

5. Зависимость пола дрозофилы от отношения числа х-хромосом к числу наборов аутосом (Бриджес, 1932)
|
Набор хромосом |
Фототипическое проявление пола |
Отношение числа Х-хромосом к числу наборов аутосом |
|
2А + ЗХ |
Сверхсамка |
1,5 |
|
4А + 4Х |
Нормальная самка |
1 |
|
ЗА + ЗХ |
Тоже |
1 |
|
ЗА + ЗХ + 1Y |
» |
1 |
|
2А + 2Х |
» |
Г |
|
2А + 2Х + 1Y |
» |
1 |
|
ЗА + 2Х + 1Y |
Интерсекс |
0,67 |
|
ЗА + 2Y |
Летальный |
0 |
|
ЗА + 2Х |
Интерсекс |
0,67 |
|
2А + IX + 1Y |
Нормальный самец |
0,5 |
|
2А + IX + 2Y |
» |
0,5 |
|
2А+ IX |
» |
0,5 |
|
ЗА+ IX |
Сверхсамец |
0,33 |
Нарушения в системе половых хромосом. В процессе изучения кариотипов животных были выявлены аномалии в системе половых хромосом, связанные в основном с нарушениями расхождения в мейозе, а также в результате обмена клетками разных индивидуумов. В таблице 6 дана сводка таких нарушений и их связь с фенотипом. Как видно из ее данных, нарушения в системе половых хромосом приводят к резко выраженным нарушениям воспроизводительных функций у животных.
Половой хроматин. В ядрах интерфазных клеток у нормальных особей женского пола очень часто обнаруживают небольшую глыбку хроматина, лежащую у ядерной оболочки или представленную в форме барабанной палочки, прикрепленной тонкой нитью к ядру. Барр и Бертрам, впервые обнаружившие это тельце, назвали его «сателлитом ядра». Лайон установила, что эта глыбка хроматина представляет собой одну из двух Х-хромосом, которая находится во время интерфазы в гетеропикнотическом состоянии. Положительный гетеропикноз, по гипотезе Лайон, свидетельствует о генетической инактивации одной из Х-хромосом кариотипа женского пола. Ввиду того что это тельце встречается только в ядрах клеток самок, его рассматривают как признак, отличающий клетки самок от клеток самца, и называют половым хроматином или тельцем Барра.
6. Нарушения в системе половых хромосом и их фенотипическое проявление
|
Половые хромосомы |
Виды |
Главный фенотипический эффект |
|
ХО |
Свинья, лошадь |
Интерсексуальность, гипоплазия |
|
|
|
яичников |
|
ХО/ХХ |
Кошка, лошадь |
Смерть до рождения, гипоплазия |
|
|
|
яичников |
|
XO/XX/XYXXX |
Свинья, корова |
Интерсексуальность, гипоплазия |
|
|
|
яичников |
|
XXY |
Лошадь, крупный |
Стерильность, гипоплазия |
|
|
рогатый скот, овцы, |
семенников |
|
|
свиньи, собаки, кошки |
|
|
XXY/XY |
Крупный рогатый скот |
Тоже |
|
XXY/XX |
Тоже |
Интерсексуальность |
|
|
Ч^ШШЬЯ Лошадь |
1 |
|
|
Кошка |
Тестикулярная гипоплазия |
|
XXY/XX/XY |
Крупный рогатый скот |
Тоже |
|
XXY/XY/X0 |
Тоже |
» |
|
XXY/XY/XX/XO |
Лошадь |
Крипторхизм |
|
XXXY |
» |
Интерсексуальность |
|
XXXY/XXY |
Свинья |
Не установлен |
|
XYY/XY |
Крупный рогатый скот |
» |
Этот признак можно использовать для изучения нарушений в системе половых хромосом, в диагностике пола при интерсексуальности. Количество телец Барра всегда на единицу меньше числа Х-хромосом. Так, если у самок обнаруживается два тельца Барра, то они являются носителями трисомии по Х-хромосоме. Если половой хроматин отсутствует, то у особи женского пола имеется только одна Х-хромосома. Если у самца обнаруживают тельце Барра, это значит, что у него в кариотипе не одна, а две Х-хромосомы.
Для исследования полового хроматина используют клетки лейкоцитов, слизистых полостей и др.
Интерсексуальность у животных. Влияние внутренней среды организма на изменение признаков пола в онтогенезе прослеживается при изучении интерсексуальности. У домашних животных существуют разные формы интерсексуальности, которые объединяются под названием гермафродитизм. Кроме того, обнаружены отдельные формы, очень сходные с известными синдромами Клайнфельтера и Тернера.
Образование гермафродитов — особей, имеющих гонады и (или) половые органы противоположного пола, рассматриваются как результат нарушения мейоза в период развития бластоцисты.
Синдром Клайнфельтера характеризуется недоразвитием гонад, повышением выделения гонадотропина и другими изменениями. Классической формой кариотипа синдрома Клайнфельтера является 2л (аутосом) + XXY. Однако часто в кариотипе обнаруживают мозаицизм по половым хромосомам: XXY/XY/XX и трисомию XXX (синдром «трипло-Х»). У крупного рогатого скота XXY-синдром наблюдали в сочетании с Х-три-сомией. Характерными признаками животных были нарушения роста и развития, двусторонняя гипоплазия семенников с олиго-я некроспермией и другими изменениями.
Возникновение синдрома Клайнфельтера связано с нерасхождением половых хромосом в мейозе. Примечательной особенностью фенотипа при этом синдроме является то, что животные при наличии даже двух Х-хромосом и одной Y-хромосомы имеют признаки мужского пола, хотя и стерильные. При отсутствии Y-хромосомы животные имеют женский фенотип. Это указывает на то, что Y-хромосома у животных содержит гены, контролирующие развитие мужского пола и плодовитость.
СиндромТернера характеризуется женским фенотипом с дисгенезией гонад и другими генитальными дефектами. Цито-гснетически синдром Тернера определяется формулой 2л (аутосом) + ХО (отсутствует вторая Х-хромосома).
Животных с признаками гермафродитизма, а также носителей синдромов Клайнфельтера и Тернера следует своевременно выделять в группы откорма.
Фримартинизм— особая форма интерсексуальности, выявляемая у крупного рогатого скота. Наблюдения показывают, что бесплодные телки — фримартины рождаются в двойне с бычком. Установлено, что в среднем 95 % телок из разнополых двоен имеют признаки фримартинизма. У них часто обнаруживают мужской тип экстерьера, недоразвитие матки и др.
Для объяснения причин бесплодия телок-фримартинов ученые выдвигают разные теории. Основные из них — гормональная и клеточная. Гормональная теория предполагает возникновение фримартинизма вследствие слияния плодного мешка при беременности двойней и образования анастомозов между плацентарными сосудами разнополых плодов, по которым осуществляется обмен мужскими гормонами — андрогенами (тестостерон) и женскими — эстрогенами. Поскольку мужской гормон тестостерон начинает продуцироваться раньше, то длительное воздействие на женские половые органы приводит к вирилизации (недоразвитию) последних, отклонениям в сторону мужского пола и рождению, наконец, бесплодных телок. Клеточная теория исходит из того, что при плацентарных анастомозах происходит обмен не только гормонами, но и другими элементами. Так, у телок-фримартинов был обнаружен химеризм по эритроцитар-ным антигенам и по половым хромосомам (рис. 16).
Химеризм по половым хромосомам наблюдается и у быков из разнополых двоен. У бычков на племпредприятиях частота химезма в среднем составляла 1,5 %, у отдельных из них — свыше % (А. И. Жигачев и др., 1989). Эти животные часто имеют

Рис. 16. Химеризм XX/XY у крупного рогатого скота (2n =* 60XY/60XX):
я — мегафазная пластинка с XX хромосомами; б — метафазная пластинка с XY хромосомами
нарушения воспроизводительной функции — от снижения количества спермиев в эякуляте и пониженной оплодотворяющей способности до полного бесплодия. Степень нарушения плодовитости связана, очевидно, с временем образования сосудистых анастомозов во время внутриутробного развития быков-химери-ков. Химеризм по половым хромосомам наблюдается не только в двойнях, но и в отелах коров с большим числом телят разного пола. Присутствие Y-хромосомы в кариотипе телок-фримарти-нов и изменение у них признаков в сторону мужского пола — явное свидетельство ее влияния на половые признаки. Цитогене-тический анализ — надежный метод ранней диагностики фри-мартинизма у телок. Выявление химеризма XY/XX-хромосом у быков — это критерий малопригодности их в воспроизводстве.
Химеризм по половым хромосомам обнаружен также у коз, овец, свиней, норок. В некоторых породах коз (зааненская, тог-денбургская) интерсексуальность встречается довольно часто — 6,5—8,4 %. Этот признак связан с комолостыо животных.
По данным Г. И. Исаевой и Д. К. Беляева, химеризм наиболее часто регистрировали у норок, гомозиготных или гетерозиготных по генам алеутской окраски или хедлунд
Лекция-10
НАСЛЕДОВАНИЕ ПРИЗНАКОВ, СЦЕПЛЕННЫХ С ПОЛОМ.
План: ПРОБЛЕМА РЕГУЛЯЦИИ ПОЛА
Причины более высокой смертности среди самцов млекопитающих можно объяснить исходя из особенностей наследования признаков, сцепленных с полом. Явление сцепленного с полом наследования впервые открыл Т. Морган при скрещивании мух-дрозофил с красной и белой окраской глаз. Если в скрещивании участвовали красноглазые самки и белоглазые самцы, все потомство рождалось красноглазым. Во втором же поколении наблюдалось расщепление в соотношении 3:1. Но в отличие от мендель-ского моногибридного скрещивания расщепление было только среди самцов: одна половина из них была с белыми глазами и другая — с красными; все самки были красноглазыми. При обратном (реципрокном) скрещивании белоглазых самок с красноглазыми самцами картина была иной: все самки имели красные глаза, самцы — белые. При скрещивании этих особей во втором поколении половина самцов и самок рождалась красноглазыми. Признаки, расщепление по которым при скрещивании связано с полом, называют сцепленными с полом. Эти признаки обусловливаются генами, локализованными в половых хромосомах. Установлено, что наследование их зависит в основном от Х-хромосомы. Y-Хромосома имеет небольшие размеры, состоит преимущественно из гетерохроматина и является генетически инертной, за исключением, возможно, некоторых генов, контролирующих воспроизводительную функцию и признаки пола. У самцов млекопитающих гены, локализованные в Х-хромосоме, не имеют доминантных или рецессивных партнеров (аллелей) на Y-хромо-соме. Рецессивные гены у них проявляют свое действие уже в одинарной дозе (гемизиготном состоянии) по типу доминантного.
Практическое использование сцепленного с полом наследования признаков. В птицеводстве оказалась полезной рецессивная, сцепленная с Х-хромосомой мутация карликовости. Карликовые куры отличаются от нормальных лучшей оплатой корма продукцией, для них требуется меньшая площадь содержания. Они резистентны к отдельным болезням.
В шелководстве получил распространение метод использования сцепленных с полом деталей для получения гусениц только мужского пола, дающих более крупные коконы, содержащие шелка на 25—30 % больше, чем коконы гусениц самок. Для этого В. А. Струнников вывел линию, сбалансированную по двум Z-леталям с помощью транслокации (переноса) фрагмента половой Z-хромосомы на W-хромосому. При скрещивании самцов с одной неаллельной деталью в каждой из двух хромосом (а и Ь) с самками без транслокаций рождаются только самцы:
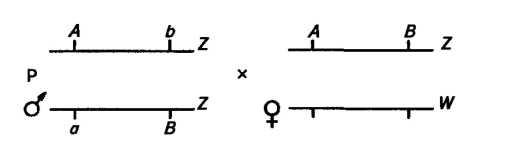
Наследственные аномалии животных, сцепленные с полом. У
сельскохозяйственных животных установлено несколько форм врожденных аномалий, обусловленных генами, локализованными в половой хромосоме. Как правило, они имеют рецессивный характер проявления, и при этом поражаются преимущественно особи гетерогамного пола: у млекопитающих — самцы, у птиц — самки. Сцепленные с полом летальные и сублетальные аномалии изменяют численное соотношение полов при рождении или после него вследствие гибели или браковки у млекопитающих самцов, у птиц — самок. Например, А. И. Жигачевым установлено, что такая аномалия, как врожденная деформация передних конечностей в сочетании с анкилозом суставов, изученная у животных черно-пестрой, сычевской и костромской пород, проявляется, как правило, у бычков, родственных между собой, что указывает на сцепленное с полом наследование.