 |
Подпись под рис. 112, 4 снизу 3 » |
a—a — щупальцевая b—b — глоточная |
a—a — глоточная b—b — щупальцевая |
А.
В. Иванов и Ю. В. Мамкаев
У большинства турбеллярий имеются два боковых канала протонефридиальной системы (рис. 55). Интересное исключение составляют Catenulida с их спинным непарным протонефридием (рис. 56). Обычно каналы открываются каждый собственной порой на брюшной стороне тела (например, у. Bothrioplana), но могут впадать в глоточный карман (некоторые Neorhabdocoela) или в непарный сократимый мочевой пузырек на заднем конце тела, обра-
Рис. 56. Stenostomum tenuicauda (Catenulida). Особь, находящаяся в процессе размножения путем паратомии. (Из: Hyman, 1951).
с — мозг, со — ресничная ямка, coz — ресничные ямки зооидов, ех —протонефридий, geo —■ ганглий ресничного органа, gph —г железы глотки, in —кишка, о — ротовое отверстие, р — выделительная пора, ph —глотка, so — блюдцевидный орган.
гантские клетки, как бы нанизанные на выделительный канал, который они объёмлют своей вакуолизированной цитоплазмой со всех сторон. Они отличаются очень крупным ядром, содержащим ядрышко. Подобные паранефроциты были обнаружены также у Typhloplanoidea и Dalyellioid^a и, вероятно, вообще широко распространены у других Neorhabdocoela.
Интересно, что иногда, например у Opisthocystis goettei, канал, проходя сквозь клетку, значительно суживается, как бы сжимается
химе и снабженные пучком ресничек. Они обнаружены у многих видов бес
кишечных турбеллярий. Некоторые авторы (Marcus, 1952; Reisinger, 1961)
рассматривают мерцательные тельца как приползающие из паренхимы клетки-
заместительницы, за счет которых восстанавливаются регенерирующие участки
поврежденного эпидермиса.
7 А. В. Иванов и Ю. В. Мамкаев
1 Критический разбор этих представлений см. в работах А, В. Иванова (1967, 1968),
1 Е. Д. Логачев (1970) выделяет ленточных червей в самостоятельный тин без достаточно серьезных оснований. Принадлежность Cestoidea к Plathelminthes не вызывает сомнений и явствует прежде всего из их близости к Gyrocotyloidea и Monogenoidea (см.: Быховский, 1937, 1957).
И
1 В настоящее время единственная форма, выделявшаяся в этот отряд, относится к Prolecithophora (Karling, 1968).
1 Паренхимой плоских червей и немертин называют клеточную массу, или ткань внутренней среды, заполняющую пространство между стенкой тела и внутренними органами. Некоторые авторы (см., например: Ливанов, 1937, 1955) предпочитают называть ее мезенхимой, подчеркивая тем самым, что она всегда имеет чисто мезодермальную природу. Однако это справедливо # лишь отчасти, поскольку у Acoela центральная пищеварительная часть внутренней клеточной массы, очевидно, также гомологична эпителизованному кишечнику других турбеллярий. Клеточная масса внутренней среды у Acoela отвечает, таким образом, и мезодерме и энтодерме, вместе взятым. Это фаго- цитобласт И. И. Мечникова. Именно поэтому следует отдать предпочтение нейтральному названию «паренхима».
1По мнению Штерера (Sterner, 1969, стр. 95), эпидермис гнатостомулид ререт не жгуты, а одинотщые реснички; эпидермальные клетки гнатостомулид
он характеризует как «моноцилиарные» в отличие от «полицилиарных» клеток типичного ресничного эпителия.
Природа «ресничек», по Штереру, доказывается особенностями их локомоторных движений; каждая из них совершает «типичный удар реснички», при котором возникает поток жидкости, направленный параллельно поверхности тела. Напротив, удар жгута вызывает поток жидкости в направлении, перпендикулярном к этой поверхности. Различие между жгутом и ресничкой носит отнюдь не морфологический, а чисто функциональный характер.
Мы думаем, что эти утверждения Штерера слишком категоричны. Функциональные различия между жгутами и ресничками не всегда так определенны, как он думает. Так, многочисленные длинные жгуты жгутиконосцев из отряда Hypermastlgida (паразитов кишечника термитов и таракана Crypto- cercus) совершают согласованные ритмические движения ресничного типа. Паразитические жгутиконосцы Opalinida плавают при помощи множественных ресничкообразных жгутиков, не отличающихся функционально от типичных ресничек инфузорий. У гипермастигид и опалинид имеются слабо развитые кинодесмальные волоконца, отходящие от базальных телец (Noi- rot-Timothes, 1959). Это только подчеркивает тесное генетическое родство, существующее между жгутами и ресничками.
1 По-видимому, клеточными образованиями Н. А. Ливанов здесь называет ткани, слагающиеся из клеток.
1 Этим термином обозначают обособленные клетки, лежащие в парен
1 Два других вида Diopisthoporus обнаружены пока лишь в женском состоянии (Dorjes, 1968). Вестблад (Westblad, 1940) также наблюдал иногда в пробах только женские экземпляры D. longitubus. Это не только свидетельствует о сходном характере сексуальности всех трех видов, но и склоняет к тому, что у них происходит смена половых фаз во времени.
1 Отмеченные здесь и ранее случаи раздельнополого состояния показывают, что обособление полов не столь уж исключительно для Acoela. Исходя из приведенной выше трактовки этого явления, мы рассматриваем его как примитивный признак, свидетельствующий о низком уровне развития сексуальности. Гермафродитизм здесь еще не облигатный. У прочих турбеллярий он достигает более высокой степени развития, хотя и там известны случаи раздельнополости.
1 Однако единой терминологии для женских копулятивных органов еще не выработано.
1Под названием Macrostomum, appendiculatum nomen nudum объединялось несколько видов (см.: Беклемишев, 1951; Papi, 1951; Ах, 1959).
Ю А. В. Иванов и Ю. В. Мамкаев
1 Этим примером Акс (Ах, 1961) без достаточных оснований пытается подкрепить представление о первичности квартетного дробления.
1Мы цитируем их по реферату Гюнтера (Gunther, 1962).
1Олигомерные Coelomata, по предложению Ульриха (Ulrich, 1950), нередко называются Archicoelomata. Концепцию Альварадо о происхождении Plathelminthes можно было бы провизорно именовать архицеломатной гипотезой.
2Это название, употребленное, как нам кажется, впервые Аксом (Ах, 1960), нельзя признать удачным по той причине, что многие другие гипотезы происхождения ресничных червей также не противоречат концепции филог нетичеекого единства Spiralia.
1Акс ничего не говорит об исчезновении наружной и внутренней сегментации. Между тем, согласно теории Spiralia, этот процесс должен был происходить; более того, он осуществился независимо в каждой ветви амерных Spiralia: у Plathelminthes, Nemathelminthes и у Nemertini.
1Инвагинационная эмбриональная кишка у Acoela ошибочно описывалась С. М. Переяславцевой, а в недавнее время — С. И. Богомоловым (1960). Критические замечания по этому поводу читатель найдет в книге А. В. Иванова (1968, стр. 252).