

ТЕОРЕТИЧЕСКИЕ ВОПРОСЫ:
1. Биология: определение, современный этап развития биологии,
место и задачи биологии в системе подготовки врача.
Биология - наука изучает общие закономерности, характерные для всего живого и раскрывающие сущность жизни, её формы и развитие. Методы изучения биологии:
1)наблюдение и описание – самый старый (традиционный) метод биологии. Этот метод широко используется и в наше время (в зоологии, ботанике, цитологии, экологии и др.)
2)сравнение, т.е. сравнительный метод дает возможность найти сходства и различия, общие закономерности в строении организмов.
3)опыт или эксперимент. Например, опыты Г.Менделя или работы И.П.Павлова в физиологии.
4)моделирование – создание определенной модели или процессов и их изучения. Например, моделирование условий и процессов (недоступных наблюдению) происхождения жизни.
5)исторический метод – Биология(bios – жизнь и logos – учение) – комплексная наука, изучающая закономерности, изучение закономерности появления и развития организмов.
Выделяют 3 основных этапа в развитии биологии:
1)Этап систематики. Связан с именем Карла Линнея.
2)Эволюционный этап. Связан с именем Чарльза Дарвина.
3)Биология микромира. Связан с работа Грегора Менделя.
Врач, академик И.В. Давыдовский писал: «Медицина, взятая в плане общей теории, – это прежде всего биология». Такое утверждение основано на следующем.
1. Развитие медицины во многом связано с достижениями в области биологии:
·Исследования Луи Пастера (1862) показали невозможность самопроизвольного зарождения жизни. Он доказал, что гниение и брожение происходят благодаря наличию микроорганизмов. На основе исследований Пастера возникло в хирургии учение об асептике и антисептике.
·Исследования И.И. Мечникова (1845 – 1916), изучавшего внеклеточное пищеварение у кишечнополостных, позволили открыть фагоцитоз и стали основой для создания учения об иммунитете.
·Д.И. Ивановский (1864 – 1920) открыл вирус табачной мозаики и положил начало вирусологии и изучению целой группы заболеваний, вызываемых вирусами.
·М. Шлейден и Т. Шванн создали клеточную теорию и положили тем самым начало изучению патологии на клеточном уровне.
2. Объектом медицины является человек – существо биолого-социальное. Все структуры и функции человека являются результатом длительного исторического развития предшествующих форм жизни, поэтому многие пороки его развития имеют филогенетическую обусловленность. Следовательно, врачи должны хорошо представлять филогенез всех систем органов человека.
3. Известно более 4000 заболеваний, которые имеют отношение к наследственности полностью или частично. Поэтому врач должен хорошо знать закономерности наследственности и изменчивости, которые изучаются в курсе биологии.
4. Здоровье человека сегодня в большой степени зависит от состояния окружающей среды. Наука, изучающая взаимодействие организма с окружающей средой, – экология – также является частью биологии. 5. В курсе биологии изучаются закономерности регенерации и трансплантации, которые должны знать врачи всех специальностей. 6. Раздел практической медицины – медицинская паразитология – изучается только в курсе биологии.
7. На основе биологических наук возникли и развиваются медицинские науки. Так, на базе анатомии, цитологии и гистологии возникла патологическая анатомия; на базе физиологии, иммунологии и биохимии
– патологическая физиология; физиология и биохимия послужили основой терапии
2. Определение понятия жизнь и свойства живого. Уровни
организации живого.
Определение сущности жизни (по Энгельсу) – жизнь – это способ существования белковых тел, существенным моментом которого является постоянный обмен веществ с окружающей средой в результате которого происходит самообновление состава структур этих белковых тел. Прекращение этого обмена – прекращение жизни.
Определение жизни М.В.Волькенштейном (более современная версия): «Живые тела, существующие на Земле, представляют собой открытые саморегулирующиеся, самовоспроизводящиеся системы, построенные из полимеров – белков и нуклеиновых кислот и поддерживающие свое
существование в результате обмена веществ и энергии с окружающей средой».
Признаки жизни:
Самовоспроизведение
Саморегуляция
Самообновление Свойства жизни:
|
Питание |
|
Раздражимость |
|
Дыхание |
|
Наследственность и |
|
Движение |
изменчивость |
|
|
Выделение (экскреция) |
|
Пространственная и |
|
Репродукция |
временная организация |
|
|
Рост и развитие |
|
Дискретность и |
|
Обмен веществ |
целостность |
|
Гомеостаз
Уровни организации живого:
1.Суборганизменный уровень
Атомарный
Молекулярный
Субклеточный
Клеточный
Тканевой
Органный
2.Организменный (онтогенетический) уровень
3.Надорганизменный уровень
Популяционно-видовой
Биогеоценотический
Биосферный
3.Создание клеточной теории и ее основные положения.
Клеточная теория была сформулирована после более трехсотлетнего периода накопления знаний о строении различных одноклеточных и многоклеточных организмов растений и животных. Накопление знаний зависело от развития оптических методов исследований.
Известно, что Роберт Гук (1665) первым наблюдал с помощью увеличительных линз ткани пробки, и которые он подразделил на «ячейки», или «клетки». После его наблюдений появились
систематические исследования анатомии растений (Мальпиги, 1671; Грю, 1671), которые показали, что разнообразные части растений состоят из тесно расположенных «пузырьков», или «мешочков». В 1680 году А. Левенгук впервые увидел одноклеточные организмы и клетки животных (эритроциты). Позднее клетки животных были описаны Ф. Фонтана (1781).
С развитием микроскопирования в 19 в. изменились представления о строении клеток. Главным в организации клетки стала считаться не клеточная стенка, а собственно ее содержимое, протоплазма ( Ян Пуркинье, 1830). В протоплазме был открыт постоянный компонент клетки – ядро ( Роберт Браун, 1833). Эти и многочисленные другие наблюдения позволили в 1838 г. немецкому ученому Т. Шванну сделать ряд обобщений, в которых он показал, что клетки растений и животных принципиально сходны между собой (гомологичны).
Так в 1838-1839 году Шван и Шлейдн, независимо друг от друга, положили начало клеточной теории. Они утверждали:1) Все живые организмы состоят из клеток( кроме вирусов)
2) Клетки животных и растений схожи по строению и выполняемой функции.
Эти представления получили дальнейшее развитие в работах русского ученого Рудольфа. Вирхова (1858).
Клеточная теория – это обобщенные представления о строении клеток как единиц живого, об их размножении и роли в формировании многоклеточных организмов. Создание клеточной теории стало важнейшим событием в биологии, одним из решающих доказательств единства всей живой природы.
ОСНОВНЫЕ ПОЛОЖЕНИЯ СОВРЕМЕННОЙ КЛЕТОЧНОЙ ТЕОРИИ
1)Клетка – элементарная структурная, функциональная и генетическая единица живого.
2)Клетки растений, животных и грибов сходны по строению, химическому составу, обмену веществ.
3)Клетка – функциональная единица живого. Сходные по строению и выполняемым функциям клетки объединяются в ткани, ткани – в органы и системы органов, которые формируют целостный организм. Это обеспечивает дискретность и целостность организма.
4)Клетка – единица развития всего живого. Любой организм начинает свое индивидуальное развитие из одной клетки – зиготы.
5)Новые клетки образуются в результате деления материнской
клетки.
6)Клетка имеет мембранный принцип строения.
7)Ядро является главным компонентом эукариотической клетки, хранителем генетической информации и регуляторным механизмом.
8)Сходство строения клеток растений и животных доказывает общность их происхождения.
9)Дифференциация клеток лежит в основе усложнения строения живых организмов.
Клетка – элементарная единица живого.Первое представление о клетке как о самостоятельной жизнедеятельной единице, которая несет в себе полную характеристику жизни, было дано в работах Т. Шванна. К настоящему времени наукой это положение полностью доказано. Клетка
– это наименьшая единица живого, вне которой нет жизни.
4.Про - и эукариоты. Основные особенности их строения (примеры).
Прокариоты (от др.-греч. προ «перед» и κάρυον «ядро»), или доядерные
— одноклеточные живые организмы, не обладающие (в отличие от эукариот) оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий).
Для клеток прокариот характерно отсутствие ядерной оболочки, ДНК упакована без участия гистонов. Тип питания осмотрофный и автотрофный (фотосинтез и хемосинтез). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды.
Характерные особенности прокариот:
·Отсутствие четко оформленного ядра;
·Наличие жгутиков, плазмид и газовых вакуолей;
·Структуры, в которых происходит фотосинтез;
·Формы размножения — бесполый способ, имеется псевдосексуальный процесс, в результате которого происходит лишь обмен генетической информацией, без увеличения числа клеток;
·Размер рибосомы — 70s(по коэф. седиментации различают и рибосомы др. типов, а также субчастицы и биополимеры, входящие в состав рибосом).
Эукариоты, или ядерные (от греч. εύ- — хорошо и κάρυον — ядро) — домен (надцарство) живых организмов, клетки которых содержат ядра. Все организмы, кроме бактерий и архей, являются ядерными (вирусы и вироиды также не являются эукариотами, но не все биологи считают их живыми организмами).
Животные, растения, грибы, а также группы организмов под общим названием протисты — все являются эукариотическими организмами. Они могут быть одноклеточными и многоклеточными, но все имеют общий план строения клеток. Считается, что все эти столь несхожие организмы имеют общее происхождение, поэтому группа ядерных рассматривается как монофилетический таксон наивысшего ранга. Согласно наиболее распространённым гипотезам, эукариоты появились 1,5—2 млрд лет назад.
Эукариотические клетки в среднем намного крупнее прокариотических, разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органоиды (или органеллы, что, правда, несколько искажает первоначальное значение этого термина), из которых многие отделены от цитоплазмы одной или несколькими мембранами (в прокариотических клетках внутренние органоиды, окруженные мембраной, встречаются редко). Ядро — это часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК, «упакованные» в хромосомы. Ядро обычно одно, но бывают и многоядерные клетки.
5. Цитоплазма. Химический состав, физико-химические свойства, структурная организация. Цитоскелет.
Цитоплазмаэто внутреннее содержимое клетки. Компоненты:
Цитоплазматический матрикс(гиалоплазма)
Цитоскелет (микротрубочки, микрофиламенты, микрофибриллы)
Органоиды
Включения
Цитоплазма клеток имеет следующий состав: вода 80%, белок около 10%, ДНК 0,4%, РНК 0,7%, липиды 2%, органические соли 1% и неорганические соли 1%.
Содержимое клетки можно разделить на три части: органоиды
(обязательные для любой клетки компаненты), включения (необязательные компаненты) и гиалоплазма (основная жидкая фаза клетки – цитозоль).
Вода содержится в цитоплазме в двух видах – свободном и связанном. Свободная вода – это вода, которая может участвовать в процессах обмена веществ. Связанная вода удерживается белковыми молекулами при помощи водородных связей и образует поэтому часть структуры цитоплазмы (подсчитано, что всего 4,5 % всей воды цитоплазмы находится в связанном состоянии).
Соли содержатся во всех клетках и необходимы для процессов жизнедеятельности. Следует отметить, что из катионов наивысшей концентрации в клетке достигает калий. Содержание натрия и кальция ниже, в цитоплазме концентрация свободного кальция составляет всего 10 –7 моль/л. Преобладающим анионом в растительной клетке является хлорид. Наряду с основными солями в клетке найдены также многие другие элементы. Как уже отмечалось, характерная структура цитоплазмы определяется белками; молекулярный вес этих веществ очень высок – от 13 тыс. дальтон до многих миллионов. Своеобразные свойства белков обусловлены, вероятно, крупным размером их молекул. Все белки состоят из углерода, водорода, кислорода, азота и обычно серы, а некоторые, кроме того, содержат фосфор.
Гиалоплазма. Сложный состав. По консистенции приближается к гелю. Гели – структурированные коллоидные системы с жидкой дисперсной средой. Частицы дисперсной фазы соединены между собой в рыхлую пространственную структуру, что лишает систему текучести. Гель цитоплазмы относится к тиксотропным гелям, которые под воздействием внешних условий, которые могут менять свое агрегатное состояние и переходить в менее вязкую фазу, которая называется золь. При изменении состава микротрубочки разрушаются.
Гиалоплазма выполняет:
1)Синтез и отложение запасных полисахаридов, накопление липидов
2)Место протекания гликолиза и синтеза АТФ.
3)Место синтеза белка. Активация аминокислот с помощью специфических ферментов и связывания их с транспортными РНК.
4)В гиалоплазме происходит модификация ферментов, которая приводит к изменению функций и структуры белков.
5)Место локализации всех строительных блоков биомембран, а также всех промежуточных метаболитов.

6) Локализация всех неорганических соединений, причем концентрация неорганических веществ строго детерминирована и регулируется органоидами клетки.
Цитоскеле́т — это клеточный каркас или скелет, находящийся в цитоплазме живой клетки. Он присутствует во всех клетках эукариот, причем в клетках прокариот обнаружены гомологи всех белков цитоскелета эукариот. Цитоскелет — динамичная, изменяющаяся структура, в функции которой входит поддержание и адаптация формы клетки ко внешним воздействиям, экзо- и эндоцитоз, обеспечение движения клетки как целого, активный внутриклеточный транспорт и клеточное деление.Цитоскелет образован белками, выделяют несколько основных систем, называемых либо по основным структурным элементам, заметным при электронно-микроскопических исследованиях (микрофиламенты, промежуточные филаменты, микротрубочки), либо по основным белкам, входящим в их состав (актин-миозиновая система, кератины, тубулин-динеиновая система).
Цитоскелет выполняет три главные функции.
1.Служит клетке механическим каркасом, который придаёт клетке типичную форму и обеспечивает связь между мембранной и органеллами. Каркас представляет собой динамичную структуру, которая постоянно обновляется по мере изменения внешних условий и состояния клетки.
2.Действует как «мотор» для клеточного движения. Двигательные
(сократительные) белки содержатся не только в мышечных клетках, но и в других тканях. Компоненты цитоскелета определяют направление и координируют движение, деление, изменение формы клеток в процессе роста, перемещение органелл, движение цитоплазмы.
3. Служит в качестве «рельсов» для транспорта органелл и других крупных комплексов внутри клетки.
6.Строение и функции органоидов общего назначения: эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы, митохондрии, пластиды, рибосомы, клеточный центр, микротрубочки. Органоиды специального назначения.
Эндоплазматическая сеть: одномембранный.(Составляет 70 % клетки.) Система уплощенных мембранных мешочков–цистерн- в виде трубочек и пластинок. Образует единое целое с наружной мембраной ядерной оболочки.
Если поверхность ЭПС покрыта рибосомами, то она называется гранулярной. По цистернам такой ЭПС транспортируется белок, синтезированный на рибосомах. Гладкая ЭПС (без рибосом) служит местом синтеза липидов и стероидов Комплекс Гольджи: одномембранный. Представляет собой стопку
уплощенных мешочков-цистерн, на коцах которых имеются отшнуровывающиеся участки в виде пузырьков.
Структурной единицей является диктиосома-стопка уплощенных цистерн. Является бункером клетки, так как в него поступают белки( синтезируется на рибосомах, затем идет в ЭПС, а после накапливается в комплексе).
Основная функция: формирование лизосом.
Лизосомы: Одномембранный. Простой сферический мешочек, образуется в комплексе Гольджи, заполнен пищеварительным (гидролитическим) ферментом. Функция: растворение «балласта» клетки.
Перокрисомы: Одномембранные. Являются разновидностью лизосом. С помощью ферментов (каталаза и пероксидаза) удаляют перикись. Митохондрии: Двумембранные. (автономные органоиды). Окружена оболочкой из двух мембран, внутренняя мембрана образует складки (кристы). Содержит собственные рибосомы и кольцевую ДНК. Функция: синтез АТФ.
Пластиды: Двумембранные. Содержатся только в растительных клетках. Различают: Хлоропласты, Хромопласты, Лейкопласты. Имеются телокоиды, которые образуют граны, содержащие пигмент хлорофил. Внутренне содержимое представлено стромой. Функция:участие в фотосинтезе.
Рибосомы: Немембранные. Очень мелкие органеллы состоящие из двух субъединиц( большой и малой). Которые соеденины между собой с помощью функционального центра рибосомы. Образуются в ядрышках ядра. Много рибосом, нанизанных на нить матричной РНК образуют полирибосому Ф-я: Синтез белка.
Клеточный центр: Немембранный. Главный центр организации микротрубочек, которые образуют цилиндр. Лучистая сфера образуется за счет белка тубулина. Функция: образование нитей веретена деления и участие в клеточном цикле эукариот.
Микротрубочки: Немембранные. Белковые внутриклеточные структуры, входящие в состав цитоскелета.Микротрубочки представляют собой полые цилиндры диаметром 25 нм. Длина их может быть от нескольких микрометров до, вероятно, нескольких миллиметров в аксонах нервных клеток. Их стенка образована димерами тубулина. Микротрубочки, подобно актиновым микрофиламентам, полярны: на одном конце
происходит самосборка микротрубочки, на другом — разборка. В клетках микротрубочки играют роль структурных во многих клеточных процессах.
Органоиды специального назначения: Органоиды специального назначения содержатся во многих растительных и животных клетках. К ним относятся органоиды движения (миофибриллы, реснички, жгутики, стрекательные капсулы и др.), опорные структуры (тонофибриллы), органоиды, воспринимающие внешние раздражения (например, фоторецепторы, статорецепторы и фонорецепторы), нейрофибриллы, а также структуры клеточной поверхности, связанные с всасыванием и перевариванием пищи (микроворсинки, кутикула и др. виды.)
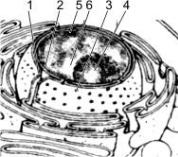
7.Строение и функции ядра.
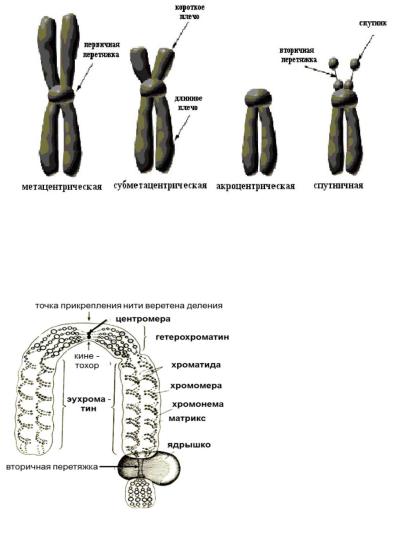
Как правило, эукариотическая клетка имеет одно ядро, но встречаются двуядерные (инфузории) и многоядерные клетки (опалина). Некоторые высокоспециализированные клетки вторично утрачивают ядро (эритроциты млекопитающих, ситовидные трубки покрытосеменных). Форма ядра — сферическая, эллипсовидная, реже лопастная, бобовидная и др. Диаметр ядра — обычно от 3 до 10 мкм.
Строение ядра:
1 — наружная мембрана; 2 — внутренняя мембрана; 3 — поры; 4 — ядрышко; 5 — гетерохроматин; 6 — эухроматин.
Ядро отграничено от цитоплазмы двумя мембранами (каждая из них имеет типичное строение). Между мембранами — узкая щель, заполненная полужидким веществом. В некоторых местах мембраны сливаются друг с другом, образуя поры (3), через которые происходит обмен веществ между ядром и цитоплазмой. Наружная ядерная (1) мембрана со стороны, обращенной в цитоплазму, покрыта рибосомами, придающими ей шероховатость, внутренняя (2) мембрана гладкая. Ядерные мембраны являются частью мембранной системы клетки: выросты наружной ядерной мембраны соединяются с каналами эндоплазматической сети, образуя единую систему сообщающихся каналов.
Кариоплазма (ядерный сок, нуклеоплазма) — внутреннее содержимое ядра, в котором располагаются хроматин и одно или несколько ядрышек. В состав ядерного сока входят различные белки (в том числе ферменты ядра), свободные нуклеотиды.
Ядрышко (4) представляет собой округлое плотное тельце, погруженное в ядерный сок. Количество ядрышек зависит от функционального состояния ядра и варьирует от 1 до 7 и более. Ядрышки обнаруживаются только в неделящихся ядрах, во время митоза они исчезают. Ядрышко образуется на определенных участках хромосом, несущих информацию о структуре рРНК.
Хроматин — внутренние нуклеопротеидные структуры ядра, окрашивающиеся некоторыми красителями и отличающиеся по форме от ядрышка. Хроматин имеет вид глыбок, гранул и нитей. В зависимости от функционального состояния хроматина различают: гетерохроматин (5) и эухроматин (6). Эухроматин — генетически активные, гетерохроматин — генетически неактивные участки хроматина. Эухроматин при световой микроскопии не различим, слабо окрашивается и представляет собой деконденсированные (деспирализованные, раскрученные) участки хроматина. Гетерохроматин под световым микроскопом имеет вид глыбок или гранул, интенсивно окрашивается и представляет собой конденсированные (спирализованные, уплотненные) участки хроматина. Хроматин — форма существования генетического материала в интерфазных клетках. Во время деления клетки (митоз, мейоз) хроматин преобразуется в хромосомы.
Функции ядра: 1) хранение наследственной информации и передача ее дочерним клеткам в процессе деления, 2) регуляция жизнедеятельности клетки путем регуляции синтеза различных белков, 3) место образования субъединиц рибосом.
8.Состав, строение, свойства и функции клеточных мембран.
Состоит из билипидного слоя (представленного фосфолипидами), пронизанного молекулами белков. Различают Интегральные, полуинтегральные и структурные молекулы белка. На наружной поверхности мембраны находится гликокаликсгликопротеиновый слой, выполняющий рецепторную функцию.
Функции клеточных мембран :
Барьерная (обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой)
Транспортная (осуществляется перенос веществ ИЗ клетки и В
клетку)
Матричная (Обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие)
Механическая (Обеспечивает автономность клетки, ее структур, и соединений с другими клетками)
Энергетическая (При фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки)
Рецепторная (некоторые белки, находящиеся в мембране, являются рецепторами)
Ферментативная (мембранные белки нередко являются ферментами)
Маркировка клеткина мембране есть антигены, действующие как маркеры,- «ярлыки», позволяющие опознать клетку. Это гликопротеины, играющие роль «антенн»
9.Виды пассивного и активного мембранного транспорта. Осмос, осмотические свойства клеток, диализ.
Молекулы веществ, диаметр которых не превышает 50 нм, могут проникать путем пассивного и активного транспорта через поры в структуре мембраны. Крупные вещества попадают в клетку путем эндоцитоза (транспорт в мембранной упаковке), требующего затраты энергии. Его разновидностями являются фаго- и пиноцитоз.
Пассивный транспорт – вид транспорта, в котором перенос веществ осуществляется по градиенту химической или электрохимической концентрации без затраты энергии АТФ. Выделяют два вида пассивного транспорта: простая и облегченная диффузия. Диффузия – это перенос ионов или молекул из зоны более высокой их концентрации в зону более низкой концентрации, т.е. по градиенту.
Простая диффузия – ионы солей и вода проникают через трансмембранные белки или жирорастворимые вещества по градиенту концентрации.
Облегченная диффузия – специфические белки-переносчики связывают вещество и переносят его через мембрану по принципу «пинг-понга». Таким способом через мембрану проходят сахара и аминокислоты. Скорость такого транспорта значительно выше, чем простой диффузии. Кроме белковпереносчиков, в облегченной диффузии принимают
участие некоторые антибиотики – например, грамитидин и ваномицин. Поскольку они обеспечивают транспорт ионов, их называют ионофорами. Активный транспорт – это вид транспорта, при котором расходуется энергия АТФ, он идёт против градиента концентрации. В нем принимают участие ферменты АТФ-азы. В наружной клеточной мембране находятся АТФ-азы, которые осуществляют перенос ионов против градиента концентрации, это явление называется ионным насосом. Примером является натрий-калиевый насос. В норме в клетке больше ионов калия, во внешней среде – ионов натрия. Поэтому по законам простой диффузии калий стремится из клетки, а натрий – в клетку. В противовес этому натрий-калиевый насос накачивает против градиента концентрации в клетку ионы калия, а ионы натрия выносит во внешнюю среду. Это позволяет поддерживать постоянство ионного состава в клетке и её жизнеспособность. В животной клетке одна треть АТФ расходуется на работу натрий-калиевого насоса.
Разновидностью активного транспорта является транспорт в мембранной упаковке – эндоцитоз. Крупные молекулы биополимеров не могут проникать через мембрану, они поступают в клетку в мембранной упаковке. Различают фагоцитоз и пиноцитоз. Фагоцитоз – захват клеткой твердых частиц, пиноцитоз – жидких частиц. В этих процессах выделяют стадии:
1) узнавание рецепторами мембраны вещества; 2) впячивание (инвагинация) мембраны с образованием везикулы (пузырька); 3) отрыв пузырька от мембраны, слияние его с первичной лизосомой и восстановление целостности мембраны; 4) выделение непереваренного материала из клетки (экзоцитоз).
Эндоцитоз является способом питания для простейших. У млекопитающих и человека имеется ретикуло-гистио-эндотелиальная система клеток, способная к эндоцитозу – это лейкоциты, макрофаги, клетки Купфера в печени.
ОСМОТИЧЕСКИЕ СВОЙСТВА КЛЕТКИ Осмос – односторонний процесс проникновения воды через
полупроницаемую мембрану из области с меньшей концентрацией раствора в область с более высокой концентрацией. Осмос обусловливает осмотическое давление.
Диализ – односторонняя диффузия растворенных веществ.
Раствор, в котором осмотическое давление такое же, как и в клетках, называют изотоническим. При погружении клетки в изотонический раствор её объем не изменяется. Изотонический раствор называют физиологическим – это 0,9% раствор хлорида натрия, который широко
применяется в медицине при сильном обезвоживании и потери плазмы крови.
Раствор, осмотическое давление которого выше, чем в клетках, называют гипертоническим. Клетки в гипертоническом растворе теряют воду и сморщиваются. Гипертонические растворы широко применяются в медицине. Марлевая повязка, смоченная в гипертоническом растворе, хорошо впитывает гной.
Раствор, где концентрация солей ниже, чем в клетке, называют гипотоническим. При погружении клетки в такой раствор вода устремляется в нее. Клетка набухает, ее тургор увеличивается, и она может разрушиться. Гемолиз – разрушение клеток крови в гипотоническом растворе.
Осмотическое давление в организме человека в целом регулируется системой органов выделения.
10.Понятие о жизненном, клеточном и митотическом цикле клетки. Интерфаза, виды интерфаз. Периоды аутосинтетической интерфазы
Жизненный цикл клетки(ЖЦК)-это период с момента ее появления до следующего деления или гибели ЖЦК= митоз+ Гетеро-синтетическая интерфаза+ Аутосинтетическая интерфаза
Клеточный цикл(КЦ)- период подготовки клетки к делению и само деление.
КЦ= Аутосинтетическая интерфаза+ митоз Митотический цикл клетки(МЦК)-все процессы митоза.
Интерфазастадия жизненного цикла клетки между двумя последовательными митотическими делениями.
Виды интерфаз:
o Аутосинтетическая-происходит подготовка клетки к следующему делению.
G1-Предсинтетический или постмитотический периодклетка интенсивно растет.
S- Синтетический период-(если клетка не делится, то у нее нет этого периода(пример-нервная клетка))- происходит удвоение ДНК
G2-Предмитотический или постсинтетический периодклетка накапливает АТФ и тубулин-готовиться к делению.
11. Митоз. Его сущность, фазы, биологическое значение. Амитоз.

Митознепрямое, полноценное деление соматических клеток. Обеспечивает формирование дочерних клеток с таким же набором хромосом, как и материнская (2n).
Изучают на стадии метафазы. Профаза:
Хроматин спирализуется
Растворяются ядрышки
Растворяется ядерная оболочка
Центриоли расходятся к полюсам клетки и начинается формирование нитей веретена деления.
Метафаза:
Заканчивается формирование нитей веретена деления
Хромосомы выстраиваются на экваторе клетки, образуя метафазную пластинку.
Анафаза:
Дочерние хромосомы расходятся к полюсам клеток. Телофаза:
Хромосомы деспирализуются
Появляется ядро и ядрышкинити веретена деления разрушаются, обеспечивается цитокинез.
Разделение цитоплазмы на две дочерние клетки
Образование ядерной оболочки
В Телофазе происходят процессы обратные профазе.
Амитозпрямое неполноценное деление клетки. (Способ образования злокачественных новообразований). Деление клеток простым делением ядра на пополам.
Нити веретена деления не образуются, ядро делится пополам случайным образом, генетический материал распределяется не равномерно.
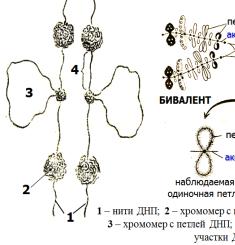
12. Мейоз. Стадии, биологическое значение.
Мейозпроцесс образования половых клеток. Биологическое значение:
1)является основным этапом гаметогенеза;
2)обеспечивает передачу генетической информации от организма к организму при половом размножении;
3)дочерние клетки генетически не идентичны материнской и между собой.
Первое деление (редукционное)
Профаза I
По-другому этот этап называют стадией тонких нитей.
лептотена
Хромосомы выглядят в микроскопе как спутанный клубок. Иногда выделяют пролептотену, когда отдельные ниточки еще сложно разглядеть.
Стадия сливающихся нитей. Гомологичные, то есть сходные между собой по морфологии и в генетическом отношении,
зиготена
пары хромосом сливаются. В процессе слияния, то есть конъюгации, образуются биваленты, или тетрады. Так называют довольно устойчивые комплексы из пар хромосом.
Стадия толстых нитей. На этом этапе хромосомы спирализуются и завершается репликация ДНК, образуются
пахитена
хиазмы - точки контакта отдельных частей хромосом - хроматид. Происходит процесс кроссинговера. Хромосомы перекрещиваются и обмениваются некоторыми участками генетической информации.
Также называется стадией двойных нитей. Гомологичные диплотена хромосомы в бивалентах отталкиваются друг от друга и
остаются связанными только в хиазмах.
диакинез |
На этой стадии биваленты расходятся на периферии ядра. |
Метафаза
Оболочка ядра разрушается, формируется веретено
I
деления. Биваленты перемещаются к центру клетки и выстраиваются вдоль экваториальной плоскости.
Биваленты распадаются, после чего каждая хромосома из Анафаза I пары перемещается к ближайшему полюсу клетки.
Разделения на хроматиды не происходит.
Завершается процесс расхождения хромосом. Происходит формирование отдельных ядер дочерних клеток, каждое - с
Телофаза I гаплоидным набором. Хромосомы деспирализуются, образуется ядерная оболочка. Иногда наблюдается цитокинез, то есть деление самого тела клетки.
Второе деление (эквационное)

|
Происходит конденсация хромосом, клеточный центр |
|
Профаза II |
делится. Разрушается ядерная оболочка. Образуется |
|
|
веретено деления, перпендикулярное первому. |
|
|
|
|
Метафаза |
В каждой из дочерних клеток хромосомы выстраиваются |
|
II |
вдоль экватора. Каждая из них состоит из двух хроматид. |
|
|
|
|
Анафаза II |
Каждая хромосома делится на хроматиды. Эти части |
|
расходятся к противоположным полюсам. |
||
|
||
|
|
|
Телофаза II |
Полученные однохроматидные хромосомы |
|
деспирализуются. Образуется ядерная оболочка. |
||
|
|
13. Гаметогенез: ово - и сперматогенез.
Гаметогенез – это последовательный процесс, который обеспечивает размножение, рост и созревание половых клеток в мужском организме (сперматогенез) и женском (овогенез).
Гаметогенез протекает в половых железах - сперматогенез в семенниках у мужчин, а овогенез в яичниках у женщин. В результате гаметогенеза в организме женщины образуются женские половые клетки - яйцеклетки, а у мужчин - мужские половые клетки сперматозоиды. Именно процесс гаметогенез (сперматогенез, овогенез) дает возможность мужчине и женщине возможность воспроизведения потомства.
Характеристика овогенеза и сперматогенеза Гаметогенез имеет несколько стадий. Сходство сперматогенеза и
овогенеза как раз и заключается в том, что три стадии у них одинаковы.
1.Стадия размножения. Первичные клетки на этой стадии называются сперматогониями и овогониями, из них в последующем образуются мужские и женские половые клетки. Половые клетки несколько раз делятся путем митоза, и количество их значительно возрастает. Сперматогонии размножаются у мужчины в течение всего репродуктивного периода, а размножение овогоний происходит в эмбриональном периоде и наиболее интенсивно происходит во 2 - 5 месяц внутриутробного развития.
2.Стадия роста. В этот период клетки значительно увеличиваются в размерах. Сперматогонии и овогонии превращаются в сперматоциты и овоциты I порядка. Овоциты I порядка достигают больших размеров, поскольку накапливают питательные вещества.
3.Стадия созревания. На этой стадии происходят два следующих друг за другом деления - мейоз I и мейоз II. После первого деления образуются
сперматоциты и овоциты II порядка, а после второго деления - сперматиды и зрелые яйцеклетки с тремя полярными тельцами, которые в процессе размножения не участвуют и погибают. При созревании один сперматоцит I порядка дает четыре сперматиды, а один овоцит I порядка образует одну яйцеклетку и три полярных тельца.
Эти особенности сперматогенеза и овогенеза имеют биологический смысл, который связан с разным назначением мужских и женских гамет. Неравномерное деление клеток при овогенезе (меньше) обеспечивает формирование крупной яйцеклетки, в ней накапливается большее количество питательных веществ, так как из оплодотворенного яйца будет развиваться новый организм.
При сравнительной характеристике овогенеза и сперматогенеза можно заметить, что сперматозоидов образуется значительно больше, и это также имеет биологический смысл. Яйцеклетку достигает только один сперматозоид, проникает в нее и доставляет свой набор хромосом. Остальные же в процессе поиска яйцеклетки массово погибают. При сравнении овогенеза и сперматогенеза становится понятным, почему сперматозоидам нет необходимости в запасании питательных веществ - их существование кратковременно, а подвижность должна быть высокой. 4. Стадия формирования. Она характерна только для сперматогенеза. Незрелая сперматида превращается в сперматозоид, приобретая свойственный ему вид. Образование сперматозоидов у мужчин начинается только в период полового созревания и происходит в течение всего года. Период развития сперматогоний в зрелые сперматозоиды составляет 74 дня.
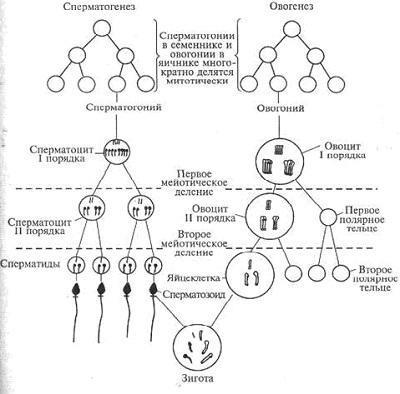
Схема овогенеза и сперматогенеза
Краткую, обобщенную характеристику овогенеза и сперматогенеза представляет следующая таблица сперматогенеза и овогенеза:
признаки |
сперматогенез |
овогенез |
Половые железы, |
Яички, сперматозоиды |
Яичники, яйцеклетки |
половые клетки |
|
|
Характерные |
|
|
особенности половых |
|
|
клеток: |
55 мк |
130-169 мк |
- относительные |
подвижные |
неподвижные |
размеры |
округлые со жгутиком |
округлые |
- подвижность |
отсутствует |
имеется |
- форма |
|
|
- наличие питательных |
|
|
веществ |
|
|
Особенности развития |
путем митоза |
путем митоза |
половых клеток на |
образуются |
образуются овоциты; |
разных стадиях: |
сперматоциты; |
увеличиваются в |
- стадия размножения |
увеличиваются в |
размерах; |
|
размерах; |
путем мейоза |
- стадия роста |
путем мейоза |
образуются гаплоидные |
- стадия созревания |
образуются гаплоидные |
овоциты, их которых |
|
сперматиды, их которых |
формируется яйцеклетка |
|
формируются |
|
|
сперматозоиды |
|
|
|
|
Подытоживая все, можно привести сравнительную характеристику овогенеза и сперматогенеза. В сущности, это и будут особенности сперматогенеза и овогенеза.
1.Мы выяснили, что гаметогенез включает стадии размножения, роста и созревания клеток. Сперматогенез включает также стадию формирования (ее нет при овогенезе), в этом заключаются отличия сперматогенеза от овогенеза. Сперматозоиды проходят дополнительную четвертую стадию для того, чтобы приобрести своеобразную форму и сформировать аппарат движения.
2.Второе отличие сперматогенеза от овогенеза: из сперматоцита I порядка получается четыре половых клетки, а из ооцита I порядка получается одна полноценная половая клетка.
3.Яйцеклетки образуются циклически, процесс повторяется через 21-35 дней (менструальный цикл). После гибели яйцеклетки, что сопровождается кровотечением, изменившийся гормональный фон дает толчок к созреванию другой яйцеклетки. Сравнительная характеристика овогенеза и сперматогенеза показывает, что у женщин мейоз начинается в период внутриутробного развития. Ооциты I порядка у новорожденной девочки останавливаются в фазе мейоз I, и завершается созревание ооцита к моменту полового созревания. У мальчиков процесс образования сперматозоидов идет непрерывно и сохраняется в течение всей половой зрелости мужчины.
4.Из характеристики овогенеза и сперматогенеза следует, что существуют значительные различия в количестве образованных половых клеток в женском и мужском организме. Взрослый мужчина производит 30 миллионов спермиев в день, а женщина - порядка 500 зрелых яйцеклеток за всю свою жизнь.
5.Различия сперматогенеза и овогенеза заключаются также в том, что стадия размножения при сперматогенезе идет постоянно, а при овогенезе заканчивается после рождения.
6.Стадия роста при сперматогенезе короче, чем при овогенезе.
7.Стадия созревания при овогенезе имеет особенности, которые заключаются в неравномерности делений при созревании, что приводит к выделению полярных телец, что отсутствует при сперматогенезе.
8.Различия сперматогенеза и овогенеза заключаются в том, что сперматогенез более подвержен влиянию внешней среды, нежели овогенез, что связано с различием в расположении половых органов - семенники находятся вне брюшной полости.
9.Из сравнительной характеристики овогенеза и сперматогенеза можно увидеть, что, поскольку образование яйцеклеток начинается еще до рождения девочки, а завершается для яйцеклетки только после ее оплодотворения, то неблагоприятные факторы внешней среды могут повлечь генетические аномалии у потомства.
14.Хромосомы. Их химический состав, надмолекулярная организация (уровни упаковки ДНК).
Хромосомы — это цитологические палочковидные структуры, представляющие собой конденсированный хроматин и появляющиеся в клетке во время митоза или мейоза.
Основу хромосомы составляет одна непрерывная двухцепочечная молекула ДНК; длина ДНК одной хромосомы может достигать нескольких сантиметров. Понятно, что молекула такой длины не может располагаться в клетке в вытянутом виде, а подвергается укладке, приобретая определенную трехмерную структуру, или конформацию.
Хромосомы: 1 — метацентрическая; 2 — субметацентрическая; 3, 4 — акроцентрические. Строение хромосомы: 5 — центромера; 6
— вторичная перетяжка; 7 — спутник; 8 — хроматиды; 9 — теломеры
.
Метафазная хромосома (хромосомы изучаются в метафазу митоза) состоит из двух хроматид (8). Любая хромосома имеет первичную перетяжку (центромеру) (5), которая делит хромосому на плечи.
Некоторые хромосомы имеют вторичную перетяжку (6) и спутник (7). Спутник — участок короткого плеча, отделяемый вторичной перетяжкой. Хромосомы, имеющие спутник, называются спутничными (3). Концы

хромосом называются теломерами (9). В зависимости от положения центромеры выделяют: а) метацентрические (равноплечие) (1), б) субметацентрические (умеренно неравноплечие) (2), в) акроцентрические (резко неравноплечие) хромосомы (3, 4).
Соматические клетки содержат диплоидный (двойной — 2n) набор хромосом, половые клетки — гаплоидный (одинарный — n). Диплоидный набор аскариды равен 2, дрозофилы — 8, шимпанзе — 48, речного рака
— 196. Хромосомы диплоидного набора разбиваются на пары; хромосомы одной пары имеют одинаковое строение, размеры, набор генов и называются гомологичными.
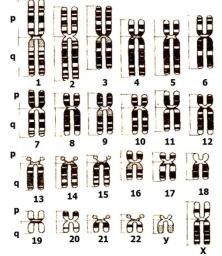
Кариотип — совокупность сведений о числе, размерах и строении метафазных хромосом. Идиограмма — графическое изображение кариотипа. У представителей разных видов кариотипы разные, одного вида — одинаковые. Аутосомы — хромосомы, одинаковые для мужского
иженского кариотипов. Половые хромосомы — хромосомы, по которым мужской кариотип отличается от женского.
Хромосомный набор человека (2n = 46, n = 23) содержит 22 пары аутосом
и1 пару половых хромосом. Аутосомы распределены по группам и пронумерованы:
Половые хромосомы не относятся ни к одной из групп и не имеют номера. Половые хромосомы женщины — ХХ, мужчины — ХУ. Х- хромосома — средняя субметацентрическая, У-хромосома — мелкая акроцентрическая.
Функции хромосом: 1) хранение наследственной информации, 2) передача генетического материала от материнской клетки к дочерним. Химический состав хромосом.
Выделяют 3 уровня структурной организации хромосом.
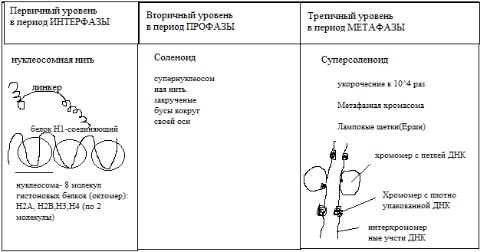
Молекула ДНК в хромосомах тесно связана с двумя классами белков - гистонами (основные белки) и негистонами (кислые белки).
Гистоны - это небольшие по величине белки с высоким содержанием заряженных аминокислот (лизина и аргинина).
Суммарный положительный заряд позволяет гистонам связываться с ДНК независимо от нуклеотидного состава. Им принадлежит в основном структурная функция. Это очень стабильные белки, молекулы которых могут сохраняться в течение всей жизни клетки.
В эукариотической клетке присутствуют 5 типов гистонов, которые распределяются на две основные группы: первая группа (их обозначают как Н2А, Н2В, НЗ, Н4) отвечает за формирование специфических дезоксирибонуклеопротеидных комплексов - нуклеосом. Вторая группа гистонов (H1) располагается между нуклеосомами и фиксирует укладку нуклеосомной цепи в более высокий уровень структурной организации (супернуклеосомную нить).
Среди гистоновых белков, кроме структурных, встречаются такие, которые способны ограничивать доступность ДНК для ДНК - связывающих регуляторных белков и тем самым участвовать в регуляции активности генов.
Негистоновые белки весьма разнообразны. Число их фракций превышает 100. Они присутствуют в меньших количествах в хромосомах в сравнении с гистонами и выполняют в основном регуляторную функцию. Участвуют в регуляции транскрипционной активности генов, в обеспечении редупликации и репарации ДНК.
Большинство негистоновых белков хроматина присутствуют в клетках в
небольшом количестве (минорные) - это регуляторные белки, узнающие
специфические последовательности ДНК и связывающиеся с ними. Они вовлечены во многие генетические процессы, но известно о них пока немного. Количественно преобладают негистоновые белки (мажорные), высокоподвижные, относительно малого размера, с большим электрическим зарядом - они всегда соединяются с нуклеосомами, содержащими активные гены. Кроме того, в группу негистоновых белков входит много ферментов.
Надмолекулярная организация хромосом
Надмолекулярная организация хромосом называется еще или спирализацией, или конденсацией, или компактизацией.
В настоящее время принято три уровня надмолекулярной организации хромосом: первичный, вторичный, третичный.
Компактизация ДНК для эукариотической клетки важна по двум причинам: она позволяет не запутать и упорядоченно расположить очень длинные молекулы ДНК в небольшом объеме клеточного ядра и, кроме того, это один из способов функционального контроля генов - характер упаковки ДНК влияет на активность некоторых участков генома.
Первичный уровень надмолекулярной организации — нуклеосомный
(рис.4). Элементарной структурой хромосомы, различаемой с помощью электронного микроскопа, является нить диаметром 10-13 нм, представляющая собой комплекс ДНК и гистоновых белков. Эта нить состоит из гистонового остова (в виде цепочки расположенных друг за другом белковых телец дисковидной формы), поверх которого спирально закручена нить ДНК. Комплекс ДНК и гистонов на уровне одного дисковидного тельца называется нуклеосомой. Она содержит по две молекулы каждого из 4-х типов гистона (Н2А, Н2В, НЗ, Н4), соединенных в форме октамера. ДНК в нуклеосоме лежит поверх октамера, накручиваясь спирально на гистоновый остов. На уровне каждой нуклеосомы ДНК образует 2,3 оборота спирали, что соответствует примерно 200 парам нуклеотидов. Связь между соседними нуклеосомами осуществляется за счет гистона H1. На этот связывающий участок приходится 60 пар нуклеотидов. Формируется нить диаметром примерно 11 нм.
Нуклеосома - это универсальная частица, которая обнаруживается как в эухроматине, так и в гетерохроматине, в интерфазном ядре и метафазных хромосомах.
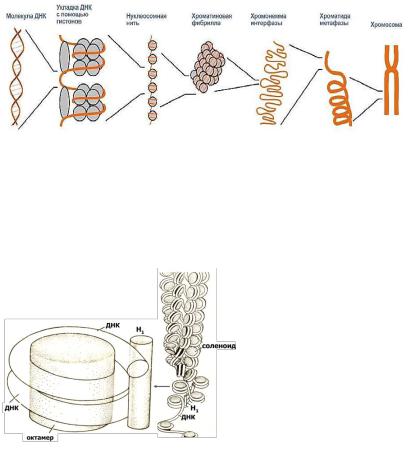
Рисунок 4. Уровни надмолекулярной организации хромосом.
В случае линейной выпрямляемости, которая едва ли присутствует в живой клетке, образуемая нуклеосомами структура напоминает нитку "бус" и называется нуклеосомной нитью. Благодаря нуклеосомной организации хромосом происходит укорочение исходной длины ДНК в 7раз, т.е. происходит компактизация. Это, видимо, состояние
|
интерфазной |
|
хромосомы, |
|
ее |
||
|
эухроматиновых участков. |
|
|
||||
|
Дальнейшая компактизация ДНК в |
||||||
|
составе |
хромосом связана |
с |
||||
|
образованием |
наднуклеосомных |
|||||
|
структур. Так, вторичный уровень |
||||||
|
хромосомной |
|
укладки |
ДНК |
|||
|
выражается |
в |
формировании |
||||
|
суперспиральной |
нити |
|||||
|
(соленоида), в которой исходная |
||||||
Рисунок 5. Строение соленоида. |
молекула |
ДНК |
укорачивается |
в |
|||
40раз. Толщина достигает 30-40 |
|||||||
|
|||||||
|
нм. |
При |
|
образовании |
|||
суперспирали нуклеосомная нить спирально закручивается за счет взаимодействия гистонов HI и НЗ. Не исключено также и участие в этом негистоновых белков (рис. 5). Этот уровень укладки ДНК соответствует, по-видимому, наблюдаемым под световым микроскопом профазным митотическим и мейотическим хромосомам. Или интерфазным, но не транскрибируемым, возможно, участкам хромосом, т. е. гетерохроматиновым.
Третий уровень хромосомной укладки изучен менее всего.
Существует две модели: в основу первой положен принцип спиральной укладки, в основе второй - строение по принципу складывания петель. В последние годы накоплен многочисленный материал, говорящий о реальности петлеобразных структур в хромосоме и их плотной упаковке в метафазной хромосоме вокруг осевого каркаса, построенного из

негистоновых белков. Петлевые структуры, но не плотно упакованные, есть и в интерфазной хромосоме. Вокруг каркаса, как в щетке-ерше, располагаются петли суперспиральной нити. Причем концы каждой петли локализуются на одной и той же точке белкового каркаса. Предполагается также, что петли могут скручиваться вокруг своей собственной оси, т.е. метафазную хромосому можно изобразить в виде плотно уложенных соленоидных петель, свернутых в тугую спираль. Типичная хромосома человека может содержать до 2600 петель (рис. 6).
Третий
уровень укладки -
это
конденсаци
я
профазной
хромосомы
в
метафазну ю. Толщина такой структуры достигает
1400 нм
(две хроматиды), а молекула
ДНК при этом укорачивается в I04 раз, т.е. с 5 см растянутой ДНК до 5 мкм. Эта суперспирализация сопровождается фосфорилированием в клетке всех молекул HI. В любом случае ДНК в ядрах эукариотических клеток образует иерархическую систему спиралей и петель, основной единицей которой является нуклеосома. Нуклеосомы, в свою очередь, расположены не везде строго одинаково. Эти малозаметные и малоизученные различия биологически очень важны, т.к., по-видимому, они преимущественно происходят в тех областях хроматина, где находятся активные гены. В S- период интерфазы процесс репликации каким-то образом, как - неизвестно, проходит через нуклеосомы родительской цепи хроматина, которые переходят на одну из дочерних спиралей ДНК. Тогда все новые октамеры гистонов присоединяются ко второй дочерней спирали ДНК, свободной от нуклеосом.
Нуклеосомная структура сохраняется и во время транскрипции ДНК, хотя довольно трудно представить себе, как РНК-полимераза может транскрибировать ассоциированную с гистонами ДНК без каких-либо заметных изменений в организации нуклеосомы. Но в клетках эмбрионов насекомых в области активированных генов для р-РНК, по-видимому, нуклеосомы отсутствуют. И биохимические отличия между транскрибируемым активным и неактивным хроматином обнаружены. В частности, H1 соединен с нуклеосомами гораздо менее прочно в активном хроматине, и вообще гистоны в этих участках обнаруживают более высокую степень ацетилирования.
15. Особенности строения интерфазных хромосом. Их функция. Понятие о хроматине, виды хроматина. Половой хроматин.
Продольная организация хромосом высших организмов, в основе которой лежит взаимосвязь морфологических, химических и функциональных закономерностей, характеризуется линейной неоднородностью. Уже интерфазные хромосомы оказываются глубоко дифференцированными по степени конденсации хроматина, что первоначально было обнаружено с помощью световой микроскопии.
Одни их участки становятся деконденсированными (эухроматин), другие остаются конденсированными (гетерохроматин). В метафазных хромосомах подразделение на эти два типа хроматина не исчезает. Оно проявляется в естественном ходе митотической конденсации: в ранней профазе районы гетерохроматина опережают в своей конденсации эухроматиновые участки. Остаточные явления неодинаковой конденсации интерфазной хромосомы обнаруживаются морфологически и в метафазе (в области вторичной перетяжки).
Понятие "гетерохроматин" и "эухроматин" в результате цито - генетических исследований получили генетическое содержание. Гетерохроматин в отличие от эухроматина не содержит структурных генов или обеднён ими. В то же время эухроматин - это функционально активный, транскрибируемый хроматин, т.е. структура хроматина оказывает влияние на регуляцию экспрессии эукариотических генов. Подобно митотическому хроматину гетерохроматин не участвует в транскрипции, ДНК в составе гетерохроматина реплицируется в позднем периоде S-периода клеточного цикла. Биохимические основы наблюдаемых различий между гетеро- и эухроматином неизвестны.
Некоторые участки хромосом конденсируются в гетерохроматин во всех клетках организма - это конститутивный гетерохроматин. Другие участки хромосом формируют гетерохроматин лишь в определенных клетках -
факультативный гетерохроматин.
Конститутивный гетерохроматин содержит ДНК, которая, по всей видимости, никогда ни в одной клетке не транскрибируется.
Вхромосомах человека он локализуется вокруг центромер и легко обнаруживается в митотических хромосомах с помощью специальной окраски, хотя может быть обнаружен и в других районах некоторых хромосом (1, 9, 16, У). Подобное же состояние характерно для сателлитной ДНК и ДНК с высокоповторяюшимися последовательностями. Следовательно, большая часть конститутивного гетерохроматина содержит серии сравнительно простых, многократно повторяющихся последовательностей ДНК. В целом функция конститутивного гетерохроматина остается неясной. Предполагают, что некоторые сегменты этого хроматина играют определенную роль в спаривании хромосом в мейозе. Возможно, он влияет на стабилизацию структуры хроматина и защищает генетически значимые последовательности эухроматических районов от внешних воздействий, но классических менделевских генов здесь, скорее всего, нет.
Винтерфазной клетке участки конститутивного хроматина агрегируют с образованием хромоцентров, что мы видим в световой микроскоп в виде мельчайших "глыбок хроматина". У млекопитающих количество их и характер распределения варьируют в зависимости от типа клетки и стадии развития организма.
Факультативный гетерохроматин имеет более отчетливое функциональное значение. Почти не вызывает сомнения, что он отражает устойчивые различия в характере генетической активности клеток разных типов, и количество этого хроматина в разных клетках варьирует: в эмбриональных клетках его совсем немного, тогда как высокоспециализированные клетки содержат его в чрезвычайно больших количествах, т.е. часть генов выключается из транскрипции. Факультативный гетерохроматин содержит уникальные участки ДНК, а не высокоповторяющиеся, и ничем не обнаруживает себя при окрашивании митотических хромосом. Подобный способ генетической регуляции бактериям недоступен.
Частный случай факультативной гетерохроматизации - это инактивация одной из двух Х-хромосом в клетках женских особей млекопитающих, которая происходит на ранних стадиях эмбрионального развития (в трофобласте человека на 12-й день развития, а собственно в эмбрионе -
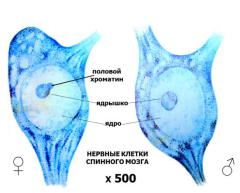
на 16-й день). Одновременно во всех клетках эмбриона женской особи с равной вероятностью одна или другая Х-хромосома конденсируется и образует гетерохроматин. Это состояние хромосомы устойчиво наследуется во всех последующих циклах репликации. Из-за этого каждый женский организм имеет как бы мозаичное строение, т.к. образован клональными группами клеток, примерно в половине которых гетерохроматизирована Х-хромосома, унаследованная по материнской линии, а в другой - Х-хромосома, унаследованная по отцовской линии.
В интерфазе гетерохроматизированные Х- хромосомы представляют собой четко оформленные структурные образования, называемые
тельцами Барра, которые близко прилегают к внутренней мембране ядра и хорошо различимы в световой микроскоп. Тельца Барра
Рисунок 7. Половой хроматин. |
называются |
ещё |
глыбками |
|
полового Х-хроматина (рис.7). |
||
16.Политенные хромосомы. Эндомитоз.
Политенные хромосомы
Чтобы уловить изменения в структуре хроматина на уровне индивидуальных генов, необходимо изучать растянутые интерфазные хромосомы. В обычных клетках это невозможно, потому что нити интерфазного хроматина слишком тонки и запутаны. Благодаря же явлению политении на интерфазных хромосомах отчетливо видны многочисленные поперечные полосы, частота чередования которых позволяет предположить, что они соответствуют индивидуальным генам.
Политенные хромосомы (гигантские хромосомы) содержат во много раз больше ДНК, чем обычные (рис.8, 9). Они не изменяют своей формы на протяжении всего митотического цикла и достигают длины до 0,5 мм, а толщины - до 25 мкм. Они встречаются, например, в слюнных железах
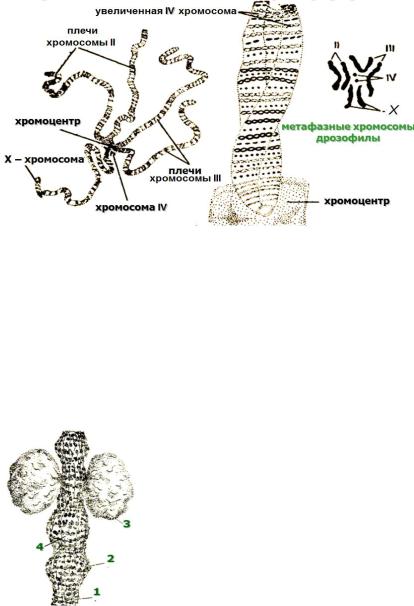
двукрылых (мух, комаров), в макронуклеусе инфузории и в тканях завязи бобов. Чаще всего они видны в гаплоидном числе, т.к. гомологичные хромосомы бывают тесно спарены. Клетки с такими хромосомами вырастают до необычно большого размера.
Возни
кают
полите
нные
хромо
сомы
вследс
твие
много
кратно
повто
ряющ
Рисунок 8. Политенные хромосомы слюнных желез дрозофилы |
егося |
|
проце |
сса редупликации ДНК. При этом разные участки ДНК редуплицируются в разной степени. Большинство генетически информативных областей редуплицируются 1000 раз, а некоторые - более чем 30 тыс. раз. При этом циклы редупликации ДНК не сопровождаются делением клетки.
По существу, политенные хромосомы представляют собой пучки множества неполностью разделенных, тесно прилежащих друг к другу индивидуальных хроматиновых нитей. В частности, политенные хромосомы слюнных желез личинки Drosophila содержат 1024 таких нити.
Итак,
Рисунок 9. Политенные хромосомы: 1 – нити ДНП; 2 – диски;
3– пуфы;
4– интерхромомерные участки ДНП.
интерфазные политенные хромосомы хорошо видны в световой микроскоп, хроматиновые петли в них располагаются в линейном
порядке, при окрашивании этих хромосом заметны перемежающиеся поперечные полосы: темные - диски и светлые - междисковые участки. Предполагают, что именно диски содержат 1024 плотно уложенных гомологичных петли индивидуального петельного участка и расположенные там гены. Структурная организация и функция ДНК междисковых участков пока что неизвестны.
С началом транскрипции генов диски, в которых они содержатся, декомпактизуются, становятся как бы вздувшимися и называются пуфами. Образующая их ДНК упакована гораздо менее плотно. По всей видимости, подобные структурные модификации хроматина, когда происходит его частичная деконденсация, и являются первым этапом активации эукариотических генов. Биохимически пуфы содержат меньше гистона HI, много РНК-полимеразы и, как минимум, один общий негистоновый белок.
Возможно, функциональная единица генома и у высших кариот, в том числе у человека, устроена и функционирует так же.
Хромосомы типа ламповых щеток
Еще одним примером клеток, в которых хорошо различимы транскрипционно активные хромосомы, являются незрелые яйцеклетки, или ооциты. Усиленный синтез РНК в них сопровождается растяжением длинных хроматиновых петель, к которым присоединены многочисленные новообразованные транскрипты, упакованные в РНК-
комплексы. Эти так называемые хромосомы типа ламповых щеток
хорошо видны в световой микроскоп, хотя они не очень конденсированы. Хромосомы типа ламповых щеток (рис. 10) появляются во время диплонемы мейоза при образовании половых клеток у большинства позвоночных, беспозвоночных и зеленых водорослей. Содержание ДНК в таких хромосомах соответствует норме, они не политенны (каждая хромосома содержит две молекулы ДНК).
В хромосомах типа ламповых щеток, помимо петлеобразной укладки суперспирали в виде ерша, имеются отдельные значительно вытянутые симметричные петли, выступающие над поверхностью основной структуры хромосомной укладки.
Рисунок 10. Хромосомы типа ламповой щетки.
хромосомы типа ламповых щеток, по-видимому, создают
запас РНК для последующих стадий развития. Наблюдаемые структуры типа ламповых щеток представляют собой транскрипционно активный хроматин и не являются типичными для соматических клеток.
Эндомитоз-внутриядерное увеличение набора хромосом
Обычно во время клеточного деления РНК не синтезируется, а
17. Особенности строения метафазных хромосом. Типы хромосом. Хромосомный набор. Правила хромосом.
Хромосомы как отдельные структуры становятся доступными для исследования только после значительной конденсации хроматина, которая наступает во время митоза (в соматических клетках) либо во время мейоза (при образовании половых клеток). Начавшаяся в профазе конденсация хроматина заканчивается в метафазе, поэтому, как правило, хромосомы изучаются на стадии метафазной пластинки.
Винтерфазе хромосомы находятся в деконденсированном состоянии, и определить их как отдельные структуры не представляется возможным.
Вметафазе каждая хромосома имеет как бы иксобразную форму и состоит из двух идентичных половин - хроматид (сестринских хромосом), тесно прилежащих друг к другу только в области первичной перетяжки (центромеры), а на остальном протяжении между хроматидами видна большая щель. Центромера - это тот участок, где хромосома находится в деконденсированном состоянии, и к ней прикрепляются нити веретена деления. Центромера делит хромосомы на плечи (рис. 1). По положению центромеры выделяют три вида хромосом.
1.Метацентрические, у которых плечи имеют примерно одинаковую длину (т.е. центромера расположена посередине хромосомы).
2.Субметацентрические, у которых центромера смещена от середины, располагается субмедиально и делит хромосому на два плеча неравной длины. Верхнее всегда меньшее.
З. Акроцентрические, у которых центромера расположена почти на конце хромосомы, отделяя от длинного плеча очень короткое верхнее плечо.
Рисунок 1. Метафазные хромосомы.
Верхние короткие плечи принято обозначать буквой "р", а нижние длинные - буквой "q". Характерной чертой для некоторых хромосом является наличие вторичных перетяжек, они возникают в участках неполной конденсации хромосом и располагаются в околоцентромерных участках 1-й, 9- й и 16-й хромосом. Вторичные перетяжки имеются также в 13-15 и 21 -22-й хромосомах, однако здесь они занимают удаленное от центромеры положение, отделяя небольшой концевой участок короткого плеча хромосом в виде спутника. Эти хромосомы называют спутничными. В этих хромосомах в
области вторичной перетяжки Рисунок 2. Строение хромосомы. сосредоточены гены, кодирующие
р-РНК, и в прилежащих участках кариоплазмы образуются ядрышки. Поэтому такого рода вторичные перетяжки называют ядрышковыми организаторами (рис.2).
В хромосомных наборах одних людей указанные хромосомы имеют вторичную перетяжку, а в этих же хромосомах у других людей её может не быть
18. Кариотип человека. Его определение. Кариограмма, принцип составления. Идиограмма, ее содержание.
Все, что изложено выше относительно химического состава и структуры хромосом эукариот, типично и для хромосом человека. Некоторой детализации требует информация, позволяющая идентифицировать с большей степенью точности любую хромосому человека.
1956 год - шведы Тио и Леван, англичане Форд и Хамертон установили, что ядро диплоидной клетки человека содержит 46 хромосом – это хромосомный набор или кариотип человека; в 1960 – Мурхед и сотр. (США) разработали метод приготовления препаратов хромосом из кратковременной культуры лимфоцитов; в 1968-70 гг. разработаны методы дифференциального окрашивания хромосом, что позволило однозначно идентифицировать все хромосомы человека - все эти манипуляции производились и производятся только на метафазных хромосомах, ибо они различимы лучше всего, т.к. они максимально укорочены и утолщены, лежат свободно одна от другой, располагаются все в одной плоскости клетки (экваториальной); кроме того, исследуются только те метафазные хромосомы, хроматиды которых отделились друг от друга в области плеч, а в центромерной части еще соединены.
Совокупность всех метафазных хромосом, расположенных относительно произвольно в экваториальной плоскости клетки, именуется метафазной пластинкой или просто хромосомным набором. После приготовления препаратов хромосом, которые можно приготовить из всех тканей и клеточных суспензий, содержащих делящиеся клетки (в зависимости от целей важно, конечно, количество метафаз), хромосомы окрашиваются, ибо только после этого их можно различить в световой микроскоп, получить микрофотографию, идентифицировать и, расположив их в определенном порядке, т.е. составив кариограмму, получить целостное представление о кариотипе конкретного человека. Кариограмма - это те же хромосомы метафазной пластинки, но расположенные упорядоченно. Принцип упорядоченности - общий для всего вида и определяется идиограммой. Идиограмма - это графическое изображение гаплоидного набора хромосом (можно и диплоидного) и расположения их по группам в зависимости от формы и величины. Группы располагаются в порядке уменьшения величины входящих в них хромосом.
В современных цитогенетических лабораториях процесс составления кариограммы компьютеризирован.
19.Денверская классификация хромосом и их Парижская номенклатура.
Наиболее простой способ окрашивания хромосом - красителем Гимза или 2%-м ацетоорсеином, или 2%-м ацетокармином. При этом хромосомы окрашиваются целиком, равномерно и интенсивно. Окрашенные таким образом хромосомы, согласно Денверской классификации (I960), располагались в идиограмме в зависимости от их длины и нумеровались по парам от 1 до 23. Тогда же Патау предложил разбить 23 пары
хромосо м на 7
групп от А до G с
учетом
располо
Рисунок 11. Д
жения
центром
еры
(рис.11).
Важным
признако м, уточняю щим форму хромосо
мы, стал центромерный индекс: отношение длины короткого плеча к длине всей хромосомы, выраженное в %. Комплекс этих параметров позволял с немалой степенью точности распределить хромосомы по группам, но идентифицировать их, особенно в группах В, С, D, F и G, было невозможно.
Однако уже при стандартном (рутинном) равномерном окрашивании хромосом замечали, но оставили без внимания некоторую неоднородность в плотности окрашивания по длине хромосом. И только позже (1968 г.), когда Касперсон с сотрудниками обнаружили, что после обработки акрихин-ипритом флуоресценция по длине хромосом распределена не равномерно, а в виде сегментов, они показали, что каждую хромосому можно надежно идентифицировать с помощью такого метода дифференциального окрашивания, ибо расположение сегментов для каждой хромосомы строго специфично. Вскоре стало ясно, что очень
сходный рисунок сегментации хромосом можно получить и с помощью красителя Гимза, дополнив окрашивание некоторыми приемами. Впоследствии при разных способах обработки хромосом были обнаружены разные типы сегментов.
На Парижской конференции по стандартизации и номенклатуре хромосом человека (1971) все полученные к тому времени данные по дифференциальному окрашиванию хромосом были сопоставлены, и оказалось, что все методы в принципе выявляют одни и те же структуры, но каждый специфичен в отношении определенных сегментов (рис. 12). И обозначать различные типы сегментов решили по методам, с помощью которых
они выявляются.
Q - сегменты - флуоресцирующие после окраски акрихин-ипритом; G - сегменты (Гимза) - выявляются при окрашивании красителем Гимза в сочетании с дополнительными процедурами; Q и G сегменты идентичны, но в большинстве лабораторий предпочитают этот метод, т.к. он
не требует использования флуоресцентного микроскопа, и эти препараты дольше хранятся; однако только с помощью Q-метода можно идентифицировать Y-хромосому человека даже в интерфазном ядре;
R - сегменты - окрашиваются после контролируемой тепловой денатурации, располагаются между Q и G - сегментами;
С - сегменты - конститутивный гетерохроматин, располагается в прицентромерных районах обоих плеч хромосомы; Т - сегменты - расположены в теломерных (концевых) районах хромосом.
Химическая природа дифференциального окрашивания еще только исследуется. Обсуждаются две основные гипотезы: первая исходит из того, что различные участки хромосом человека отличаются по количественному содержанию пар оснований аденин - тимин и гуанин - цитозин. Отсюда разная степень усвоения ими красителей. В частности, блоки с большим содержанием пар А-Т связываются преимущественно с
акрихин-ипритом, следовательно, Q-сегменты соответствуют участкам, богатым А-Т - парами; R-сегменты соответствуют участкам, богатым Г-Ц - парами, которые более устойчивы к тепловой денатурации - это, однако, не объясняет всех особенностей сегментации хромосом. Вторая гипотеза, белковая, исходит из данных о том, что предварительная протеолитическая обработка перед окрашиванием красителем Гимза индуцирует появление G-сегментов, а так как разные по составу участки ДНК связаны с разными белками, можно полагать, что рисунок сегментации зависит от особенностей комплекса ДНК - белок.
И все же, что собой представляют полосы - сегменты митотических хромосом, остается загадкой. Даже небольшие тонкие полосы содержат не менее 30 гигантских петель, суммарный нуклеотидный состав которых более 1 млн. нуклеотидов. Возможно, существование таких структурных блоков связано с функционированием эукариотического генома вообще, хотя сами по себе сегменты ничего конкретного о функционировании индивидуальных генов не говорят, ибо в самой тонкой полосе, которую еще можно различить, содержится от 10 до 100 генов. Но то, что картина распределения сегментов в хромосомах почти не изменилась за долгие периоды эволюции (почти каждая хромосома человека имеет своего аналога в кариотипе шимпанзе, гориллы, орангутана), свидетельствует о большом значении пространственной организации ДНК для экспрессии соответствующих генов.
Итак, информация, полученная в результате анализа дифференциально окрашенных хромосом, позволяет представить идиограмму хромосом человека следующим образом:
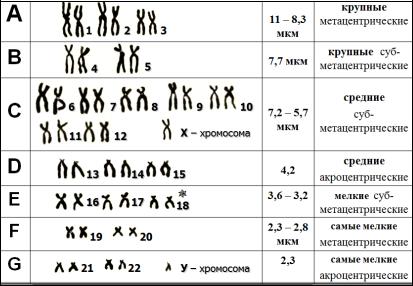
Группа А, 1-3 хромосомы - большие метацентрические и субметацентрические хромосомы; 1-ая - самая большая метацентрическая, центромерный индекс (ЦИ) 48 - 49%, в длинном плече вблизи центромеры часто обнаруживается вторичная перетяжка; вторая самая большая субметацентрическая ЦИ 38-40%; 3-я -почти на 20% короче 1 -ой, ЦИ 45-46%, метацентрическая.
Группа В, 4 и 5 хромосомы - большие субметацентрические. ЦИ 24-30%, без дифференциального окрашивания друг от друга не отличаются.
Группа С, 6-12 хромосомы и Х-хромосома - средние субметацентрические хромосомы 6, 7, 8,11 и 12 - относительно субметацентрические, ЦИ - 27-35; 11 и 12 обнаруживают очень сходный рисунок сегментации, однако 11-я хромосома более метацентрическая; в 9-й в длинном плече часто обнаруживают вторичную перетяжку, которая не окрашивается ни акрихином, ни красителем Гимза; Х-хромосома значительно варьирует по длине, в целом сходна с самыми длинными из
С-группы, ЦИ - 40,12+2,12, отличить от других при стандартном окрашивании очень трудно.
Группа D, 13-15 хромосомы - акроцентрические, ЦИ около 15 - наименьший в кариотипе человека, все они могут иметь вторичную перетяжку на коротком плече или не иметь, а следовательно, иметь спутники или не иметь, спутники могут быть очень большими, а иногда двойными; короткие плечи этих хромосом содержат ядрышковый организатор.
Группа Е, 16-18 хромосомы - относительно короткие метацентрические и субметацентрические; 16 - ЦИ - около 40, длина вариабельна, в длинном плече в 10% случаев выявляется вторичная перетяжка; 17-я, ЦИ -31; 18-ая хромосома на 5 -10%короче17,ЦИ-26.
Группа F, 19, 20 хромосомы - мелкие метацентрические, ЦИ - 36-46, при стандартной окраске выглядят одинаково, при дифференциальной - резко отличаются.
Группа G, 21, 22, Y-хромосомы - мелкие акроцентрические, ЦИ - 13-33; 21 и 22-ая могут иметь спутники, короткие плечи имеют ядрышковый организатор; Y-хромосома обычно (но не всегда) больше, хроматиды ее длинного плеча, как правило, лежат параллельно одна другой, а у 21 и 22
-ой хромосомы они чаще образуют широкий угол; спутники в Y- хромосоме отсутствуют, ЦИ от 0 до 26. В интерфазных ядрах дистальный участок длинного плеча при окрашивании акрихин-ипритом сильно флуоресцирует и выявляется как яркое пятно, которое называется Y-
хроматин.
В соответствии с Парижской номенклатурой в хромосомах идиограммы показан рисунок сегментации (G - Q , R - сегменты) - позитивные светлые G (они же Q) сегменты, негативные темные - R, районы с варьирующей окраской заштриховываются. Латинскими буквами р и q обозначаются соответственно короткое и длинное плечи, в каждом плече выделяются районы, обозначенные арабскими цифрами, районы нумеруются от центромеры к теломерным участкам хромосомы. А уже внутри района выделяются сегменты (англ. bands), обозначенные арабскими цифрами по такому же принципу, т.е. сегмент имеет свой символ, например, 1 q 32
-второй сегмент третьего района в длинном плече 1 -ой хромосомы (при чтении справа налево).
Если в заключение сформулировать интегральную модель хромосомы, то она состоит из единственной двойной спирали ДНК, объединенной с гистонами в нуклеосомы. Некоторые районы этой двойной спирали представлены повторяющимися последовательностями, которые могут
быть рассеяны по всему геному. Участки с повторяющимися
последовательностями обнаруживают признаки конститутивного гетерохроматина. Участки с уникальными последовательностями пар нуклеотидов проявляют свойства эухроматина, это транскрибирующиеся участки - т.е. собственно гены, они соответствуют светлым G- и темным R- сегментам дифференциально окрашенных хромосом.
Благодаря успехам в молекулярной генетике человека разработан принципиально новый метод изучения хромосом – метод флюоресцентной гибридизации in situ (FISH) (in situ – в месте нахождения).
Суть этого молекулярно-цитогенетического метода заключается в следующем:
1 – для изучаемой хромосомы или ее конкретного участка готовят комплементарный однонитевой участок ДНК, к которому присоединяют биотин или дигоксигенин, - такой помеченный участок ДНК называется
зондом;
2 – на микроскопическом препарате хромосом (in situ) при обработке щелочью хромосомная ДНК денатурирует, т.е. разрываются водородные связи между двумя комплементарными нитями ДНК; 3 – полученным ранее зондом обрабатывают препарат – зонд
присоединяется к хромосоме в комплементарном участке ДНК, происходит ренатурация – между зондом и соответствующим участком молекулы ДНК формируются водородные связи; 4 – затем препарат обрабатывают веществом, которое избирательно
может присоединяться к биотину или дигоксигенину, после чего к зонду можно присоединить флюоресцентный краситель (или красный – родамин, или зеленый – флюоресцеина изотиоцианат); 5 – теперь с помощью люминесцентного микроскопа можно увидеть окрашенные хромосомы на фоне неокрашенных.
Можно использовать не только двух, но и трехцветные варианты метода.
Метод FISH применяется очень широко – от определения локализации гена до расшифровки сложных перестроек между несколькими хромосомами. Он требует меньше времени, чем кариотипирование дифференциально окрашенных хромосом.
Метод FISH можно применять для диагностики анеуплоидий в интерфазных ядрах – интерфазная цитогенетика. Например, в течение нескольких часов можно получить информацию о количестве 21 - й хромосомы в клетках амниотической жидкости (пренатальная – дородовая диагностика синдрома Дауна у плода) – специфический ДНК – зонд для 21 - ой хромосомы покажет в ядрах этих клеток или 2 - е
светящиеся точки, что соответствует двум 21-м хромосомам, или три – что выявит трисомию по 21-ой хромосоме
20.Биосинтез белка. Транскрипция, процессинг, трансляция.
Биосинтез белка протекает во все клетки, особенно хорошо идет в молодых клетках.
биосинтез белка протекает в двух стадиях: транскрипции и трансляции.
Транскрипции-это переписывание информации с ДНК матрицы на информационную РНК.
ДНК состоит из генов, каждый ген несет информацию о синтезе одного специфического белка. Все нуклеотиды в ДНК соединяясь по 3, образуют триплеты или кадоны, которые кодируют молекулы ДНК. Во время стадии транскрипции ДНК происходит разрыв водородных связей между комплементарными азотистыми связями, молекулы ДНК раскручиваются. На участке гена ферментативным путем начинает строиться информационная ДНК.
Трансляция является важной составной частью общего метаболизма клетки и ее сущность заключается в переводе генетической информации с мРНК, являющейся первичным продуктом действия генов, в аминокислотную последовательность белков. Трансляция происходит в цитоплазме на рибосомах и является центральным процессом в синтезе белков, в котором помимо рибосом участвуют мРНК, 3-5 молекул рРНК, 40-60 молекул разных тРНК, аминокислоты, около 20 ферментов (аминоацил-тРНК синтетаз), активирующих аминокислоты, растворимые белки, вовлекаемые в инициацию, элонгацию и терминацию полипептидной цепи.
Трансляция:
1.Активация аминокислот. 2.Инициация, начало синтеза белка. 3.элонгация, удлинение. 4.терминация, окончание синтеза.
Все участвующие в процессе синтеза белка аминокислоты находятся в цитоплазме: метионин, лейцин, серин, лизин.
Процессинг – это процесс образования зрелых молекул рибонуклеиновой кислоты из пре-РНК. Иначе говоря, это совокупность реакций, которые приводят к трансформации первичных продуктов транскрипции (пре-РНК разных типов) в уже функционирующие молекулы.
Что касается процессинга р- и тРНК, он чаще всего сводится к отсечению с концов молекул лишних фрагментов. Если говорить об иРНК, то здесь
можно отметить, что у эукариот данный процесс протекает многоступенчато.
21.ДНК. Строение, свойства, кодовая система.
Дезоксирибонуклеи́новая кислота́(ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК содержит информацию о структуре различных видов РНК и белков.
Вклетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одноили двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих … вирусов.
Строение С химической точки зрения ДНК — это длинная полимерная молекула,
состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы (фосфодиэфирные связи). В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула спирализована. В целом структура молекулы ДНК получила название «двойной спирали».
Кодовая система
ВДНК встречается четыре вида азотистых оснований (аденин, гуанин, тимин и цитозин). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин соединяется только с тимином, гуанин — только с цитозином. Последовательность нуклеотидов позволяет «кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК
синтезируются на матрице ДНК за счёт копирования последовательности ДНК в последовательность РНК, синтезируемой в процессе транскрипции, и принимают участие в биосинтезе белков (процессе трансляции).
Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции. Кроме того, в геноме эукариот часто встречаются участки, принадлежащие «генетическим паразитам», например, транспозонам
22.Генетика как наука. Основные понятия генетики: наследственность, изменчивость; аллельные гены, гомо- и гетерозиготы; признаки - доминантные, рецессивные, альтернативные; генотип, фенотип; менделирующие признаки.
ГЕНЕТИКА – наука о наследственности и изменчивости. Наследственность – всеобщее свойство живого в виде способности организмов передавать свои признаки и свойства из поколения в поколение.
Изменчивость – свойство прямо противоположное наследственности – способность организмов приобретать новые признаки и свойства в процессе индивидуального развития организмов (онтогенеза).
1900 год – год рождения генетики как науки.
«Отцом», основоположником генетики считается чешский естествоиспытатель Грегор Мендель Аллель – одна из двух или более альтернативных форм гена, каждая из
которых характеризуется уникальной последовательностью нуклеотидов. Гетерозигота – клетка (или организм), содержащая два различных аллеля в конкретном локусе гомологичных хромосом.
Гомозигота – клетка (или организм), содержащая два одинаковых аллеля в конкретном локусе гомологичных хромосом.
Доминантные признаки- признак, преобладающий над рецессивным у гетерозигот.
Рецессивный признак- признак, проявляющийся только у гомозигот Альтернативный признак-взаимоисключающий признак Генотип – совокупность всех генов организма, определяющих его фенотип.
Фенотип- Совокупность внешних проявлений признаков одного организма Менделирующие признаки- признаки, наследование которых,
подчиняется закону Менделя.
23.Гибридологический метод, его сущность. Виды скрещиваний - моно- и полигибридное, анализирующее. Их сущность.
Для постановки эксперимента при изучении наследования признаков, Г.
Менделем был разработан метод гибридологического анализа. Вот основные его свойства:
1)в скрещивании участвуют организмы, принадлежащие к одному
виду;
2)изучаемые признаки должны быть взаимоисключающими или контрастными (альтернативными);
3)исходные родительские формы должны быть «чистыми линиями» (гомозиготами) по изучаемым признакам;
4)при изучении закономерностей наследования, необходимо начинать работу с анализа минимума количества признаков, постепенно усложняя эксперимент: родительские особи должны отличаться по одной паре альтернативных признаков → двум парам → небольшому числу пар альтернативных признаков;
5)проводить индивидуальный анализ потомства и при наличии расщепления в поколении необходимо проводить статистический анализ;
6)изучение закономерностей наследования проводится на протяжении нескольких поколений.
Таким образом, гибридологический анализ – это система скрещиваний, позволяющая проследить в ряду поколений характер наследования признаков и выявить новообразования.
Моногибидное скрещивание – родительские особи, взятые для скрещивания, отличаются одной парой альтернативных признаков. Дигибридное скрещивание – организмы, взятые для скрещивания, отличаются двумя парами альтернативных признаков.
Закономерности наследования, установленные Г. Менделем при моно- и дигибридном скрещиваниях:
1 закон – «Закон единообразия гибридов первого поколения»;
2 закон – «Закон расщепления»;
3 закон – «Закон независимого наследования (комбинирования) признаков».
1 закон – «Закон единообразия гибридов первого поколения»
При скрещивании родительских особей, отличающихся друг от друга по одной паре альтернативных признаков, все гибриды (потомки) первого

поколения (F1) будут единообразны как по генотипу, так и по фенотипу.
|
|
|
|
желтые |
зеленые |
|
Условные |
P: |
♀ AA |
x |
♂ aa |
||
обозначения: |
|
|
|
|
||
А – желтая окраска |
G: |
A |
|
a |
||
|
|
желтые |
|
|||
семян; |
|
|
|
|||
F1: |
Aa – 100% |
|
||||
|
|
|
|
|||
а – зеленая окраска |
|
гетерозиготы |
||||
Выводы из первого закона Менделя: |
|
|
|
|||
|
|
|
|
|||
|
|
|
по генотипу: 100% все гибриды – гетерозиготы |
|||
|
|
|
||||
|
F1: |
|
(Аа) |
|
|
|
|
|
по фенотипу: 100% все гибриды с желтой |
||||
|
|
|
||||
|
|
|
окраской семян (доминантным признаком) |
|||
|
|
|
|
|
|
|
2 закон – «Закон расщепления»
В ходе дальнейшего скрещивания гибридов первого поколения, во втором поколении (F2) наблюдается расщепление по генотипу 1:2:1, по фенотипу 3:1.
|
|
желтые |
|
желтые |
Условные |
P (F1): |
♀ Aа |
x |
♂ Аa |
обозначения: |
|
|
|
|
А – желтая окраска |
G: |
A, а |
|
А, a |
|
|
|
|
|
семян; |
|
желтые |
|
зеленые |
|
|
|
||
а – зеленая окраска |
|
|
|
|
|
F2: |
АА, Aa, |
Aa, |
аa |
|
|
25% |
50% |
25% |
Выводы из второго закона Менделя:
по генотипу: 1 : 2 : 1
25% – доминантные гомозиготы (АА);
50% – гетерозиготы (Аа);

F2:
3 закон – «Закон независимого наследования (комбинирования) признаков»
При скрещивании родительских особей, отличающихся друг от друга по двум парам альтернативных признаков, во втором поколении (F2) наблюдается независимое наследование признаков в соотношении по фенотипу 9:3:3:1 по двум парам признаков или (3:1)2 по каждой паре признаков.
Условные обозначения:
1-ая пара признаков – окрас-ка семян:
А – желтая; а – зеленая.
2-ая пара признаков – форма семян:
В – гладкая;
b – морщинистая.
|
жел. ∕ гл. |
зел. ∕ мор. |
P: |
♀ AAВВ |
x ♂ aabb |
G: |
AВ |
ab |
|
жел. ∕ гл. |
|
F1: |
AaВb – 100% |
|
|
дигетерозиготы |
|
Продолжаем скрещивать гибридов первого поколения (F1).
|
жел. ∕ гл. |
|
жел. ∕ гл. |
P (F1): |
♀ AаВb |
x |
♂ АaВb |
G: |
AB |
АB |
|
Аb |
Аb |
|
аB |
аB |
|
аb |
аb |
|
F2: |
см. решетку Пеннета |
|
|
|
|
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
AB |
|
Ab |
|
aB |
|
Ab |
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
AB |
|
жел. ∕гл. |
|
жел. ∕гл. |
|
жел .∕гл. |
|
жел. ∕гл. |
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
AABB |
|
|
AABb |
|
|
AaBB |
|
|
AaBb |
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
Ab |
|
жел. ∕гл. |
|
жел. ∕ мор. |
|
жел. ∕ гл. |
|
жел. ∕ мор. |
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
ААВb |
|
|
ААbb |
|
|
АаВb |
|
|
Ааbb |
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
аB |
|
жел. ∕ гл. |
|
жел. ∕гл. |
|
зел. ∕ гл. |
|
зел. ∕ гл. |
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
АаВВ |
|
|
АаВb |
|
|
ааВВ |
|
|
ааВb |
|
||||||||||
|
|
|
|
|
|
|
|
|
||||||||||||||
аb |
|
жел. ∕гл. |
|
жел. ∕ мор. |
|
зел. ∕ гл. |
|
зел. ∕ мор. |
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
АаВb |
|
|
Ааbb |
|
|
ааВb |
|
|
ааbb |
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Выводы из третьего закона Менделя:
Если проводить анализ по каждой паре признаков, то из 16 гибридов второго поколения (F2) желтую окраску семян имеют 12 потомков, а 4 – зеленую. Таким образом, соотношение по окраске семян будет 3:1. Тоже самое наблюдается при наследовании формы семян 12:4 или 3:1. Следовательно, соотношение по фенотипу при анализе по каждой паре признаков соответствует (3:1)2.
Необходимо помнить, что третий закон Менделя выполним при наличии
следующих условий:
1) полное доминирование (гетерозигота фенотипически выглядит так же, как и доминантная гомозигота); расположение генов, отвечающих за разные признаки (окраску и форму
семян), в разных парах гомологичных хромосом. Только так у дигеторозиготы образуется 4 сорта половых клеток в процессе гаметогенеза. Анализирующее скрещивание проводится с целью
установления генотипа исследуемой особи. Для этого исследуемую особь
(?) скрещивают с рецессивной гомозиготой (аа).
Если в F1 наблюдается расщепление 1:1, то исследуемая особь является по генотипу гетерозиготой – Аа.
P: ♀ (?) |
Х |
♂ аа |
|
P: ♀ Aa |
Х |
♂ аа |
||||
G: |
? |
|
a |
|
G: |
A, a |
|
a |
||
F1: |
Aa; |
aa |
– 1:1 |
выяснение |
F1: |
Aa; |
aa |
– 1:1 |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
генотипа |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Если в F1 наблюдается единообразие, то исследуемая особь является по генотипу гомозиготой – АА или аа.
|
|
|
|
|
P: ♀ AA |
Х |
♂ аа |
|
P: ♀ |
(?) |
Х |
♂ аа |
|
||||
|
G: A |
|
a |
|||||
G: |
? |
|
a |
выяснение |
|
|||
|
F1: Aа |
– |
100 % |
|||||
F1: |
Aа |
– |
100 % |
|||||
генотипа |
||||||||
|
|
|
||||||
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ОПЫТЫ ТОМАСА МОРГАНА
2 пары анализируемых |
|
|
сер./норм. |
чер./зач. |
признаков: |
|
P: ♀ ААBB |
Х ♂ ааbb |
|
А – серое тело; |
|
G: |
АB |
аb |
а – черное тело; |
|
|||
|
|
|
|
|
В – нормальные крылья; |
|
F1: |
АаBb – |
100 % |
|
серые с нормальными крыльями |
|||
b – зачаточные крылья. |
|
|||
|
|
|
|
|
|
|
|
|
|
При проведении анализирующего скрещивания самки (F1) из первого поколения с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% – серые длиннокрылые и 41,5% – черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% – черные длиннокрылые и 8,5% – серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены,

отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов – АВ и аb, а отцовский – один тип – аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип АаВb и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb.
Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток – мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
АНАЛИЗИРУЮЩЕЕ СКРЕЩИВАНИЕ
ОПЫТ №1. ПОЛНОЕ СЦЕПЛЕНИЕ
(из F1 берется самец – АаBb)
P: |
♀ ааbb |
Х ♂ (F1) АаBb |
G: |
аb |
? |
F2: АаBb – 50% – серые с нормальными крыльями;
ааbb – 50% – черные с зачаточными крыльями.
Следовательно, дигетерозиготный самец (АаBb) дает два сорта гамет:
●АB;
●аb
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза), или в разных (транс-фаза) (рис. 9).
Рис. 9. 1 – механизм цис-фазы (некроссоверные гаметы); 2 – механизм транс-фазы (некроссоверные гаметы).
25. Гипотеза чистоты гамет, ее цитологическое обоснование.
Г. Менделю не было известно, где находятся открытые им "наследственные факторы" (гены) и что они собой представляют. В 1910 г. американский биолог Томас Хант Морган (1866-1945) и его последователи создали и обосновали хромосомную теорию наследственности. Согласно этой теории, гены расположены в хромосомах, в клеточном ядре. Научная школа Моргана выяснила цитологические основы законов Менделя.
Вкаждой клетке взрослого организма (за исключением половых клеток) имеется двойной набор хромосом. Половина этих хромосом получена от отца, половина – от матери. В половые клетки попадает только одинарный набор хромосом. Это происходит при мейозе.
Цитологические основы выполнения закона расщепления заключаются в том, что в процессе мейоза во время гаметогенеза гомологичные хромосомы, несущие гены исследуемого признака, расходятся в разные половые клетки, унося с собой либо отцовский аллель, либо материнский.
Висходной диплоидной материнской клетке (2n) в первом делении мейоза в дочерние клетки расходятся пары гомологичных хромосом, а во втором – хроматиды. В результате мейоза образуются гаплоидные клетки, несущие по одной копии гена каждой хромосомы.
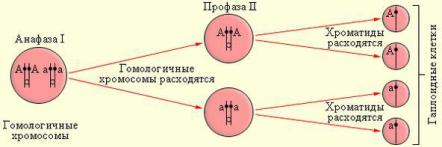
Рис 5. Первый закон Менделя на языке хромосом.
В конце мейоза образуются четыре клетки, две из которых несут отцовскую копию хромосомы, а две – материнскую (рис. 5). Значит, каждая гамета несёт только одну копию данного гена, то есть может содержать лишь один аллель данного гена. Это положение получило название принципа чистоты гамет. Таким образом, цитологической основой закона расщепления является мейоз, гаплоидность и принцип чистоты гамет, а также случайное объединение наследственного материала в зиготе при оплодотворении.
Сам Г. Мендель на основе анализа своих скрещиваний выдвинул гипотезу о том, что рецессивные задатки не исчезают в гетерозиготном организме, а остаются неизменными и вновь проявляются при встрече с такими же рецессивными задатками в последующих поколениях или в анализирующих скрещиваниях.
Позднее У. Бетсон, исходя из этого феномена, сформулировал правило «чистоты гамет», согласно которому наследственные задатки не смешиваются в гетерозиготном организме и расходятся "чистыми" при образовании гамет (в гамету попадает по одному фактору наследственности (аллелю) каждого типа).
Таким образом, у гибридов (гетерозиготных особей) примерно 50% половых клеток несут только доминантный ген (А), остальные – только рецессивный (а). Встреча и слияние двух половых клеток происходит на основании случайности. Каким же окажется потомство двух таких гибридов? Очевидно, что по теории вероятности половина потомства окажется гетерозиготной (Аа и Аа), а половина – гомозиготной (АА и аа). Но только четверть потомства, несущая гены аа, проявит рецессивный признак. Отсюда и открытое Менделем соотношение "три к одному"
26. Закон Менделя, основанный на дигибридном скрещивании.
Эксперимент расписать.
3 закон – «Закон независимого наследования
(комбинирования) признаков»
При скрещивании родительских особей, отличающихся друг от друга по двум парам альтернативных признаков, во втором поколении (F2) наблюдается независимое наследование признаков в соотношении по фенотипу 9:3:3:1 по двум парам признаков или (3:1)2 по каждой паре признаков.
Условные обозначения:
1-ая пара признаков – окрас-ка семян:
А – желтая; а – зеленая.
2-ая пара признаков – форма семян:
В – гладкая;
b – морщинистая.
|
жел. ∕ гл. |
зел. ∕ мор. |
P: |
♀ AAВВ |
x ♂ aabb |
G: |
AВ |
ab |
|
жел. ∕ гл. |
|
F1: |
AaВb – 100% |
|
|
дигетерозиготы |
|
Продолжаем скрещивать гибридов первого поколения (F1).
|
жел. ∕ гл. |
|
жел. ∕ гл. |
P (F1): |
♀ AаВb |
x |
♂ АaВb |
G: |
AB |
|
АB |
|
Аb |
|
Аb |
|
аB |
|
аB |
|
аb |
|
аb |
F2: |
см. решетку Пеннета |
||
|
AB |
Ab |
aB |
Ab |
|
|
|
|
|

|
|
AB |
|
жел. ∕гл. |
жел. ∕гл. |
|
жел .∕гл. |
жел. ∕гл. |
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
AABB |
|
|
AABb |
|
|
|
AaBB |
|
|
AaBb |
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
Ab |
|
жел. ∕гл. |
жел. ∕ мор. |
|
жел. ∕ гл. |
жел. ∕ мор. |
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
ААВb |
|
|
ААbb |
|
|
|
АаВb |
|
|
Ааbb |
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
аB |
|
жел. ∕ гл. |
жел. ∕гл. |
|
зел. ∕ гл. |
зел. ∕ гл. |
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
АаВВ |
|
|
АаВb |
|
|
|
ааВВ |
|
|
ааВb |
|
|
||||||
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
аb |
|
жел. ∕гл. |
жел. ∕ мор. |
|
зел. ∕ гл. |
зел. ∕ мор. |
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
АаВb |
|
|
Ааbb |
|
|
|
ааВb |
|
|
ааbb |
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Выводы из третьего закона Менделя: |
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
по фенотипу: |
9 : 3 : 3 : 1 |
(по двум парам |
||||||||||||
|
|
|
|
|
|
|
|
|
признаков): |
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
9∕ 16 |
– желтые и гладкие (А•В•); |
||||||||||||
|
|
F2: |
|
|
|
|
|
|
|
3∕ 16 |
– желтые и морщинистые (А•bb); |
||||||||||||
|
|
|
|
|
|
|
|
|
|
3∕ 16 |
– зеленые и гладкие (ааВ•); |
||||||||||||
|
|
|
|
|
|
|
|
|
|
1∕ 16 |
– зеленые и морщинистые (ааbb). |
||||||||||||
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Если проводить анализ по каждой паре признаков, то из 16 гибридов второго поколения (F2) желтую окраску семян имеют 12 потомков, а 4 – зеленую. Таким образом, соотношение по окраске семян будет 3:1. Тоже самое наблюдается при наследовании формы семян 12:4 или 3:1. Следовательно, соотношение по фенотипу при анализе по каждой паре признаков соответствует (3:1)2.
Необходимо помнить, что третий закон Менделя выполним при наличии
следующих условий:
2)полное доминирование (гетерозигота фенотипически выглядит так же, как и доминантная гомозигота);
3)расположение генов, отвечающих за разные признаки (окраску и форму семян), в разных парах гомологичных хромосом. Только так у дигеторозиготы образуется 4 сорта половых клеток в процессе гаметогенеза (рис. 4).

А |
|
а |
|
В |
|
|
|
b |
|||
• |
|
• |
АаВb |
||
|
|
• |
• |
||
|
|
|
|
|
|
|
|
|
|
|
|
дигетерозигота дает 4 сорта гамет
Рис 4. Гаметы дигетерозиготы
27.Хромосомный механизм детерминации признаков пола.
Начало изучению генотипического определения пола было положено открытием американскими цитологами у насекомых различия в форме, а
иногда |
и в числе хромосом у особей разного пола (Мак-Кланг, 1906, |
|||||
Уилсон, |
1906) |
и |
классическими |
опытами |
немецкого |
генетика |
К. Корренса |
по |
скрещиванию однодомного и двудомного видов |
||||
брионии. |
|
|
|
|
|
|
Уилсон обнаружил, что у клопа Lydaeus turucus самки имеют 14 хромосом (7 пар), как и самцы. Из них 6 пар – одинаковые у обоих полов, а в седьмой паре у самки – две одинаковые хромосомы, а у самца одна хромосома такая же, как соответствующая хромосома самки, а другая маленькая.
Пара хромосом, которые у самца и самки разные, получила название
гетерохромосомы, или половые хромосомы. У самки две одинаковые половые хромосомы, обозначаемые как Х-хромосомы, у самца одна Х- хромосома, другая – Y-хромосома. Остальные хромосомы, одинаковые у самца и у самки, были названы аутосомами.
Таким образом, хромосомная формула у самки названного клопа – 12A + XX, у самца – 12A + XY. В процессе гаметогенеза самка клопа образует один сорт гамет – 7A + X, а самец 2 сорта гамет – 7A + Y или 7A + X. Пол организма с генотипом ХХ называют гомогаметным, так как у него образуются одинаковые гаметы, содержащие только Х-хромосомы, а пол с генотипом XY – гетерогаметным, так как половина гамет содержит Х-, а половина – Y-хромосому.
Сочетание XX–XY (рис. 17) определяет пол млекопитающих, человека и некоторых насекомых (клопы, мушка-дрозофила). При гаметогенезе наблюдается типичное менделевское расщепление по половым

хромосомам. Каждая яйцеклетка содержит одну Х-хромосому. Сперматозоид содержит одну Y-хромосому или одну Х-хромосому. Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку. При слиянии сперматозоида и яйцеклетки, несущих Х-хромосомы, образуется зигота с двумя Х-хромосомами. В дальнейшем из таких зигот развиваются нормальные самки. Если в зиготу попадают Х- и Y-хромосомы, то в дальнейшем из них развиваются самцы.
Сочетание ZZ–ZW (рис. 17) характерно для птиц, бабочек, шелкопряда, рептилий, земноводных. При слиянии сперматозоида и яйцеклетки, несущих Z-хромосомы, образуется зигота с двумя Z-хромосомами. В дальнейшем из таких зигот развиваются нормальные самцы (гомогаметный пол). Если в зиготу попадают Z- и W-хромосомы, то в дальнейшем из них развиваются самки (гетерогаметный пол). В научной литературе вместо символа Z используется Х, имея в виду, что Z- хромосома соответствует Х-хромосоме.
Рис. 17. Гомо- и гетерогаметный пол. Сочетание XX–XО (рис. 18)
характерно для некоторых
насекомых (кузнечики, моль). Сочетание ХХ характерно для гомогаметного пола, появляется оно при слиянии яйцеклетки и сперматозоида, несущих по Х- хромосоме. Если происходит слияние половой клетки, несущей Х-хромосому, с гаметой, у которой нет ни одной половой хромосомы, образуется сочетание ХО и развиваются организмы гетерогаметного пола.
|
|
|
|
Рис. 18. Сочетание XX–XО. |
||
Совершенно |
другой |
механизм |
|
|||
определения |
|
|
|
пола, |
|
|
называемый |
гаплодиплоидный |
|
||||
– сочетание n – 2n широко |
|
|||||
распространен у пчел и муравьев |
|
|||||
(рис.19). |
|
|
|
|
|
|
У этих |
организмов нет половых |
|
||||
хромосом: |
самки |
|
|
– это |
|
|
диплоидные |
особи, |
а |
самцы |
|
||
(трутни) |
– |
гаплоидные. |
Самки |
|
||
развиваются |
|
|
|
из |
|
|
оплодотворенных |
яиц (2n), а из |
|
||||
неоплодотворенных |
|
путем |
Рис. 19. Гаплодиплоидный |
|||
партеногенеза |
развиваются |
механизм наследования пола. |
||||
трутни (n). |
|
|
|
|
|
|
Балансовая теория определения пола
К. Бриджес в 1919 году обнаружил, что у плодовой мушки дрозофилы на проявление пола оказывает влияние соотношение (баланс) между количеством Х-хромосом и набором аутосом, а Y-хромосома на детерминацию пола влияния не оказывает.
У нормальных самок на две Х-хромосомы (2Х) приходится два набора аутосом (2А). Половой индекс в этом случае равен 1 (2Х:2А). У нормальных самцов только одна Х-хромосома. Половой индекс в этом случае равен 0,5 (1Х:2А).
Рис. 20. Самец (а), самка (б) и некоторые ненормальные половые типы дрозофилы: интерсекс (в), сверхсамка (г), сверхсамец (д).
В случае неравномерного распределения генетического материала в кариотипе может оказаться на одну Х-хромосому больше (ХХХ или ХХY), а количество аутосом останется в норме. При наличии 3Х хромосом половой индекс будет равен 1,5 (3Х:2А) и разовьются сверхсамки. При наличии 2Х хромосом и 1Y хромосомы половой индекс будет равен 1
(2Х:2А) и появятся нормальные самки, так как Y-хромосома в этом случае на пол влияния не оказывает.
При неравномерном распределении генетического материала в кариотипе может оказаться нормальное число половых хромосом, а число наборов аутосом будет кратно увеличено. При кариотипе 2Х+3А половой индекс будет равен 0,67 (2Х:3А) и разовьються интерсексы, а при кариотипе ХY+3А половой индекс будет равен 0,33 (1Х:3А) и разовьются сверхсамцы (бесплодны).
Таким образом, следует, что при половом индексе равном 1 развиваются нормальные самки, 0,5 – нормальные самцы, выше 1 – сверхсамки, ниже 0,5 – сверхсамцы, при половом индексе меньше 1, но выше 0,5 – интерсексы (рис. 20).
Генетический пол в популяциях людей
Половые хромосомы (X и Y) являются единственными из 23 хромосом, которые определяют пол организма. Половые клетки, продуцируемые женским организмом, содержат Х-хромосому, тогда как сперматозоиды – X-или Y-хромосому. Остальные 22 пары хромосом предопределяют характеристики организма, не связанные с полом, и поэтому называются аутосомами.
Во время оплодотворения образуется зигота, содержащая 23 пары хромосом. В зиготе, из которой будет развиваться плод женского пола, содержится две Х-хромосомы, т. е. по одной из каждой женской и мужской гаметы. В зиготе, из которой будет развиваться плод мужского пола, содержится Х- и Y-хромосомы (рис. 21). Развитие гонад и гениталий в мужском организме обусловлено Y-хромосомой. При отсутствии в генах эмбриона Y-хромосомы у эмбриона развиваются яичники. Y-хромосома ответственна за начальный половой диморфизм.
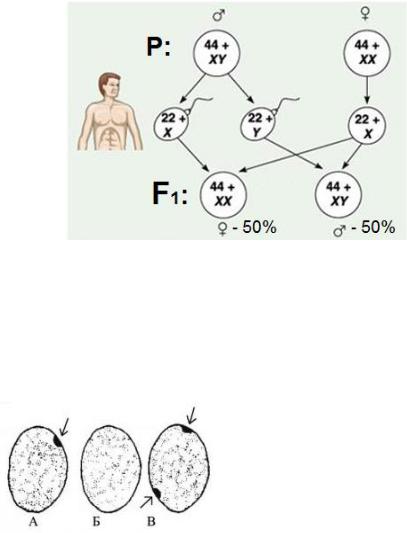
Рис. 21. Сингамное определение пола у человека.
У человека генотипический пол определяют, изучая соматические клетки буккального соскоба. Одна Х-хромосома всегда оказывается в активном состоянии и имеет обычный вид. Другая, если она имеется, бывает в покоящемся состоянии в виде плотного темно-окрашенного тельца, называемого тельцем Барра (факультативный гетерохроматин). Число телец Барра всегда на единицу меньше числа наличных Х-хромосом, т.е. в мужском организме (ХУ) в норме их нет вовсе, у женщин (ХХ) – одно (рис.
22).
эпителиальная соматическая клетка женщины (ХХ); Б – соматическая клетка мужчины (ХУ); В – соматическая клетка индивида с тремя Х- хромосомами (ХХХ).
Рис. 22. Тельце Барра или половой хроматин: А –
У человека Y-хромосома является генетически инертной, так как в ней очень мало генов. Однако влияние Y-хромосомы на детерминацию пола у человека очень сильное. Хромосомная формула мужчины 44A+XY и женщины 44A+XX такая же, как и у дрозофилы, однако у человека особь с кариотипом 44A+XО оказывается женщиной, а особь 44A+XXY – мужчиной. В обоих случаях они проявляли дефекты развития, но все же пол определялся наличием или отсутствием Y-хромосомы. Организм с генотипом 47, XXX представляют собой бесплодную женщину, с генотипом 48, XXXY – бесплодного умственно отсталого мужчину. Такие
генотипы возникают в результате нерасхождения половых хромосом, что приводит к нарушению развития (например, синдром Клайнфельтера, XXY). Нерасхождение хромосом случается как в мейозе, так и в митозе. Нерасхождение может быть следствием физического сцепления Х-хромосом, в таком случае нерасхождение имеет место в 100% случаев.
Всем млекопитающим мужского пола, включая человека, свойственен так называемый H-Yантиген, находящийся на поверхности клеток, несущих Y-хромосому. Единственной функцией его считается дифференцировка гонад.
Вторичные половые признаки развиваются под влиянием стероидных гормонов, вырабатываемых гонадами. Развитие мужских вторичных половых признаков контролирует тестостерон, воздействующий на все клетки организма, включая клетки гонад. Мутация всего одного гена Х- хромосомы – гена рецептора андрогенов (AR), кодирующего белокрецептор тестостерона, приводит к синдрому тестикулярной феминизации особей XY (Х-сцепленный рецессивный тип наследования). Клетки-мутанты не чувствительны к действию тестостерона, в результате чего взрослый организм приобретает черты, характерные для женского пола (ложный мужской гермафродитизм, мужской псевдогермафродитизм). При этом внутренние половые органы оказываются недоразвитыми и такие особи полностью стерильные (рис. 23). Таким образом, в определении и дифференцировке пола млекопитающих и человека взаимодействуют хромосомный и генный механизмы.
Рис. 23. Типичный внешний вид полной нечувствительности к андрогенам при синдроме тестикулярной феминизации.
Несмотря на то, что женщины имеют две Х-хромосомы, а мужчины – только одну, экспрессия генов Х-хромосомы происходит на одном и том же уровне у обоих полов. Это объясняется тем, что у женщин в каждой клетке полностью инактивирована одна Х-хромосома (тельце Барра). Х- хромосома инактивируется на ранней стадии эмбрионального развития, соответствующей времени имплантации. При этом в разных клетках отцовская и материнская Х-хромосомы выключаются случайно.

Состояние инактивации данной Х-хромосомы наследуется в ряду клеточных делений. Таким образом, женские особи, гетерозиготные по генам половых хромосом, представляют собой организмы-мозаики (пример, черепаховые кошки (рис. 24)).
Рис. 24. ХbХb – рыжая кошка; ХbУ – рыжий кот; ХBХB – черная кошка; ХBУ – черный кот; ХBХb –
черепаховая (полосатая) кошка.
Таким образом, пол человека представляет собой менделирующий признак, наследуемый по принципу обратного (анализирующего) скрещивания. Гетерозиготой оказывается гетерогаметный пол (XY), который скрещивается с рецессивной гомозиготой, представленной гомогаметным полом (XX). В результате в природе обнаруживается наследственная дифференцировка организмов на мужской и женский пол и устойчивое равенство полов –
50% на 50%.
28.Сцепленное наследование, кроссинговер, определение расстояния между генами на эксперименте с дрозофилами. Группы сцепления, карты хромосом.
Г. Мендель изучил наследование только семи пар признаков у душистого горошка. Его законы подтвердились на самых разных видах организмов, т.

е. было признано, что эти законы носят всеобщий характер. Однако позже, в 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Постепенно таких исключений из третьего закона Менделя накапливалось все больше. Стало ясно, что принцип независимого распределения в потомстве и свободного комбинирования распространяется не на все гены.
Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы изучено более 500 генов, у мухи дрозофилы – более 1 тыс., а у человека – около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно.
Это дало основание предположить, что в каждой хромосоме локализовано множество генов.
Экспериментальные исследования явления сцепленного наследования провел выдающийся американский генетик Т. X. Морган со своими сотрудниками. Сцепленным наследованием называется такой вариант наследования признаков, при котором гены, отвечающие за развитие данных признаков, располагаются в одной хромосоме, составляют группу сцепления (рис.7) и наследуются совместно (сцепленно).
|
|
|
|
|
|
|
|
|
|
|
C |
|
Гены A, b, Z располагаются в одной |
|
|
|
|
|
|
хромосоме и составляют одну группу |
А |
|
|
|
D |
сцепления |
|
|
||||||
|
|
|
|
|
||
|
|
|
|
|
|
Гены C, D, f располагаются тоже в |
b |
|
|
|
|
|
|
|
|
|
|
|
одной хромосоме, но составляют |
|
|
|
|
|
|
|
другую группу сцепления |
Z |
|
|
|
f |
||
|
|
|
|
|||
|
|
|
|
|||
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
Рис. 7. Группы сцепления генов в хромосомах. |
||||
Если Г. Мендель проводил свои опыты на горохе, то для Т. Моргана основным объектом стала плодовая мушка дрозофила.
Дрозофила каждые две недели при температуре 25° С дает многочисленное потомство. Самец и самка внешне хорошо различимы – у самца брюшко меньше и темнее (рис. 8). Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках
Рис. 8. Самка и самец плодовой на недорогой питательной среде
мушки дрозофилы.
.
Томас Морган, как в свое время Грегор Мендель в опытах с горохом, изучал наследование 2 пар признаков: окраска тела и длина крыльев у мушки дрозофилы. Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении (F1) Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, – над геном недоразвитых).
Следовательно, в F1 Томас Морган получил единообразие гибридов первого поколения, как и Грегор Мендель, но в F2 Томас Морган никогда не получал соотношения 9:3:3:1 как в третьем законе Менделя. Тогда для выяснения генотипов особей первого поколения (F1) он провел анализирующее скрещивание.
Анализирующее скрещивание проводится с целью установления генотипа исследуемой особи. Для этого исследуемую особь (?) скрещивают с рецессивной гомозиготой (аа).
Если в F1 наблюдается расщепление 1:1, то исследуемая особь является по генотипу гетерозиготой – Аа.
P: ♀ (?) |
Х |
♂ аа |
|
P: ♀ Aa |
Х |
♂ аа |
||||
G: |
? |
|
a |
|
G: |
A, a |
|
a |
||
F1: |
Aa; |
aa |
– 1:1 |
выяснение |
F1: |
Aa; |
aa |
– 1:1 |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
генотипа |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Если в F1 наблюдается единообразие, то исследуемая особь является по генотипу гомозиготой – АА или аа.
|
|
|
|
|
P: ♀ AA |
Х |
♂ аа |
|
P: ♀ |
(?) |
Х |
♂ аа |
|
||||
|
G: A |
|
a |
|||||
G: |
? |
|
a |
выяснение |
|
|||
|
F1: Aа |
– |
100 % |
|||||
F1: |
Aа |
– |
100 % |
|||||
генотипа |
||||||||
|
|
|
||||||
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ОПЫТЫ ТОМАСА МОРГАНА
2 пары анализируемых |
|
|
сер./норм. |
чер./зач. |
признаков: |
|
P: ♀ ААBB |
Х ♂ ааbb |
|
А – серое тело; |
|
G: |
АB |
аb |
а – черное тело; |
|
|||
|
|
|
|
|
В – нормальные крылья; |
|
F1: |
АаBb – |
100 % |
|
серые с нормальными крыльями |
|||
b – зачаточные крылья. |
|
|||
|
|
|
|
|
|
|
|
|
|
При проведении анализирующего скрещивания самки (F1) из первого поколения с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% – серые длиннокрылые и 41,5% – черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% – черные длиннокрылые и 8,5% – серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов – АВ и аb, а отцовский – один тип – аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип АаВb и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb.

Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток – мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
АНАЛИЗИРУЮЩЕЕ СКРЕЩИВАНИЕ
ОПЫТ №1. ПОЛНОЕ СЦЕПЛЕНИЕ
(из F1 берется самец – АаBb)
P: |
♀ ааbb |
Х ♂ (F1) АаBb |
G: |
аb |
? |
F2: АаBb – 50% – серые с нормальными крыльями;
ааbb – 50% – черные с зачаточными крыльями.
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза), или в разных (транс-фаза) (рис.
9).
Рис. 9. 1 – механизм цис-фазы (некроссоверные гаметы); 2 – механизм транс-фазы (некроссоверные гаметы).
Следовательно, дигетерозиготный самец (АаBb) дает два сорта гамет:
●АB;
●аb
ОПЫТ №2. НЕПОЛНОЕ СЦЕПЛЕНИЕ. КРОССИНГОВЕР.
(из F1 берется самка – АаBb (рис. 10))
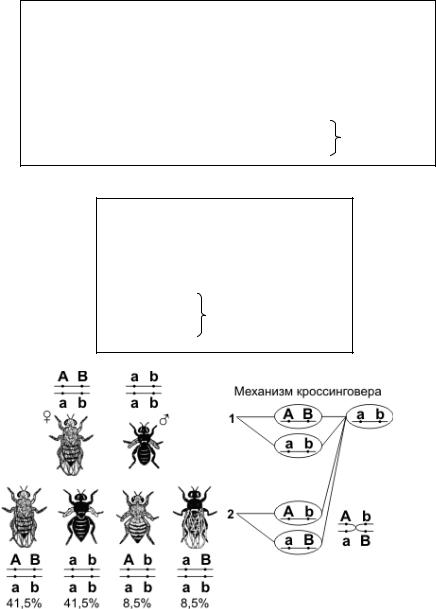
P: ♀( F1) АаBb |
Х ♂ ааbb |
|
G: |
? |
аb |
F2: АаBb – 41,5% – серые с нормальными крыльями; ааbb – 41,5% – черные с зачаточными крыльями;
ааBb – 8,5% – черные с нормальными крыльями Ааbb – 8,5% – серые с зачаточными крыльями
17% – КРОС-
СОВЕРНЫЕ
ОСОБИ
Следовательно, дигетерозиготная самка (АаBb) дает четыре сорта гамет:
●АB – 41,5%;
●аb – 41,5%;
●аB – 8,5%
●Аb – 8,5%
17% –
КРОССОВЕРН ЫЕ ГАМЕТЫ
Рис. 10. 1 – некроссоверные гаметы; 2 – кроссоверные гаметы.
Таким образом, у самки, как и у самца, гены А, B находятся в одной хромосоме. Но у самца они располагаются ближе друг к другу и удерживаются сильнее (ПОЛНОЕ СЦЕПЛЕНИЕ). У самки гены А и В находятся на некотором расстоянии друг от друга, сцеплены более слабо
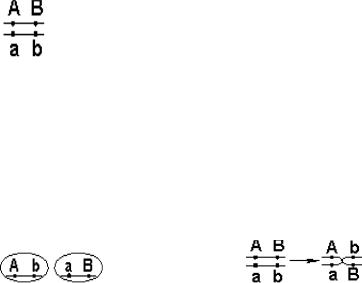
друг с другом, чем у самца, поэтому в 17% наблюдается кроссинговер (НЕПОЛНОЕ СЦЕПЛЕНИЕ).
Расстояние между генами измеряется в морганидах – условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозигота АаВb образует четыре типа гамет (АВ, Аb, аВ и аb) в равных количествах, то такая же дигетерозигота в случае сцепления генов А и В образует только два типа гамет – АВ и аb – тоже в равных количествах. Это некроссоверные гаметы
– гаметы, в процессе образования которых кроссинговер не произошел. Последние повторяют комбинацию генов в хромосоме родителя.
образуются гаметы:  Нерекомбинанты – гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Нерекомбинанты – гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Однако полное сцепление генов наблюдается достаточно редко, и в потомстве обычно бывают представлены все четыре фенотипа. Таким образом, и в этом случае при дигибридном скрещивании образуются новые сочетания признаков – рекомбинантные фенотипы. Было установлено, что кроме обычных гамет в этом случае возникают и другие
– Аb и аВ – кроссоверные гаметы, то есть появляются гаметы с новым сочетанием (комбинацией) аллелей, отличающимися от родительской гаметы.
образуются гаметы:
Причиной возникновения новых гамет является обмен участками гомологичных хромосом, или кроссинговер. Если особи с новыми генными комбинациями (рекомбинанты – гибридные особи, имеющие иное сочетание признаков, чем у родителей) встречаются в потомстве реже, чем особи с родительскими фенотипами, то это верный признак сцепленности соответствующих генов.
После опытов Томаса Моргана мушка дрозофила стала излюбленным объектом исследования генетиков, потому что:
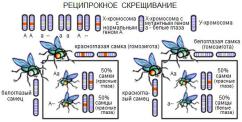
1)неприхотлива в содержании (легко размножается в лабораторных условиях при температуре 25°Ϲ);
2)выраженный половой диморфизм;
3)очень плодовита (большое число потомков);
4)быстрая смена поколений;
5)только 8 хромосом (небольшое число групп сцепления), но огромное количество альтернативных признаков.
Группа сцепления – гены, локализованные в одной хромосоме и наследующиеся совместно. Число групп сцепления соответствует гаплоидному набору хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены.
У человека количество групп сцепления зависит от пола – женский пол – 23 группы сцепления, у мужчин – 24 группы сцепления, поскольку две половые хромосомы (Х и Y) не являются гомологичными и несут различный набор генов.
29.Сцепленное с полом наследование. Примеры расписать.
Наследование, сцепленное с полом – это наследование признака, ген которого находится в половых хромосомах.
Т. Морган и его сотрудники заметили, что наследование окраски глаз у дрозофилы зависит от пола родительских особей, несущих альтернативные аллели. Красная окраска глаз доминирует над белой.
Реципрокное скрещивание (рис. 25) – два скрещивания, которые характеризуются взаимно противоположным сочетанием анализируемого признака и пола у форм, принимающих участие в этом скрещивании. Например, если в первом скрещивании самка имела доминантный признак, а самец – рецессивный, то во втором скрещивании самка должна иметь рецессивный признак, а самец – доминантный. Проводя реципрокное скрещивание, Т. Морган получил следующие результаты.
Рис. 25. Х-сцепленное наследование у дрозофилы.
Условные |
|
белые |
|
красные |
обозначения: |
P: |
♀ ХаХа |
x |
♂ ХАУ |
ХА – красные |
G: |
Ха |
|
ХА, У |
глаза; |
|
|||
|
|
|
|
|
Ха – белые глаза |
|
красные |
|
белые |
|
F1: |
ХАХа; |
|
ХаУ |
|
|
50% самки |
|
50% самцы |
При скрещивании красноглазого самца с белоглазой самкой в F1 получали равное число красноглазых самок и белоглазых самцов.
Однако при |
скрещивании белоглазого самца с красноглазой самкой в F1 |
||
были получены в равном числе красноглазые самцы и самки. |
|||
|
красные |
|
белые |
P: |
♀ ХАХА |
x |
♂ ХаУ |
G: |
ХА |
|
Ха, У |
|
красные |
|
красные |
F1: |
ХАХа; |
|
ХАУ |
|
50% самки |
|
50% самцы |
При скрещивании этих мух-гибридов F1 между собой были получены красноглазые самки, красноглазые и белоглазые самцы, но не было ни одной белоглазой самки.
|
красные |
|
красные |
|
P: |
♀ ХАХа |
x |
♂ ХАУ |
|
G: |
ХА, Ха |
|
ХА, У |
|
|
красные |
красные |
белые |
|
F1: |
ХАХА; ХАХа; |
|
ХАУ; |
ХаУ |
|
50% самки |
25% самцы |
25% самцы |
|
Тот факт, что у самцов частота проявления рецессивного признака была выше, чем у самок, наводил на мысль, что рецессивный аллель, определяющий белоглазость, находится в Х-хромосоме, а Y-хромосома лишена гена окраски глаз.
Чтобы |
проверить эту гипотезу, |
Морган скрестил исходного белоглазого |
||||
самца |
с красноглазой самкой |
из F1. |
В |
потомстве |
были |
получены |
красноглазые и белоглазые |
самцы |
и |
самки. Из |
этого |
Морган |
|
справедливо заключил, что только Х-хромосома несет ген окраски глаз. В Y-хромосоме соответствующего локуса вообще нет. Это явление известно под названием наследования, сцепленного с полом.
Гены, находящиеся в половых хромосомах, называют сцепленными с полом (рис. 26).
В Х-хромосоме имеется участок, для которого в Y-хромосоме нет гомолога. Наследованием, сцепленным с X-хромосомой, называют наследование генов в случае, когда мужской пол гетерогаметен и характеризуется наличием Y-хромосомы (XY), а особи женского пола гомогаметны и имеют две X-хромосомы (XX). У человека около 60 генов наследуются сцепленно с Х-хромосомой, в том числе гемофилия, дальтонизм (цветовая слепота), мышечная дистрофия, потемнение эмали зубов, одна из форм агаммаглобулинемии и другие.
Если аллель сцепленного с полом гена, находящегося в X-хромосоме, является рецессивным, то признак, определяемый этим геном, проявляется у всех особей гетерогаметного пола, которые получили этот аллель вместе с половой хромосомой, и у гомозиготных по этому аллелю особей гомогаметного пола. Это объясняется тем, что вторая половая хромосома (Y-хромосома) у гетерогаметного пола не несет аллелей большинства или всех генов, находящихся в парной хромосоме.
Таким признаком гораздо чаще будут обладать особи гетерогаметного пола, которые являются гемизиготными организмами. Поэтому заболеваниями, которые вызываются рецессивными аллелями, сцепленными с Х-хромосомой, гораздо чаще болеют мужчины, а женщины часто являются носителями таких аллелей.
Наследование таких признаков отклоняется от закономерностей, установленных Г.Менделем. Х-хромосома закономерно переходит от
одного пола к другому, |
при этом дочь наследует Х-хромосому отца, а |
|
сын |
Х-хромосому |
матери. Наследование, при котором сыновья |
наследуют признак |
матери, а дочери – признак отца получило, |
|
название крисс-кросс (или крест-накрест).

Рис. 26. Половые хромосомы человека.
Среди рецессивных признаков, сцепленных с Х-хромосомой, известны нарушения цветового зрения, так называемая цветовая слепота. В основе появления этих дефектов зрения лежит действие ряда генов. Краснозеленая слепота обычно называется дальтонизмом. Еще задолго до появления генетики в конце XVIII и в XIX в. было установлено, что цветовая слепота наследуется согласно вполне закономерным правилам. Так, если женщина, страдающая цветовой слепотой, выходит замуж за мужчину с нормальным зрением, то у их детей наблюдается очень своеобразная картина перекрестного наследования. Все дочери от такого брака получат признак отца, т.е. они имеют нормальное зрение, а все сыновья, получая признак матери, страдают цветовой слепотой.
В том же случае, когда наоборот, отец является дальтоником, а мать имеет нормальное зрение, все дети оказываются нормальными. В отдельных браках, где мать и отец обладают нормальным зрением, половина сыновей может оказаться пораженными цветовой слепотой (рис. 27). В основном наличие цветовой слепоты чаще встречается у мужчин.
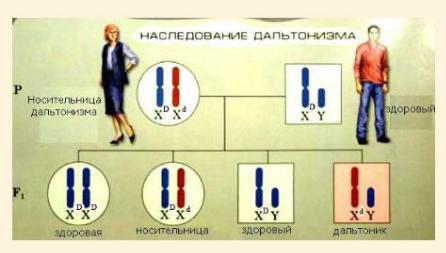
Рис. 27. Сыновья-дальтоники (25 %) в семье здоровых родителей.
Э.Вильсон объяснил наследование этого признака, предположив, что он локализован в Х-хромосоме и что у человека гетерогаметным (XY) является мужской пол. Становится вполне понятным, что в браке гомозиготной нормальной женщины (ХDХD) с мужчиной дальтоником (ХdY) все дети рождаются нормальными. Однако при этом, все дочери становятся скрытыми носителями дальтонизма, что может проявиться в последующих поколениях.
Другим примером наследования сцепленного с полом, может послужить рецессивный полулетальный ген, вызывающий несвертываемость крови на воздухе – гемофилию (табл. 3). Это заболевание появляется почти исключительно только у мальчиков. При гемофилии нарушается образование фактора VIII, ускоряющего свертывание крови. Ген, детерминирующий синтез фактора VIII, находится в участке Х-хромосомы (доминантный – нормальный; рецессивный – мутантный).
Таблица 3 Рецессивный полулетальный ген, вызывающий гемофилию
Возможные |
Возможные |
генотипы: |
фенотипы: |
|
|
ХH ХH |
Нормальная женщина |
ХH Хh |
Нормальная женщина (носитель) |
ХHY |
Нормальный мужчина |
ХhY |
Мужчина-гемофилик |
XhXh |
В гомозиготном состоянии у женщин ген |
|
гемофилии летален |
|
|
Рис. 28. Генеалогическое древо царствовавших семей Европы, иллюстрирующее наследование гена гемофилии, локализованного в X- хромосоме человека
Особей женского пола, гетерозиготных по любому из сцепленных с полом признаков, называют носителями соответствующего рецессивного гена. Они фенотипически нормальны, но половина их гамет несет рецессивный ген.
Несмотря на наличие у отца нормального гена, сыновья матерейносителей с вероятностью 50% будут страдать гемофилией. Один из наиболее хорошо документированных примеров наследования гемофилии – это родословная потомков английской королевы Виктории
(рис. 28).
Предполагают, что ген гемофилии возник в результате мутации у самой королевы Виктории или у одного из ее родителей. Среди унаследовавших это врожденное заболевание – цесаревич Алексей, сын последнего русского царя Николая II. Мать цесаревича, царица Александра Федоровна (Алиса), получила от своей бабушки королевы Виктории ген гемофилии и передала его в четвертом поколении бывшему наследнику царского престола (рис. 29).

Рис. 29. Королева Виктория, царица Александра Федоровна (Алиса), цесаревич Алексей, сын последнего русского царя Николая II.
Один из сцепленных с полом рецессивных генов вызывает особый тип мышечной дистрофии (тип Дюшена). Эта дистрофия проявляется в раннем детстве и постепенно ведет к инвалидности и смерти ранее 20-летнего возраста. Поэтому мужчины с дистрофией Дюшена не имеют потомства, а женщины гетерозиготные по гену этого заболевания, вполне нормальны.
Среди доминантных признаков, сцепленных с Х-хромосомой, можно указать на ген, который вызывает недостаточность органического фосфора в крови. В результате, при наличии этого гена, часто развивается рахит, устойчивый к лечению обычными дозами витамина D. В этом случае картина сцепленного с полом наследования заметно отличается от того хода передачи по поколениям, который был описан для рецессивных болезней. В браках женщин-носительниц со здоровыми мужчинами половина детей больных и половина потомков – здоровые. Здесь, в соответствии с характером наследование доминантного гена, в Х-хромосомах произошло расщепление по генотипу в соотношении
1:1:1:1.
|
|
рахит |
|
норма |
|
Условные |
P: |
♀ ХАХа |
x |
♂ ХаУ |
|
обозначения: |
|
|
|
|
|
ХА – рахит; |
G: |
ХА; Ха |
|
Ха, У |
|
Ха – норма |
|
|
|
|
|
|
|
рахит |
норма |
рахит |
норма |
|
F1: |
ХАХа; |
ХаХа; |
ХАУ; ХаУ |
|
|
|
25% ♀ |
25% ♀ |
25% ♂ |
25% ♂ |
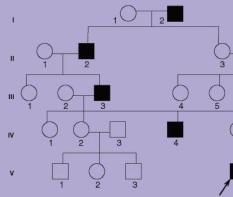
Другим примером доминантного гена, локализованного в Х- хромосоме человека, может послужить ген, вызывающий дефект зубов, приводящий к потемнению эмали зубов.
Большинство генов, имеющихся в Х-хромосоме, в Y-хромосоме отсутствует, однако в этих негомологичных хромосомах есть гомологичные участки. Тогда гены, расположенные в представленных локусах наследуются по аутосомному принципу (например, геморрагический диатез).
Условные |
|
норма |
|
норма |
|
||
обозначения: |
|
|
|
||||
P: |
♀ ХАХа |
x |
♂ ХаУА |
|
|||
ХА – норма; |
G: |
ХА; Ха |
|
Ха, УА |
|
||
Х |
а |
– геморрагический |
|
|
|||
|
|
|
|
|
|
||
диатез; |
|
|
|
|
|
||
УА – норма; |
|
норма |
диатез |
норма |
|||
Уа – геморрагический |
|
норма |
|
|
|
||
диатез. |
F1: |
ХАХа; |
ХаХа; |
ХАУА; |
ХаУА |
||
|
|
|
|
25% ♀ |
25% ♀ |
25% ♂ |
25% ♂ |
передается от отца всем его сыновьям, и только им. Следовательно, для генов, содержащихся только в Y- хромосоме, характерно
голандрическое наследование,
т.е. они передаются от отца к сыну и проявляются только по мужской линии (рис. 30).
Рис. 30. Родословная при голандрическом наследовании.
Кроме того есть гены,
присутствующие только в Y-
хромосоме. Y-хромосома
Y-хромосому называют генетически инертной или генетически пустой, так как в ней очень мало генов. У человека в Y-хромосоме содержатся по крайней мере три гена, один из которых необходим для дифференциации семенников, второй требуется для проявления антигена
гистосовместимости, а третий оказывает влияние на размер зубов. Y- хромосома имеет немного признаков, среди которых есть патологические.
Патологические признаки наследуются по параллельной схеме наследования (100% проявление по мужской линии). К ним относят: 1) облысение; 2) гипертрихоз (оволосенение козелка ушной раковины в зрелом возрасте); 3) наличие перепонок на нижних конечностях; 4) ихтиоз (чешуйчатость и пятнистое утолщение кожи).
НАСЛЕДОВАНИЕ ПРИЗНАКОВ, КОНТРОЛИРУЕМЫХ ПОЛОМ
Признаки, выражение или проявление которых различно у представителей разных полов, или проявляющиеся только у одного пола, относятся к ограниченным полом. Они могут определяться генами, расположенными как в аутосомах, так и в половых хромосомах.
Имеется ряд признаков, контролируемых генами, расположенными в аутосомах, однако для проявления этих признаков необходима определенная среда, создаваемая генами, находящимися в половых хромосомах. Например, гены, определяющие мужские признаки, находятся в аутосомах, и их фенотипические эффекты маскируются наличием пары Х-хромосом, в присутствии одной Х-хромосомы мужские признаки проявляются. Такие признаки называются обусловленными или контролируемыми полом.
Появление лысины – аутосомно-доминантный признак, но проявляется практически только у мужчин при наследовании, контролируемом полом. У женщин подавляются гены, детерминирующие рост бороды.
Примером у животных может служить наследование комолости (отсутствие рогов) у овец. У дорсетской породы овец оба пола рогаты. У суффольской породы, напротив, оба пола комолы. При скрещивании овец разных пород в F1 получают рогатых баранов и комолых ярок (самок).
Известно, что многие признаки проявляются у представителей только одного пола (например, способность давать молоко). Однако гены, определяющие этот признак, есть не только у коров, но и у быков. Установить, какова потенциальная способность быка давать высокомолочное потомство, вполне возможно, хотя и косвенным путём. Для этого необходимо получить сведения о молочности его дочерей. Более того, один из самых эффективных способов быстрого повышения удоев состоит в интенсивном отборе быков по их потенциальной молочности.
30.Основные положения хромосомной теории Т. Моргана.
Предположение о том, что гены локализуются в хромосомах, появилось еще в начале ХХ века. Так в 1902-1903 годах американский цитолог У. Сеттон и немецкий цитолог и эмбриолог Т. Бовери независимо друг от друга выявили параллелизм в поведении генов и хромосом в ходе формирования гамет и оплодотворения. Однако экспериментальное доказательство локализации конкретных генов в конкретных хромосомах было получено только в 1910 г. американским генетиком Т. Морганом, который в последующие годы (1911-1926) обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности.
Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:
гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
гены расположены в хромосомах в определенной линейной последовательности;
гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
каждый вид имеет характерный только для него набор хромосом
– кариотип.

31.Взаимодействия аллельных генов: полное и неполное доминирование, сверхдоминирование, кодоминирование, аллельное исключение. Примеры
Аллельные гены – различные формы одного и того же гена, расположенные в одинаковых участках (локусах) гомологических хромосом. Аллели определяют варианты развития одного и того же признака. В нормальной диплоидной клетке могут присутствовать не более двух аллелей одного локуса одновременно. Признак, обусловленный взаимодействием одной аллельной пары, называют простым, а форму взаимодействия – аллельной.
1. Полное доминирование (рис. 32) – вид взаимодействия аллельных генов, при котором в гетерозиготном состоянии доминантный аллель полностью подавляет проявление рецессивного аллеля, фенотипически гибриды F1 100% обладают доминантными признаками (гетерозигота имеет фенотип одного из родителей – фенотип доминантной гомозиготы).
При наследовании всех менделирующих признаков наблюдается полное доминирование. Менделирующие признаки – признаки, наследующиеся по законам Менделя, например: альбинизм – рецессивный признак, не проявляется в гетерозиготном состоянии. Поэтому у гибридов F2 наблюдается расщепление по генотипу 1:2:1, по фенотипу – 3:1.
Рис. 32. Полное доминирование.
ПОЛНОЕ |
F2 |
по генотипу: |
1 : 2 : 1 |
|
|||
ДОМИНИРОВАПНИЕ |
|
по фенотипу: |
3 : 1 |
|
|
|
|
2. Неполное доминирование (рис. 33) – вид взаимодействия аллельных генов, при котором в гетерозиготном состоянии доминантный аллель
неполностью подавляет проявление рецессивного аллеля. В результате имеет место промежуточный характер наследования (гетерозигота имеет промежуточный фенотип).
Рис. 33. Неполное доминирование.
У гибридов F2 расщепление по генотипу и фенотипу совпадают – 1:2:1.
НЕПОЛНОЕ |
F2 |
по генотипу: |
1 : 2 : 1 |
ДОМИНИРОВАПНИЕ |
по фенотипу: |
1 : 2 : 1 |
|
|
|
|
|
Другим примером неполного доминирования служит серповидноклеточная анемия – наследственная гемоглобинопатия, связанная с таким нарушением строения белка гемоглобина, при котором он приобретает особое кристаллическое строение – так называемый гемоглобин S. Эритроциты, несущие гемоглобин S вместо нормального гемоглобина А, под микроскопом имеют характерную форму серпа (рис. 34), за что эта форма гемоглобинопатии и получила название серповидноклеточной анемии.
Эритроциты, несущие гемоглобин S, обладают пониженной стойкостью и пониженной кислород-транспортирующей способностью, поэтому у больных с серповидноклеточной анемией повышено разрушение эритроцитов в селезенке, укорочен срок их жизни, повышен гемолиз и часто имеются признаки хронической гипоксии.
Серповидноклеточная анемия наследуется по аутосомно-рецессивному типу. У больных, гетерозиготных по гену серповидно-клеточной анемии (Аа), наряду с серповидными эритроцитами, несущими гемоглобин S, в крови наличествуют и нормальные, несущие гемоглобин А. При этом болезнь менее выражена клинически, протекает легче, а иногда вообще не вызывает симптомов, и серповидные эритроциты выявляются случайно при лабораторном исследовании крови. У гомозигот (аа) по гену
серповидноклеточной анемии в крови имеются только серповидные эритроциты, несущие гемоглобин S, болезнь протекает тяжело и заканчивается летальным исходом.
Рис. 34. Строение эритроцитов при серповидноклеточной анемии.
Серповидноклеточная анемия весьма распространена в регионах мира, эндемичных по малярии, причем больные серповидноклеточной анемией обладают повышенной (хотя и не абсолютной) врожденной устойчивостью к заражению различными штаммами малярийного плазмодия (рис.
35).
Рис. 35. Распространенность серповидноклеточной анемии и малярии.
3.Сверхдоминирование (рис. 36)
– вид взаимодействия аллельных генов, при котором фенотипическое выражение признака у гетерозиготы (Аа) намного ярче, чем у доминантной гомозиготы (АА). Этот вид взаимодействия генов лежит в основе гетерозиса.
4. Аллельное исключение (рис. 37) – вид взаимодействия аллельных генов, при котором в части соматических клеток организма, гетерозиготного по данному локусу, активен один аллель, тогда как в

других клетках другой. Такой вид взаимодействия аллельных генов наблюдается при наследовании признаков у организмов-мозаиков.
Рис. 37. Аллельное исключение.
5. Кодоминирование – вид взаимодействия аллельных генов, при котором у гетерозиготы отсутствуют доминантно-рецессивные отношения, аллели фенотипически проявляют себя как бы независимо друг от друга.
32.Специфика проявления генов в признак - экспрессивность, пенетрантность, плейотропия, генокопии.
Плейотропия – множественное действие гена (один ген контролирует развитие нескольких признаков).
Часто наблюдается явление, когда одна пара генов влияет сразу на несколько признаков. Еще Мендель в своих опытах установил, что растения гороха, имеющие пурпуровые цветки, кроме того всегда имеют красные пятна в пазухах листьев и образуют семена, покрытые серой или бурой кожурой, и что все эти признаки зависят от действия одного наследственного фактора.
При изучении генетических особенностей персидской пшеницы Н.И.Вавилов установил, что доминантный ген черной окраски всегда одновременно вызывает и сильное опушение чешуй.
Уплодовой мушки дрозофилы ген, определяющий отсутствие пигмента в глазах, снижает плодовитость, влияет на окраску некоторых внутренних органов и уменьшает продолжительность жизни.
Укаракульских овец одним геном определяются цвет шерсти и развитие рубца.
Плейотропное действие гена может быть первичным и вторичным (рис.
56).
Рис. 56. Виды плейотропии.
При первичной плейотропии ген проявляет свой множественный эффект. Например, при болезни Хартнупа мутация гена приводит к нарушению всасывания аминокислоты триптофана в кишечнике и его реабсорбции в почечных канальцах. При этом поражаются одновременно мембраны эпителиальных клеток кишечника и почечных канальцев с расстройствами пищеварительной и выделительной систем.
Рис. 57. Синдром Марфана.
У человека известен доминантный ген, определяющий признак "паучьи пальцы" (синдром Марфана). Одновременно ненормальное развитие пальцев сопровождается нарушением строения хрусталика и развитием порока сердца (рис. 57). Здесь в основе множественного эффекта тоже лежит действие одного гена, вызывающего нарушение развития соединительной ткани.
При вторичной плейотропии есть один первичный фенотипический признак – проявление мутантного гена, вслед за которым развивается ступенчатый процесс вторичных изменений, приводящих к множественным эффектам. Так, при серповидноклеточной анемии у гомозигот наблюдается несколько патологических признаков: анемия, увеличенная селезенка, поражение кожи, сердца, почек и мозга. Поэтому гомозиготы с геном серповидноклеточной анемии гибнут, как правило, в детском возрасте. Все эти фенотипические проявления гена составляют иерархию вторичных проявлений. Первопричиной, непосредственным фенотипическим признаком проявлением дефектного гена является аномальный гемоглобин и эритроциты серповидной формы. Вследствие этого происходят последовательно другие патологические процессы: слипание и разрушение эритроцитов, анемия, дефекты в почках, сердце, мозге – эти патологические признаки вторичны.
При плейотропии, ген, воздействуя на какой-то один основной признак, может также менять, модифицировать проявление других генов, в связи с чем введено понятие о генах-модификаторах. Последние усиливают или ослабляют развитие признаков, кодируемых "основным" геном.
Модифицирующее действие гена – ген усиливает или ослабляет действие неаллельного гена. Существуют гены "основного действия", т.е. такие, которые определяют развитие признака или свойства, например выработку пигмента, форму плода, чувствительность или устойчивость к заболеваниям и т.д. Наряду с такими генами, по-видимому, существуют гены, которые сами по себе не определяют какую-либо качественную реакцию или признак, они лишь усиливают или ослабляют проявление действия "основного" гена, т.е. модифицируют его, – такие гены получили название модификаторов. Любые взаимодействующие гены в одно и то же время являются генами "основного" действия по одному признаку, а по другому (или другим) являются генами-модификаторами.
Показателями зависимости функционирования наследственных задатков от характеристик генотипа является пенетрантность и экспрессивность. Рассматривая действие генов, их аллелей необходимо учитывать и модифицирующее влияние среды, в которой развивается организм. Если растения примулы скрещивать при температуре 15-20 °С, то в F1 согласно менделевской схеме, все поколения будут иметь розовые цветы. Но когда такое скрещивание проводить при температуре 35 °С, то все гибриды будут иметь цветы белого цвета. Если же осуществлять скрещивания при температуре около 30 °С, то возникает разное соотношение (от 3:1 до 100%) растений с белыми цветами.
Такое колебание классов при расщеплении в зависимости от условий среды получило название пенетрантность – сила фенотипического проявления. Итак, пенетрантность – это частота проявления гена, явление появления или отсутствия признака у организмов, одинаковых по генотипу.
Пенетрантность значительно колеблется как среди доминантных, так и среди рецессивных генов. Наряду с генами, фенотип которых появляется только при сочетании определенных условий и достаточно редких внешних условий (высокая пенетрантность), у человека есть гены, фенотипическое проявление которых происходит при любых соединениях внешних условий (низкая пенетрантность). Пенетрантность измеряется процентом (%) организмов с фенотипическим признаком от общего количества обследованных носителей соответствующих аллелей.
Если ген полностью, независимо от окружающей среды, определяет фенотипическое проявление, то он имеет пенетрантность 100 % (полная пенетрантность). Однако, некоторые доминантные гены проявляются менее регулярно. Так, полидактилия имеет четкое вертикальное наследования, но бывают пропуски поколений. Доминантная аномалия – преждевременное половое созревание – присуще только мужчинам, однако иногда заболевание может передаться от человека, который не страдал этой патологией. Пенетрантность указывает, в каком проценте носителей гена оказывается соответствующий фенотип. Итак, пенетрантность зависит от генов, от среды, от того и другого. Таким образом, это не константное свойство гена, а функция генов в конкретных условиях среды.
Экспрессивность (лат. “ехргеssio” – выражение) – это изменение количественного проявления признака в разных особях-носителях соответствующего аллеля.
При доминантных наследственных заболеваниях экспрессивность может колебаться. В одной и той же семье могут проявляться наследственные болезни от легких, едва заметных до тяжелых: различные формы гипертонии, шизофрении, сахарного диабета и т.д. Рецессивные наследственные заболевания в пределах семьи проявляются однотипно и имеют незначительные колебания экспрессивности.
Таким образом, пенетрантность – это вероятность фенотипического проявления гена, которая выражается в процентах (отношение больных особей к числу носителей соответствующего гена).
Экспрессивность – степень клинического проявления гена, которая может быть слабой или сильной. Пенетрантность и экспрессивность генов зависят от эндогенных и экзогенных факторов. Например, если для
проявления гемофилии решающее значение имеет нарушение в геноме, то возникновение сахарного диабета зависит от взаимодействия генетических факторов и внешней среды. В последнем случае говорят о наследственном предрасположении. Способность генотипа по-разному проявляться в различных условиях среды называется нормой реакции. Норма реакции наследуется, а изменения в пределах нормы реакции не наследуются.
Ряд сходных по внешнему проявлению признаков, в том числе и наследственных болезней, может вызываться различными неаллельными генами. Такое явление называется генокопией. Биологическая природа генокопий заключается в том, что синтез одинаковых веществ в клетке в ряде случаев достигается различными путями.
Одним из первых это явление изучал Н.В.Тимофеев-Ресовский. По предложению Н.В.Тимофеева-Ресовского, с середины 1930-х годов гетерогенными группами стали называть группы генов, дающих весьма сходное внешнее проявление, но локализованных в разных хромосомах или разных локусах, как, например, группа генов minute, обусловливающих редукцию щетинок у дрозофилы. Это явление широко распространено в живой природе, включая человека. Правда, в генетической литературе для его обозначения обычно используют не термин Н.В. Тимофеева-Ресовского «гетерогенные группы», а более поздний (середины 40-х годов XX века), предложенный немецким генетиком X.Нахтсхаймом – «генокопии», в дополнение к которому позже был введен термин «фенокопии».
Внаследственной патологии человека большую роль играют также фенокопии – модификационные изменения. Они обусловлены тем, что в процессе развития под влиянием внешних факторов признак, зависящий от определенного генотипа, может измениться; при этом копируются признаки, характерные для другого генотипа. Таким образом, фенокопия
– ненаследственное изменение фенотипа организма, вызванное действием определённых условий среды и копирующее проявление какого-либо известного наследственного изменения – мутации – у этого организма.
Вразвитии фенокопий могут играть роль разнообразные факторы среды
– климатические, физические, химические, биологические и социальные. Врожденные инфекции (краснуха, токсоплазмоз, сифилис) также могут стать причиной фенокопий ряда наследственных болезней и пороков развития. Существование гено- и фенокопий нередко затрудняет постановку диагноза, поэтому существование их врач всегда должен иметь в виду.

33. Множественный аллелизм. Группы крови человека по системе АВО
(генотипы, фенотипы, наследование, правила переливания)
Кодоминирование – вид взаимодействия аллельных генов, при котором у гетерозиготы отсутствуют доминантно-рецессивные отношения, аллели фенотипически проявляют себя как бы независимо друг от друга. Примером кодоминантного взаимодействия аллелей является наследование IV группы крови у человека.
В популяциях людей известны 4 антигенные группы крови человека: А, В, АВ, О и три типа аллелей этого гена: IA, IB, i (табл. 4).
|
|
|
|
Таблица 4 |
|
|
Особенности наследования групп крови |
|
|
||
|
|
|
|
|
|
|
|
|
Фенотипы |
|
|
Аллели |
Генотипы |
|
|
|
|
Группы |
Агглюти- |
Агглюти- |
|||
в |
|
крови |
ногены |
нины |
|
популя- |
|
|
(антигены) |
(антитела |
|
ции |
|
|
на |
) в |
|
|
|
|
мембране |
плазме |
|
|
|
|
эритроцитов |
крови |
|
|
|
|
|
|
|
i |
ii |
I(O) |
─ |
α, β |
|
|
|
|
|
|
|
IA |
IAIA, IAi |
II (A) |
А |
β |
|
IB |
IBIb, IВi |
III(B) |
В |
α |
|
|
IAIB |
IV (AB) |
А, В |
─ |
|
─кодоминирова
ние
Генотипы |
IAIA, IAi |
имеют |
эритроциты, |
содержащие на |
|
поверхности только антиген А, а в плазме крови гемагглютинин β (группа крови А или II). Генотипы IBIb, IВi имеют эритроциты, содержащие на поверхности только антиген В, а в плазме крови гемагглютинин α (группа крови В или III). Генотип IAIB
имеет эритроциты, несущие оба антигена А и В, а в плазме крови отсутствуют гемагглютинины α и β (группа крови IV или АВ). Это и есть случай кодоминирования
(рис. 38).
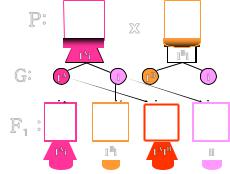
P: |
x |
|
|
|
|
II |
|
III |
|
|
|
IAi |
|
IВi |
G: IA |
i IВ |
i |
F1 :
IAi |
IВi |
IAIВ |
ii |
II |
III |
IV |
I |
|
|
КОДОМИНИРОВАНИЕ |
|
Рис. 38. Наследование IV группы крови.
У людей с генотипом ii на поверхности эритроцитов нет антигенов А и В, а в плазме крови присутствуют гемагглютинины α и β.
Агглютинин α специфически связывается и осаждает эритроциты с антигеном А, агглютинин β – эритроциты с антигеном В. На этих взаимоотношениях основана система переливания крови. Долгие годы придерживались так называемого закона Отенберга, согласно которому агглютинируют только эритроциты перелитой донорской крови (а не эритроциты реципиента), учитывая, что агглютинины донорской крови разводятся в крови реципиента и не способны агглютинировать его эритроциты. Это обстоятельство разрешало переливать наравне с одногруппной и кровь другой группы, сыворотка которой не
агглютинировала эритроциты реципиента. |
|
|
|||||
На |
практике |
используется |
поэтому лиц с I группой крови |
||||
следующая схема (рис. 39): |
называют |
универсальными |
|||||
реципиенту |
0(I) |
группы |
донорами, но им самим можно |
||||
допустимо |
|
переливать |
вводить кровь только I группы. |
||||
донорскую |
кровь |
только 0(I) |
Кровь от донора IV группы |
||||
группы, реципиентам А(II) группы |
можно переливать только лицам |
||||||
– донорскую кровь А(II) и 0(I) |
данной группы, но им самим |
||||||
групп, реципиентам В (III) группы |
можно переливать |
кровь всех |
|||||
– донорскую кровь В (III) и 0(I) |
четырех групп. Людей с IV |
||||||
групп, |
реципиентам |
АВ(IV) |
группой |
крови |
называют |
||
группы – донорскую кровь всех |
универсальными |
|
|||||
четырех |
групп. |
|
Любому |
реципиентами. |
|
||
реципиенту |
можно |
вводить |
|
|
|
||
кровь I группы (0), так как ее |
|
|
|
||||
эритроциты |
не |
|
содержат |
|
|
|
|
агглютиногены и не склеиваются, |
|
|
|
||||
Рис. 39. Схема переливания крови с учетом группы крови.
6. Множественный аллелизм – присутствие в генофонде популяции
одновременно |
|
3 |
и |
более |
|||
различных аллелей одного гена. |
|
|
|||||
Примером |
|
может |
служить |
|
|
||
разнообразие |
окраски |
глаз |
|
|
|||
человека, |
разнообразие |
групп |
|
|
|||
крови. Ген |
I может |
быть |
|
|
|||
представлен |
тремя |
разными |
|
|
|||
аллелями: IA, IB, i, которые |
|
|
|||||
комбинируются в зиготах только |
|
|
|||||
попарно. |
|
|
|
|
|
|
|
Другим |
примером |
является |
|
|
|||
наследование окраски шерсти у |
Рис. 40. Наследование |
окраски |
|||||
кроликов (рис. 40). В популяции |
|||||||
шерсти в популяции кроликов. |
|||||||
кроликов |
существует |
|
четыре |
||||
|
|
|
|||||
аллеля.
Ген А отвечает за наследование темной окраски шерсти и доминируют над всеми другими аллелями. Ген ach обуславливает шиншилловую окраску и по отношению к генам ah и a ведет себя как доминантный. Ген ah отвечает за гималайскую окраску шерсти и доминирует над геном a (белая окраска). Таким образом, А > ach > ah > a (табл. 5).
Таблица 5
Генотипы и фенотипы кроликов
Фенотипы |
Аллели |
Генотипы |

Темная окраска |
А |
АА, Ааch, Ааh, Аа |
Шиншилловая окраска |
аch |
аchаch, аchаh, аchа |
Гималайская окраска |
аh |
аhаh, аhа |
Белая окраска |
а |
аа |
|
|
|
Причиной множественного аллелизма являются случайные изменения структуры гена (мутации), сохраняемые в процессе естественного отбора в генофонде популяции.
Многообразие аллелей, рекомбинирующихся при половом размножении, определяет степень генотипического разнообразия среди представителей данного вида, что имеет большое эволюционное значение, повышая жизнеспособность популяций в меняющихся условиях их существования. Кроме эволюционного и экологического значения аллельное состояние генов оказывает большое влияние на функционирование генетического материала. В диплоидных соматических клетках эукариотических организмов большинство генов представлено двумя аллелями, которые совместно влияют на формирование признаков.
34. Взаимодействие неаллельных генов - комплементарность, эпистаз,
полимерия. Примеры.
Неаллельные гены – это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки.
Неаллельные гены также могут взаимодействовать между собой. При этом один признак проявляется под действием совокупности нескольких генов. Выделяют несколько форм взаимодействия неаллельных генов:
1)комплементарность;
2)эпистаз;
3)полимерия;
4)эффект положения.
КОМПЛЕМЕНТАРНОЕ ДЕЙСТВИЕ ГЕНОВ – это вид взаимодействия неаллельных генов по принципу дополнения или комбинирования, когда доминантные аллели этих генов при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков.
Взаимодействие аллелей с новообразованием – межаллельная комплементация. В некоторых случаях при объединении в гибриде разных аллелей независимого происхождения, рецессивных по отношению к дикому типу, наблюдают восстановления нормы, т.е. признака дикого типа. Разные рецессивные аллели одного и того же гена
отличаются друг от друга тем, что кодируют пептиды с повреждением разных доменов. Если в гетерозиготе (компаунде) объединятся аллели с разными повреждениями таким образом, что в молекуле фермента, состоящей из субъединиц-продуктов одного и того же гена, соберутся все необходимые функциональные центры, то ферментативная активность будет восстановлена.
КОМПЛЕМЕНТАРНОСТЬ (расщепление по фенотипу)
9: 7
9: 3: 4 F2 : 9: 6: 1
9: 3: 3: 1
Комплементарность (взаимодополняющий эффект)
Явление, когда признак развивается при взаимодополняющем действии доминантных неаллельных генов. По типу комплементарности взаимодействуют гены, контролирующие разные этапы одного и того же метаболического пути. Впервые такой тип взаимодействия генов был описан Бетсоном и Пеннетом при анализе наследования окраски венчика у душистого горошка. При скрещивании растений душистого горошка с белыми цветами в первом поколении все растения имели пурпурные цветы (как у дикого вида душистого горошка) формы, а во втором поколении наблюдалось расщеплении в соотношении 9
пурпурные : 7 белые.
Для объяснения полученного результаты было сделано предположение, что синтез пурпурного пигмента контролируют два неаллельных доминантных гена, продукты которых задействованы на разных этапах метаболического пути. Мутация любого из этой пары гена прерывает процесс синтеза пигмента. Взятые для скрещивания растения в своем генотипе имели мутации разных комплементарных генов.
Схема скрещивания: |
|
|
Р: белые венчики |
|
белые венчики |
♀ ААbb |
х |
♂ ааВВ |
Условные обозначения:
А – отвечает за синтез пропигмента – бесцветного предшественника пигмента; а – не происходит синтеза пропигмента; В – определяет синтез
G: Аb |
аВ |
F1: АаВb – 100% пурпурные венчики
F2: 9/16 – пурпурные (А•В•) : 7/16 белые ( 3/16 – А•bb, 3/16 – ааВ•; 1/16 – ааbb).
Вторым примером взаимодополняющего действия может служить наследование окраски шерсти у мышей (рис. 41), когда при скрещивании чистопородных линий черных и белых мышей, все гибриды первого поколения (F1) имеют серую окраску шерсти
(комплементарная), а в F2
наблюдается расщепление по фенотипу 9 серые : 3 черные : 4 белые.
Рис. 41. Наследование окраски шерсти у мышей.
При этом первая пара аллельных генов отвечает за синтез пигмента (С – синтез пигмента; с – отсутствие пигмента (альбинизм)), вторая пара генов
– за распределение пигмента по длине волоса (А – зональное распределение пигмента; а – не определяет зонального распределения пигмента).
Формула дигибридного расщепления по фенотипу:
9 |
: |
3 |
: |
3 |
: |
1 |
А•C• |
|
ааC• |
|
А•cc |
|
ааcc |
серая |
|
черная |
|
белая |
|
белая |
|
|
|
|
|
|
|
9 |
: |
3 |
: |
4 |
|
|
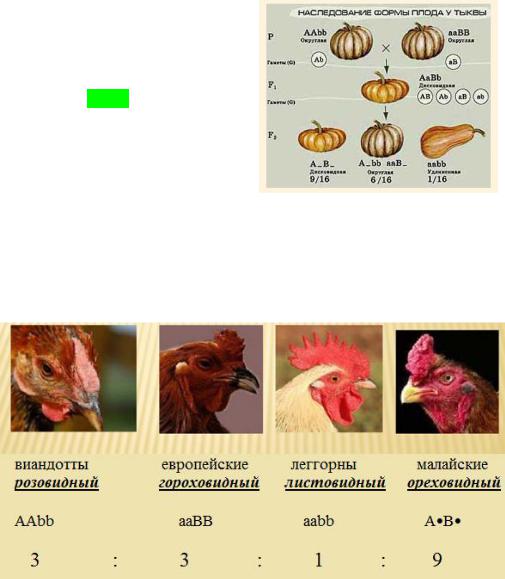
Третьим примером взаимодополняющего действия является наследование формы плода у тыквы (рис.42). В результате комплементарного взаимодействия генов в F2 наблюдается соотношение
9: 6: 1.
Рис. 42. Генотипы: А•bb – округлая форма; ааВ• – округлая форма;
А•В• – дисковидная форма (комплементарная); ааbb – удлиненная форма.
Комплементарность (комбинативное взаимодействие)
Один из первых примеров взаимодействия неаллельных генов был обнаружен в начале XX в. при анализе наследования формы гребня у кур. Описано четыре разновидности форм гребней, при этом разные породы имеют характерную морфологию гребня (рис. 43):
Рис. 43. Разновидности форм гребней у разных пород кур.
леггорны – листовидный (простой гребень),
виандотты – розовидный (низкий, утолщенный спереди, заостренный сзади и у основания покрытый сосочками),
европейские – гороховидный (невысокий, с тремя продольными пластинками),
малайские – ореховидный, напоминающий поверхность половинки грецкого ореха.
В результате скрещиваний кур, имеющих розовидный и гороховидный гребни, в F1 возникает новая форма гребня – ореховидный (из-за взаимодействия генов A и B).
Р: |
Розовидный |
|
Гороховидный |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
♀ AAbb |
х |
♂ aaBB |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
G: |
Ab |
|
|
|
aB |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
F1: |
AaBb – 100% |
Ореховидный |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
Скрещивание гибридов F1 дает следующие результаты в F2: |
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||
|
|
|
|
AB |
|
|
Ab |
|
|
aB |
|
|
|
|
Ab |
|||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
AB |
|
Орех. |
|
|
|
Орех. |
|
|
|
|
Орех. |
|
|
|
|
|
Орех. |
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
AABB |
|
|
|
AABb |
|
|
|
|
AaBB |
|
|
|
|
|
AaBb |
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
Ab |
|
Орех. |
|
|
Розов. |
|
|
|
Орех. |
|
|
|
|
Розов. |
|
|
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
ААВb |
|
|
|
ААbb |
|
|
|
АаВb |
|
|
|
|
|
Ааbb |
|
|
|
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
аB |
|
Орех. |
|
|
|
Орех. |
|
|
Горох. |
|
|
|
Горох. |
|
|
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
АаВВ |
|
|
|
АаВb |
|
|
|
ааВВ |
|
|
|
|
|
ааВb |
|
|
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||
|
|
аb |
|
Орех. |
|
|
Розов. |
|
|
Горох. |
|
|
Листов. |
|
||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
АаВb |
|
|
|
Ааbb |
|
|
|
ааВb |
|
|
|
|
|
ааbb |
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Потомство F2 характеризуется следующими особенностями:
1.Присутствие доминантных аллелей двух генов А и В у 9/16 кур второго поколения ведет к образованию ореховидного гребня.
2.Присутствие гена А в гомоили гетерозиготном состоянии при рецессивном b дает розовидную форму у 3/16 особей, а гены ааВ – у 3/16 потомства дают гороховидный гребень.
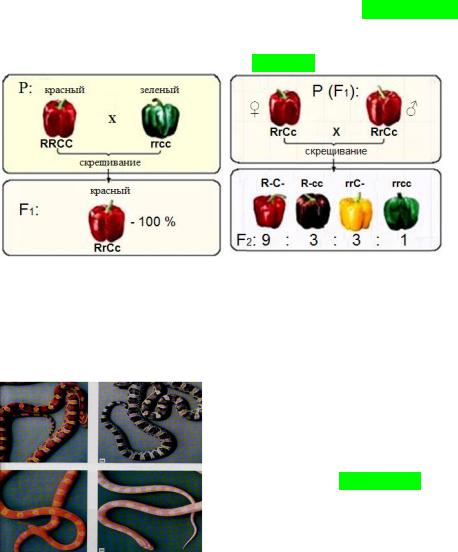
3. Гомозиготы по обоим рецессивным генам ааbb имеют новый фенотип
– простой листовидный гребень. Этот признак в последующих скрещиваниях не дает расщепления.
Итак, взаимодействие доминантных генов А и В изменяет форму гребня. В этом случае расщепление в дигибридном скрещивании нарушается, однако очевидно, что общее соотношение вариантов 9 : 3 : 3 : 1 сохраняется.
Наследование окраски плодов у перца – 9 : 3 : 3 : 1 (рис. 44).
Рис. 44. Разновидности окраски перца: генотипы R•cc – коричневая окраска,
rrC• – желтая; R•C• – красная (комплементарная); rrcc – зеленая.
Нследование окраски тела у змей (рис. 45).
Во втором гибридном поколении наблюдается расщепление в
На соотношении 9 : 3 : 3 : 1. При этом ген А отвечает за синтез
красного пигмента, а – отсутствие синтеза красного пигмента; ген В обусловливает полосатый рисунок, b – ромбовидный рисунок.
Рис. 45. Генотипы змей: (а) –

ААВВ; (b) – ааВВ; (с) – ААbb; ( d ) – ааbb
ЭПИСТАЗ (подавление) – ген одной аллельной пары подавляет действие генов другой аллельной пары. Ген, подавляющий действие другого гена,
называется эпистатическим геном, ингибитором или супрессором.
Подавляемый ген носит название гипостатического.
ЭПИСТАЗ (расщепление по фенотипу)
F2 : 13 : 3
12 : 3 : 1
9 : 3 : 4
Различают доминантный и рецессивный эпистаз.
При доминантном эпистазе доминантный аллель одного гена (генсупрессор) подавляет действие доминантной или рецессивной аллели другого гена (гипостатический ген). Ген А (супрессор) подавляет ген В (гипостатический) или ген А (эпистатический) подавляет ген b (гипостатический). При доминантном эпистазе в дигибридном скрещивании происходит расщепление в соотношении 12:3:1 или 13:3.
♂ АС Ас аС ас
♀
АС
Ас
аС
ас
Э п и с т а з 13 : 3
Рис. 46.Расщепление 13:3 в F2 при доминантном эпистазе.
Например, при скрещивании 2 белых кур леггорн ААСС и виандотт аасс во втором поколении произойдёт расщепление 13 белых : 3 окрашенных.
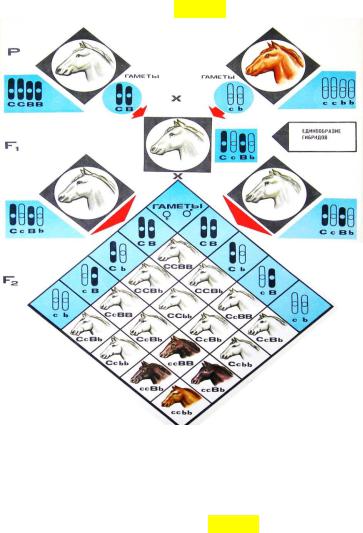
Ген С подавляет ген А. В отсутствии гена С проявляется ген А, и куры окрашены (рис. 46).
У лошадей ген В обусловливает синтез черного пигмента (вороная масть), а ген b – рыжую масть. Ген С – эпистатический ген (серая масть – результат эпистаза), а его рецессивный аллель с не подавляет проявлений гена B и b. Лошади серой масти имеют генотип – 9 •ѕ и 3 bbС•; вороные – 3В•сс, рыжие – 1bbсс. Расщепление 12 : 3 : 1 (рис. 47).
Рис. 47. Расщепление 12:3:1 в F2 при доминантном эпистазе.
В случае рецессивного эпистаза рецессивный эпистатический аллель в гомозиготном состоянии препятствует проявлению доминантной или рецессивной аллели другого гена (аа подавляет ген В или аа подавляет ген b). При рецессивном эпистазе в дигибридном скрещивании происходит расщепление в соотношении 9 : 3 : 4.
Примером рецессивного эпистаза у животных служит окраска шерсти у кроликов (рис. 48). Окраска «агути» у них определяется доминантным геном В, а чёрная окраска его рецессивным геном b. Оба признака проявляются только при наличии доминантного гена А, а его рецессивная аллель (а) подавляет цветность.
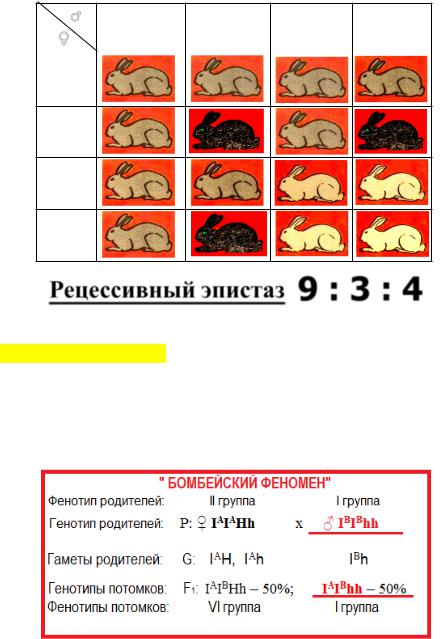
♂ |
АВ |
Аb аВ аb |
|
♀ |
|||
|
|
||
|
|
|
|
АВ |
|
|
Аb
аВ
аb
Рецессивный эпистаз 9 : 3 : 4
Рис. 48. Расщепление 9 : 3 : 4 в F2 при рецессивном эпистазе.
Ярким примером рецессивного эпистаза у человека является "Бомбейский феномен". Так, у человека группы крови АВО
контролируются тремя аллелями одного гена IA – вторая группа (синтез антигена А), IB – третья группа крови (синтез антигена В), i – первая группа кровит (синтез антигенов не происходит). В популяции встречается редкий мутантный аллель h независимого гена, который в гомозиготном состоянии подавляет действие аллелей А и В, что приводит к фенотипическому проявлению первой группы крови.
Рис. 49. Генотипы родителей и потомков с фенотипом крови Бомбей.
В Индии была описана семья, в которой родители имели вторую и первую группу крови. Оба родителя были гомозиготными по группам крови, и рождение ребенка с четвертой группой крови в их семье вызвало недоумение (рис. 49).
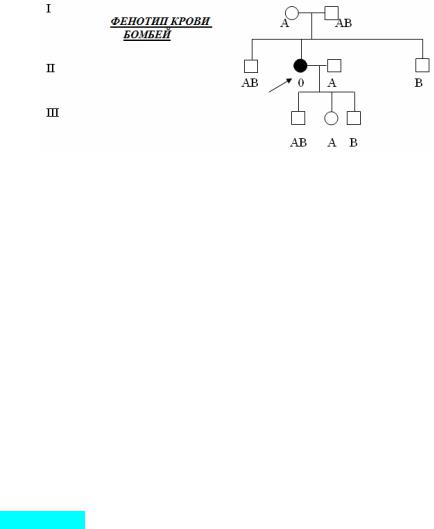
Таким образом, "Бомбейский феномен" определяется тем, что в генотипе детей есть ген подавляющий действие генов IА и IВ.
Рис. 50. Часть родословной женщины с фенотипом крови Бомбей. Генетически она имеет группу крови В, но из-за отсутствия антигенов АВ ее кровь отнесена к группе 0.
Группа крови АВ0 хорошо исследована биохимически. Известно, что антигены А и В представляют собой карбогидратные группы (углеводные остатки), связанные с жирными кислотами, молекулы которых выступают над поверхностью мембраны эритроцитов. Специфичность этих антигенов обусловлена концевым остатком карбогидратной цепи. Оба антигена происходят из одного и того же предшественника, или Н-вещества, к молекулам которого затем добавляется один или два концевых углеводных остатка сахара.
В крайне редких случаях, например, у женщин из Бомбея (Индия) имеется дефектное Н-вещество, которое не связывается с ферментом, добавляющим концевые углеводные остатки. У носителей группы крови 0 это приводит к фенотипу Бомбей. Оказалось, что этот дефект обусловлен рецессивной мутацией h в локусе, не гомологичном локусам А и В. Поэтому при наличии у человека аллелей IA и (или) IB вместе с генотипом hh антигены А и В в крови не обнаруживаются. Вот почему у одной из жительниц Бомбея с генотипом hh определили группу крови 0, хотя ее родители имели группу крови АВ, а сын был носителем аллеля IB (рис. 50). ПОЛИМЕРИЯ – явление, когда несколько неаллельных доминантных генов контролируют развитие одного и того же признака. Такие гены называются полимерными и обозначаются одной и той же буквой, но с другим индексом (например: А1, А2, А3, А4 и т.д.). Различают кумулятивную и некумулятивную полимерию.
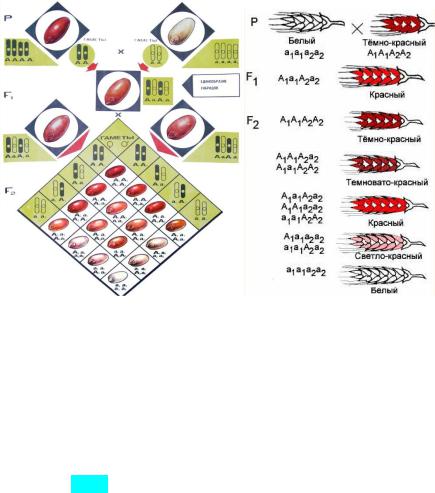
Рис. 51. Некумулятивная полимерия.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Впервые некумулятивная полимерия была описана в 1908 г. НильсономЭле при анализе наследования окраски эндосперма зерновок пшеницы (рис. 51). Доминантные неаллельные гены А1 и А2 обуславливают красную окраску зерен пшеницы, их рецессивные аллели а1 и а2 – отсутствие окраски. Действие генов А1 и А2 однозначно.
При наличии двух полимерных генов расщепление во втором поколении соответствует 15 : 1, при наличии трёх полимерных генов – 63 : 1 и т.д.). При кумулятивной полимерии действие полимерных генов суммируется, чем их больше, тем ярче выражен признак. Расщепление во втором поколении будет зависеть от количества полимерных генов (суммарный или аддитивный эффект генов), присутствующих в генотипе.
По типу кумулятивной полимерии наследуются многие количественные признаки (у человека: пигментация кожи, рост, вес, уровень интеллекта, скорость протекания биохимических реакций, артериальное давление, содержание сахара в крови; у животных: жирность молока; у растений: яровость, озимость, содержание витаминов, длина колоса у злаков, интенсивность окраски частей растения и др.).
Цвет кожи человека определяется взаимодействием нескольких пар генов по типу кумулятивной полимерии, т.е. цвет кожи тем темнее, чем больше доминантных генов в генотипе (рис. 52).
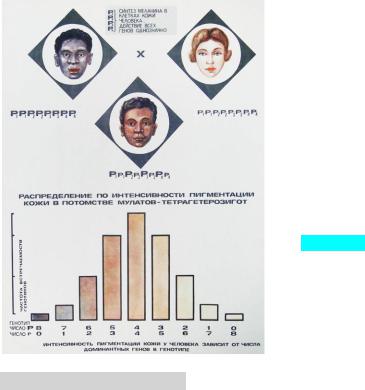
Возможные генотипы и фенотипы цвета кожи при наличии двух полимерных генов:
черная кожа – A1A1A2A2 темная – A1A1A2a2 смуглая (мулат) – A1a1A2a2 светлая – A1a1а2a2
белая – а1a1а2a2.
Расщепление по фенотипу в
F2 1: 4: 6: 4: 1
Рис. 52. Кумулятивная полимерия.
ЭФФЕКТ ПОЛОЖЕНИЯ ГЕНОВ – влияние расположения генов в хромосоме на проявление их активности. Явление открыто американским генетиком А. Стёртевантом в 1925 году. Наблюдается при структурных перестройках хромосом (транслокациях), в результате которых гены активных зон хромосом (эухроматина) могут переноситься в неактивные зоны (гетерохроматин) и инактивироваться и наоборот. При перестройке, возвращающей эухроматиновый ген из гетерохроматина в любую точку эухроматина, функционирование данного гена восстанавливается.
Эффект положения гена можно проследить при наследовании резус фактора (Rh-фактор).

Рис. 53. «Эффект положения».
В простейшем варианте резус-положительность доминирует над резусотрицательностью и практически наследование Rh-фактора иммитирует моногогенное наследование. Однако ряд исследований показывает, что система Rh определяется тремя антигенными факторами, которые детермируются тремя тесно сцепленными генами С, Д, Е, локализованными в коротком плече хромосомы 1. Внутри аллельных пар действует полное доминирование, а при взаимодействии между ними «эффект положения». Все это обусловливает разнообразие вариантов резус-антигенов и соответственно резус-несовместимости.
Основная роль в Rh-системе принадлежит антигену D. При его наличии на поверхности эритроцитов кровь является резус-положительной. Резусотрицательный фенотип формируется при отсутствии антигена D.
Однако антигены C и E оказывают влияние на резус-несовместимость. Суть «эффекта положения» в следующем: в рассмотренном примере (рис. 53) оба человека имеют одинаковые фенотипы и генотип. Они гетерозиготы по трем парам генов (CcDdEe) и оба резус-положительные, но в антигенном плане их кровь несовместима. У человека, у которого в одной хромосоме порядок расположения генов Cde, а в другой соответственно cDE, будут синтезироваться все три вида антигенов C, D, E и его кровь будет несовместима с кровью второго человека.
У второго человека порядок расположения генов внутри хромосомы cDe, а в гомологичной – CdE. Там, где в одной хромосоме расположены доминантные гены С и Е, ген Е выступает в роли супрессора по

отношению к гену С. Следовательно, фенотипическое проявление гена С в признак подавлено, антиген С не синтезируется.
35. Резус-фактор. Его фенотипическое проявление, закономерности наследования, эффект положения генов. Правила переливания крови с учетом Rh-принадлежности. Суть резус-конфликта между организмом матери и плода
Резус-фактор крови – антиген, представляющий собой особый вид протеина, присутствующего на оболочке красных кровяных телец человека и макаки-резус. Открыт белок был в 1940 году А. Винером и К.
Ландштейнером.
Этот белок есть у 85% жителей планеты европеоидной расы. Таких людей называют резус-положительными. Если же белка на оболочке эритроцитов нет – они называются резус-отрицательными. Среди людей монголоидной расы резус-отрицательных людей еще меньше – всего 1%, а среди жителей планеты негроидной расы только 7%.
Резус крови – это достаточно непростая конструкция, в которую входит не меньше сорока различных антигенов. Каждый из антигенов имеет свое «имя», которое прописывается с помощью букв и цифр. Наиболее распространенными антигенами являются Д, С, Е. Именно эти антигены наиболее активны.
|
Rh-пол. |
|
Rh-пол. |
|
P: |
♀ Rr |
x |
|
♂ Rr |
G: |
R, |
r |
|
R, r |
|
|
Rh-пол. |
|
Rh-отр. |
F1: |
RR; |
Rr; |
Rr; |
rr |
Резус крови передаётся по |
|
75% |
|
25% |
наследству как доминантный |
|
|
|
|
признак (R – ген наличия резус фактора, а r – отсутствие его) и не меняется в течение всей жизни.
Если у родителей присутствует резус (Rr, Rr), то у ребенка резус может присутствовать (RR, Rr), а может и отсутствовать (rr).
Если один из родителей резус-положителен (RR, Rr), а второй отрицателен (rr), – малыш может унаследовать как положительный резус (Rr), так и отрицательный (rr).
В том случае, если и у мамы и у папы резус отрицателен, ребенок обязательно также родится резус-отрицательным.
Обычно отрицательный резус-фактор никаких неприятностей его хозяину не приносит. Особого внимания и ухода требуют лишь резусотрицательные беременные женщины.
Если у будущей мамы резус отрицательный, а у будущего папы положительный, возникает опасность резус-конфликта. Но начаться он может лишь в том случае, если ребенок унаследует резус отца. Тогда его кровь будет плохо совместима с кровью матери (рис. 54).
Рис. 54. Резус-конфликт при беременности.
Во время беременности мать и плод едины, и, несмотря на то, что кровь их не смешивается, многие продукты обмена веществ да и отдельные клетки от плода поступают к матери, и наоборот. Этот обмен идет через структуры плаценты, через ту ее часть, которая называется плацентарным барьером.
Rh-кoнфликт возникает не ранее 7-8-й недели беременности, когда начинается формирование кроветворения у зародыша. Резусположительные эритроциты плода преодолевают плацентарный барьер и, попадая в кровь матери, а ее организм, воспринимаются как нечто чужеродное. Организм матери начинает вырабатывать защитные антитела. Защищая мать, эти антитела несут серьезную угрозу ее ребенку. При тяжелой форме резус-конфликта возможна внутриутробная гибель плода и выкидыш на любом сроке беременности.
Антитела матери проникают через плаценту и разрушают эритроциты ребенка. В крови появляется большое количество вещества, называемого билирубином. Билирубин окрашивает кожу малыша в желтый цвет.
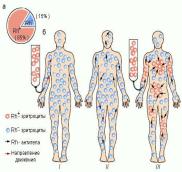
Поскольку эритроциты плода непрерывно уничтожаются, его печень и селезенка стараются ускорить выработку новых эритроцитов, увеличиваясь при этом в размерах. В конце концов, и они не справляются с восполнением убыли эритроцитов. Возникает состояние анемии (низкое содержание в крови эритроцитов, гемоглобина). Резус-конфликт может быть причиной поражения головного мозга ребенка, нарушения функции слуха и речи. В самых тяжелых случаях резус-конфликт проявляется врожденной водянкой (отеком) плода, которая может привести к его гибели.
В тяжелых случаях новорожденному может помочь заменное переливание крови. Ему вводят одногруппную резус-отрицательную кровь и проводят реанимационные мероприятия.
вырабатываться антитела (рис. 55), под действием которых наступает гемолиз эритроцитов донора (эритроциты, обладающие резусом, склеиваются в длинные цилиндры).
Для профилактики подобных осложнений необходимо переливать резусотрицательному реципиенту только резус-отрицательную кровь и перед переливанием производить пробу на совместимость не только по группам крови, но и по резусфактору.
формы изменчивости.
Изменчивость – это универсальное свойство живых организмов приобретать новые признаки в процессе индивидуального развития организмов (онтогенеза).
Как известно, эволюционная теория, разработанная Ж.Б. Ламарком (17441829), основывалась на ошибочном постулате о наследовании изменений, приобретаемых в течение жизни, т.е. о наследовании модификаций. Само по себе представление Ж.Б. Ламарка об эволюции
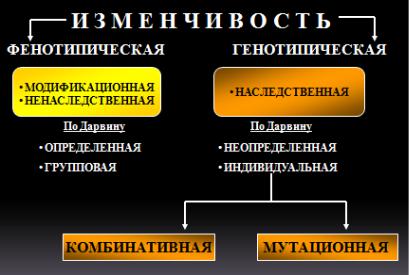
органических форм было, несомненно, прогрессивным для своего времени, но его объяснение механизма эволюционного прогресса было неверным и отражало распространенное заблуждение, характерное для биологов XVIII столетия.
Ч. Дарвин (1809-1882) в своем «Происхождении видов путем естественного отбора…» разделил изменчивость на определенную и неопределенную. Эта классификация в общем соответствует нынешнему делению изменчивости на ненаследственную и наследственную.
Различают два вида изменчивости: фенотипическую (модификационную) и генотипическую (рис. 3).
37.Мутации. Их классификация
Мутации можно объединять в группы – классифицировать по характеру проявления, по месту или по уровню их возникновения.
Классификация 1. Мутации по механизму возникновения (рис. 18). Причинами мутаций могут быть как естественные нарушения в метаболизме клеток (спонтанные мутации), так и действие различных факторов внешней среды (индуцированные мутации). Мутации в естественных условиях случаются редко – одна мутация определенного гена на 1 000 – 100 000 клеток.
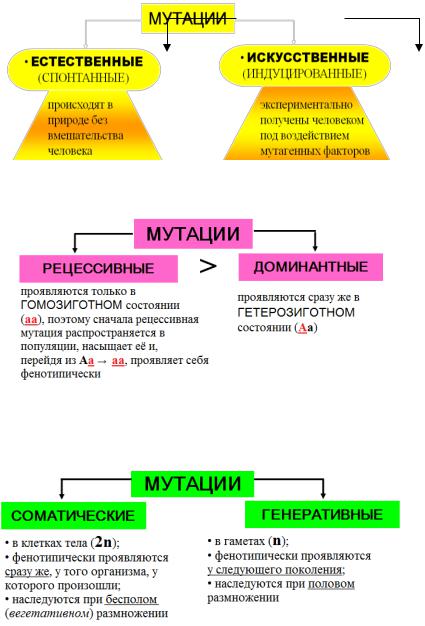
Рис. 18. Мутации по механизму возникновения.
Классификация 2. Мутации по характеру проявления (рис. 19).
Рис. 19. Мутации по характеру проявления.
Мутации чаще рецессивные, так как доминантные проявляются сразу же и легко "отбрасываются" отбором.
Классификация 3. Мутации по месту возникновения (рис. 20).
Рис. 20. Мутации по месту возникновения.
Мутация, возникшая в половых клетках, не влияет на признаки данного организма, а проявляется только в следующем поколении. Такие мутации называют генеративными. Если изменяются гены в соматических клетках, такие мутации проявляются у данного организма и затрагивают лишь
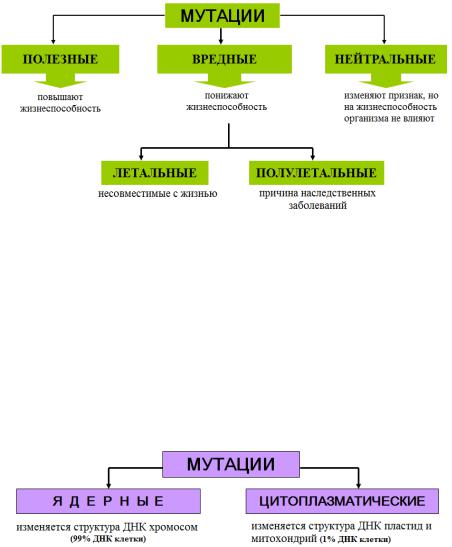
часть тела (глаза разного цвета), не передаются потомству при половом размножении. Но при бесполом размножении (вегетативном), если организм развивается из соматической клетки или группы клеток, имеющих изменившийся – мутировавший – ген, мутации могут передаваться потомству. Такие мутации называют соматическими.
Классификация 4. Мутации по адаптивному значению (рис. 21).
Рис. 21. Мутации по адаптивному значению.
Мутации бывают полезные, вредные и нейтральные. Полезные мутации – мутации, которые приводят к повышенной устойчивости организма (устойчивость тараканов к ядохимикатам), в конечном итоге, повышают приспособленность особей. Понятие «приспособленность» подразумевает жизнеспособность, плодовитость (фертильность) и конкурентоспособность особей. Вредные мутации нередко понижают жизнеспособность или плодовитость. Мутации, резко снижающие жизнеспособность, частично или полностью останавливающие развитие, называют полулетальными, а несовместимые с жизнью – летальными. Вредные мутации: глухота, дальтонизм. Нейтральные мутации никак не отражаются на жизнеспособности организма (цвет глаз, группа крови).
Классификация 5. Мутации по месту локализации в клетке (рис. 22).
Рис. 22. Мутации по месту локализации в клетке.
Мутации делятся на ядерные и внеядерные (или митохондриальные). Классификация 6. Мутации по характеру изменения фенотипа (рис. 23).
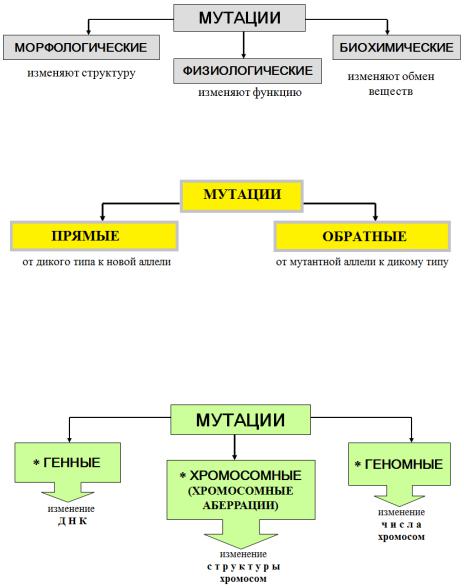
Рис. 23. Мутации по характеру изменения фенотипа.
Мутации могут быть биохимическими, физиологическими, анатомоморфологическими.
Классификация 7. Мутации по характеру изменения гена (рис. 24).
Рис. 24. Мутации по характеру изменения гена.
Мутации бывают прямые и обратные. Последние встречаются гораздо реже. Обычно прямая мутация связана с дефектом функции гена. Вероятность вторичной мутации в обратную сторону в той же точке очень мала, чаще мутируют другие гены.
Классификация 8. Мутации по уровню их возникновения или характеру изменений в генотипе (рис. 25).
Рис. 25. Мутации по характеру изменений в генотипе.
38.Генные мутации. Хромосомные мутации: аберрации, геномные мутации.
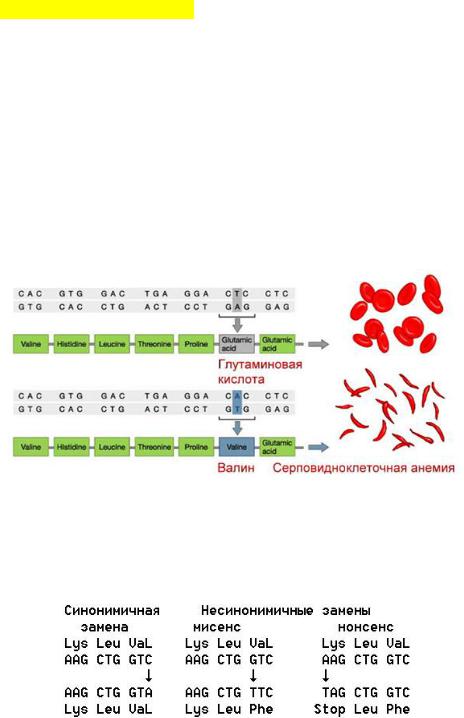
Генные мутации (точковые) представляют собой молекулярные, не видимые в световом микроскопе изменения структуры ДНК. К мутациям генов относятся любые изменения молекулярной структуры ДНК, независимо от их локализации и влияния на жизнеспособность. Некоторые мутации не оказывают никакого влияния на структуру и функцию соответствующего белка (синонимичная замена). Другая (большая) часть генных мутаций приводит к синтезу дефектного белка, не способного выполнять свойственную ему функцию (несинонимичная замена). Именно генные мутации обусловливают развитие большинства наследственных форм патологии.
Наиболее частыми моногенными заболеваниями являются; муковисцидоз, гемохроматоз, адреногенитальный синдром, фенилкетонурия, нейрофиброматоз, миопатии Дюшенна-Беккера, дальтонизм, гемофилия, серповидноклеточная анемия (рис. 26) и ряд других заболеваний.
Рис. 26. Механизм возникновения серповидноклеточной анемии.
Клинически они проявляются признаками нарушений обмена веществ (метаболизма) в организме. Мутация может заключаться:
1) в замене основания в кодоне, это так называемая миссенс-мутация (от англ. “mis” – ложный, неправильный + лат. “sensus” – смысл) – замена нуклеотида в кодирующей части гена, приводящая к замене аминокислоты в полипептиде (рис. 27);
Рис. 27. Мисенс и нонсенс мутации.
2)в таком изменении кодонов, которое приведет к остановке считывания информации, это так называемая нонсенс-мутация (от лат. “non” – нет + “sensus” – смысл) – замена нуклеотида в кодирующей части гена, приводит к образованию кодона-терминатора (стоп-кодона) и прекращению трансляции (рис. 27);
3)в нарушении считывания информации, сдвиге рамки считывания, называемом фреймшифтом (от англ. “frame” – рамка + “shift” – сдвиг, перемещение), когда молекулярные изменения ДНК приводят к изменению триплетов в процессе трансляции полипептидной цепи;
4)в замене нуклеотидов (рис. 28): транзиции – мутация, обусловленная заменой одного пуринового основания на другое или пиримидинового на другое пиримидиновое; трансверсия (от лат. “transversus” – повернутый в сторону, отведенный) – мутация, обусловленная заменой пуринового основания (аденин, тимин) на пиримидиновое (гуанин, цитозин) и наоборот. В отличие от транзиций, трансверсии иногда называют сложными или перекрестными заменами, т. к. происходит изменение ориентации пурин – пиримидин в мутантном сайте двуцепочечной молекулы нуклеиновой кислоты.
Рис. 28. Типы замен нуклеотидов.
Известны и другие типы генных мутаций.
По типу молекулярных изменений выделяют:
1)делеции (от лат. “deletio” – уничтожение), когда происходит утрата сегмента ДНК размером от одного нуклеотида до гена;
2)дупликации (от лат. “duplicatio” – удвоение), т.е. удвоение или повторное дублирование сегмента ДНК от одного нуклеотида до целых генов;
3)инверсии (от лат. “inversio” – перевертывание), т. е. поворот на 180° сегмента ДНК размерами от двух нуклеотидов до фрагмента, включающего несколько генов;
4)инсерции (от лат. “insertion” – прикрепление), т.е. вставка фрагментов ДНК размером от одного нуклеотида до целого гена.
Молекулярные изменения, затрагивающие от одного до нескольких нуклеотидов, рассматривают как точечную мутацию.
Принципиальным и отличительным для генной мутации является то, что она приводит к изменению генетической информации и может передаваться от поколения к поколению.
Определенная часть генных мутаций может быть отнесена к нейтральным мутациям, поскольку они не приводят к каким-либо изменениям фенотипа. Например, за счет вырожденности генетического кода одну и ту же аминокислоту могут кодировать два триплета, различающихся только по одному основанию. С другой стороны, один и тот же ген может изменяться (мутировать) в несколько различающихся состояний.
Например, ген, контролирующий группу крови системы АВ0, имеет три аллеля: 0, А и В, сочетания которых определяют 4 группы крови. Группа крови системы АВ0 является классическим примером генетической изменчивости нормальных признаков человека.
Именно генные мутации обусловливают развитие большинства наследственных форм патологии. Болезни, обусловленные подобными мутациями, называют генными, или моногенными болезнями, т.е.
заболеваниями, развитие которых детерминируется мутацией одного гена.
Таким образом, генные мутации приводят к появлению новых признаков.
Рис. 29. Виды хромосомных мутаций.
Хромосомные мутации (хромосомные аберрации) – это структурные изменения отдельных хромосом, как правило, видимые в световом микроскопе. В хромосомную мутацию вовлекается большое число (от десятков до нескольких сотен) генов, что приводит к изменению генотипа. Причиной этого может быть разрыв хромосомы на фрагменты и восстановление её в новых сочетаниях.
Несмотря на то, что хромосомные аберрации, как правило, не изменяют последовательность ДНК в специфических генах, изменение числа копий генов в геноме приводит к генетическому дисбалансу вследствие недостатка или избытка генетического материала. Различают две большие группы хромосомных мутаций: внутрихромосомные и межхромосомные (рис. 29).
Внутрихромосомные мутации – аберрации в пределах одной хромосомы. К ним относятся:
- делеции (от лат. “deletio” – уничтожение) – утрата одного из участков хромосомы, внутреннего или терминального (рис. 30).
Рис. 30. Схематическое изображение делеции.
А – нормальная хромосома, Б – делетированная хромосома.
Это может обусловить нарушение эмбриогенеза и формирование множественных аномалий развития (например, делеция в регионе короткого плеча 5-й хромосомы, обозначаемая как 5р-, приводит к недоразвитию гортани, порокам сердца, отставанию умственного
развития). Этот симптомокомплекс известен как синдром Лежена или синдром "кошачьего крика", поскольку у больных детей из-за аномалии гортани плач напоминает кошачье мяуканье (рис. 31);
А Б
Рис. 31. А – дети с синдромом Лежена в возрасте 8 месяцев, 2, 4 и 8 лет соответственно; Б – хромосомный набор больного с синдромом Лежена: групповая (от А до G) и индивидуальная идентификация хромосом (стрелкой указан дефект короткого плеча хромосомы 5-й пары, вторая хромосома не изменена).
-дупликации (от лат. “duplcatio” – |
|
||
удвоение) – удвоение (или |
|
||
умножение) какого-либо участка |
|
||
хромосомы (рис. 32). Например, |
|
||
дупликация |
по |
одному из |
|
коротких плеч 9-й хромосомы |
|
||
обусловливает |
множественные |
|
|
пороки, включая микроцефалию, |
|
||
задержку |
|
физического, |
|
психического |
|
и |
Рис. 32. Схематическое |
интеллектуального развития. |
изображение дупликации. А – |
||
|
|
|
нормальная хромосома, Б – |
|
|
|
дуплицированная хромосома. |
-инверсии (от лат. “inversio” – |
встраивается на прежнее место |
||
перевертывание). В результате |
после поворота на 180°. В |
||
двух точек разрывов хромосомы |
результате нарушается только |
||
образовавшийся |
фрагмент |
|
|
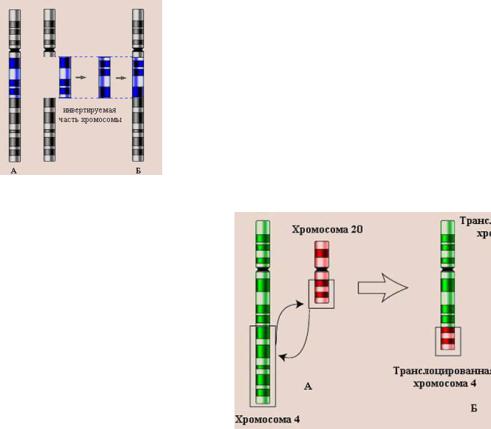
порядок расположения генов |
А – нормальная хромосома, Б – |
(рис. 33); |
инвертированная хромосома. |
Рис. 33. Схематическое |
|
|||
изображение инверсии. |
|
|||
Межхромосомные мутации, или |
|
|||
мутации |
перестройки |
– обмен |
|
|
фрагментами |
|
между |
|
|
негомологичными хромосомами. |
|
|||
Такие |
мутации |
получили |
|
|
название транслокации (от лат. |
|
|||
“trans” – за, через + “locus” – |
|
|||
место) |
|
(рис. |
34). |
|
Рис. |
34. |
Схематическое |
|
|
изображение |
транслокации |
|
||
хромосом А |
– нормальные |
|
||
хромосомы, |
Б |
– |
. |
|
транслоцированные хромосомы
.
Выделяют несколько разновидностей транслокаций. Это:
-реципрокная транслокация, когда две хромосомы обмениваются своими фрагментами;
-нереципрокная транслокация, когда фрагмент одной хромосомы транспортируется на другую. Возможно присоединение фрагмента к своей же хромосоме, но в новом месте – транспозиция (рис. 35);
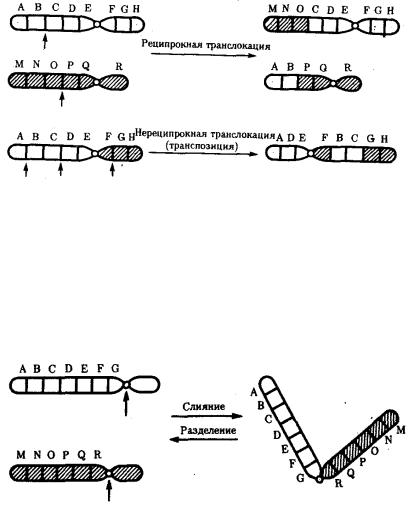
Рис. 35. Реципрокная и нереципрокная транслокация.
- "центрическое" слияние (робертсоновская транслокация) –
соединение двух акроцентрических хромосом в районе их центромер с потерей коротких плеч (рис. 36). При таких мутациях не только появляются хромосомы с новой морфологией, но и изменяется их количество в кариотипе.
При поперечном разрыве хроматид через центромеры "сестринские" хроматиды становятся "зеркальными" плечами двух разных хромосом, содержащих одинаковые наборы генов. Такие хромосомы называют
изохромосомами.
Рис. 36. Робертсоновская транслокация.
Как внутрихромосомные (делеции, инверсии и дупликации), так и межхромосомные (транслокации) аберрации и изохромосомы связаны с физическими изменениями структуры хромосом, в том числе с механическими разломами.
Описанные структурные изменения хромосом, как правило, сопровождаются изменением генетической программы, получаемой клетками нового поколения после деления материнской клетки, так как 1) изменяется количественное соотношение генов (при делециях и дупликациях),
2) меняется характер их функционирования в связи с изменением взаимного расположения в хромосоме (при инверсии и транспозиции) или с переходом в другую группу сцепления (при транслокации).
Чаще всего такие структурные изменения хромосом отрицательно сказываются на жизнеспособности отдельных соматических клеток организма, но особенно серьезные последствия имеют хромосомные перестройки, происходящие в предшественниках гамет.
Значение хромосомных аберраций. Однако, несмотря на неблагоприятные, как правило, последствия хромосомных мутаций, иногда они оказываются совместимыми с жизнью клетки и организма и обеспечивают возможность эволюции структуры хромосом, лежащей в основе биологической эволюции.
Так, небольшие по размеру делеции могут сохраняться в гетерозиготном состоянии в ряду поколений. Менее вредными, чем делеции, являются дупликации, хотя большой объем материала в увеличенной дозе (более 10% генома) приводит к гибели организма.
Нередко жизнеспособными оказываются робертсоновские транслокации, часто не связанные с изменением объема наследственного материала. Этим можно объяснить варьирование числа хромосом в клетках организмов близкородственных видов. Например, у разных видов дрозофилы количество хромосом в гаплоидном наборе колеблется от 3 до 6, что объясняется процессами слияния и разделения хромосом.
Возможно, |
|
|
существенным |
двух обезьяньих хромосом (рис |
|||||
моментом |
в |
появлении |
вида |
37). |
|
|
|
||
Homo sapiens были структурные |
К существенному варьированию |
||||||||
изменения |
|
хромосом |
у |
его |
морфологии |
|
хромосом, |
||
обезьяноподобного |
предка. |
лежащему |
в |
основе |
их |
||||
Установлено, что два плеча |
эволюции, |
|
приводят |
||||||
крупной |
второй |
хромосомы |
транслокации, транспозиции |
и |
|||||
человека |
соответствуют |
двум |
инверсии. |
Анализ хромосом |
|||||
разным |
|
|
хромосомам |
человека показал, что его 4, 5, 12 |
|||||
современных человекообразных |
и 17-я хромосомы отличаются от |
||||||||
обезьян (12-й и 13-й – шимпанзе, |
соответствующих |
хромосом |
|||||||
13-й и-14-й – гориллы и |
шимпанзе |
перицентрическими |
|||||||
орангутана). |
Вероятно, |
эта |
инверсиями. |
|
|
|
|||
человеческая |
|
хромосома |
|
|
|
|
|||
образовалась |
в |
результате |
|
|
|
|
|||
центрического слияния |
по |
типу |
|
|
|
|
|||
робертсоновской |
транслокации |
|
|
|
|
||||

Рис. 37. Плечи 2-ой пары хромосом человека соответствуют 12 и 13 хромосомам шимпанзе.
Таким образом, изменения хромосомной организации, чаще всего оказывающие неблагоприятное воздействие на жизнеспособность клетки и организма, с определенной вероятностью могут быть перспективными, наследоваться в ряду поколений клеток и организмов и создавать предпосылки для эволюции хромосомной организации наследственного материала.
Геномные мутации приводят к изменению числа хромосом. К геномным мутациям относятся анеуплоидии и изменение плоидности структурно неизмененных хромосом. Выявляются цитогенетическими методами.
Анеуплоидия – изменение числа хромосом в диплоидном наборе, некратное гаплоидному (2n + 1 → трисомия (увеличение), 2n ─ 1→ моносомия (уменьшение)).
Полиплоидия – увеличение числа наборов хромосом, кратное гаплоидному (2n + n).

Частным случаем геномных мутаций является полиплоидия, т. е. кратное увеличение числа хромосом в клетках в результате нарушения их расхождения в митозе или мейозе. Соматические клетки таких организмов содержат 3n, 4n, 8n и т. п. хромосом в зависимости от того, сколько хромосом было в гаметах, образовавших этот организм. Полиплоидия часто встречается у бактерий и растений, но очень редко – у животных. Полиплоидны три четверти всех культивируемых человеком злаков. Если гаплоидный набор хромосом (n) для пшеницы равен 7, то основной сорт, разводимый в наших условиях, – мягкая пшеница – имеет по 42 хромосомы, т. е. 6n.
Рис. 38. Полиплоиды капусты.
Полиплоидами являются окультуренная свекла, гречиха, капуста (рис. 38) и т. п. Как правило, растения-полиплоиды имеют повышенные жизнеспособность, размеры, плодовитость и т. п.
В настоящее время разработаны специальные методы получения полиплоидов. Например, растительный яд из безвременника осеннего – колхицин – способен разрушать веретено деления (рис. 39) при образовании гамет, в результате чего получаются гаметы, содержащие по 2n хромосом. При слиянии таких гамет в зиготе окажется 4n хромосом.
Рис. 39. Формирование веретена деления во время деления клетки.
Геномные мутации в животном и растительном мире многообразны, но у человека обнаружены только 3 типа геномных мутаций: тетраплоидия, триплоидия и анеуплоидия. При этом из всех вариантов анеуплоидий встречаются только трисомии по аутосомам, полисомии по половым хромосомам (три-, тетра- и пентасомии), а из моносомий встречаются только моносомия-Х.
У человека полиплоидия, а также большинство анеуплоидий являются летальными мутациями.
К наиболее частым геномным мутациям относятся:
трисомия – наличие трех гомологичных хромосом в кариотипе (например, по 21-й паре при синдроме Дауна, по 18-й паре при синдроме Эдвардса, по 13-й паре при синдроме Патау; по половым хромосомам: XXX, XXY, XYY);
|
моносомия – наличие только одной из двух |
гомологичных |
хромосом. При моносомии по любой из аутосом |
нормальное развитие эмбриона невозможно. Единственная моносомия у человека, совместимая с жизнью, – моносомия по X-хромосоме – приводит к синдрому Шерешевского-Тернера (45, Х0).
Причиной, приводящей к анеуплоидии, является нерасхождение хромосом во время клеточного деления при образовании половых клеток или утрата хромосом в результате анафазного отставания, когда во время движения к полюсу одна из гомологичных хромосом может отстать от всех других негомологичных хромосом. Термин "нерасхождение" означает отсутствие разделения хромосом или хроматид в мейозе или митозе. Утрата хромосом может приводить к мозаицизму, при котором имеется одна эуплоидная (нopмальная) клеточная линия, а другая – моносомная.
Нерасхождение хромосом наиболее часто наблюдается во время мейоза (рис. 40). Хромосомы, которые в норме должны делиться во время мейоза, остаются соединенными вместе и в анафазе отходят к одному полюсу клетки. Таким образом, возникают две гаметы, одна из которых имеет добавочную хромосому, а другая не имеет этой хромосомы. При оплодотворении гаметы с нормальным набором хромосом гаметой с лишней хромосомой возникает трисомия (т.е. в клетке присутствуют три гомологичные хромосомы), при оплодотворении гаметой без одной хромосомы возникает зигота с моносомией. Если моносомная зигота образуется по какой-либо аутосомной (не половой) хромосоме, то развитие организма прекращается на самых ранних стадиях развития.
Рис. 40. Схема нерасхождения хромосом в гаметогенезе и результаты оплодотворения.
Летальный или дизморфогенетический эффект хромосомных аномалий обнаруживается на всех стадиях внутриутробного онтогенеза (имплантация, эмбриогенез, органогенез, рост и развитие плода). Суммарный вклад хромосомных аномалий во внутриутробную гибель (после имплантации) у человека составляет 45 %. При этом, чем раньше прерывается беременность, тем вероятнее, что это обусловлено аномалиями развития эмбриона, вызванными хромосомным дисбалансом. У 2-4–недельных абортусов (эмбрион и его оболочки) хромосомные аномалии обнаруживают в 60-70 % случаев. В 1 триместре беременности хромосомные аномалии встречаются у 50 % абортусов. У плодов-выкидышей 2 триместра такие аномалии находят в 25-30 % случаев, а у плодов, погибших после 20 недели беременности, в 7 % случаев.
Наиболее тяжелые формы по дисбалансу хромосомного набора встречаются у ранних абортусов. Это полиплоидии (25 %), полные трисомии по аутосомам (50 %). Трисомии по некоторым аутосомам (1; 5; 6; 11; 19) встречаются крайне редко даже у элиминированных эмбрионов и плодов, что свидетельствует о большой морфогенетической значимости этих аутосом. Данные аномалии прерывают развитие в доимплантационном периоде или нарушают гаметогенез.
Высокая морфогенетическая значимость аутосом еще более отчетливо выражена при полных аутосомных моносомиях. Последние редко обнаруживаются даже в материале ранних спонтанных абортов из-за раннего летального эффекта такого дисбаланса.
39. Мутагены среды. Последствия мутаций для человека. Антимутационные барьеры.
В основе этой изменчивости лежат мутации. Основная причина возникновения новых признаков и свойств у живых организмов – это
проявление мутаций. Мутации – это изменения генотипа, происходящие под влиянием факторов внешней или внутренней среды. Мутации имеют принципиальные отличия от модификаций (табл. 3).
Таблица 3
Сравнение модификаций и мутаций
Впервые термин «мутация» был предложен в 1901 г. голландским ученым Гуго де Фризом, описавшим самопроизвольные мутации у растений и создавший мутационную теорию.
Основные положения мутационной теории
1.Мутации возникают внезапно, без всяких переходов. Чаще бывают рецессивными, реже – доминантными.
2.Вызываются внешними и внутренними факторами.
3.Мутации стойко передаются из поколения в поколение, наследственны.
4.Это качественные изменения, которые, как правило, не образуют непрерывного ряда вокруг средней величины признака.
5.Мутировать может любая часть организма, т.е. мутации возникают в разных направлениях, они не направленны. Мутации могут быть вредными, полезными и нейтральными.
6.Успех в выявлении мутаций зависит от числа проанализированных особей.
7.Одни и те же мутации могут возникать повторно.
8.Мутации являются и элементарным эволюционным материалом, и ненаправляющим элементарным эволюционным фактором.
9.Мутационный процесс – источник резерва наследственной изменчивости популяций.
Мутации появляются редко, но приводят к внезапным скачкообразным изменениям признаков, которые передаются из поколения в поколение. Мутационный процесс идет постоянно, с накоплением мутаций в генотипах. А если учесть, что число генов в организме велико, то можно сказать, что в генотипах всех живых организмов имеется значительное число генных мутаций. Мутации – это крупнейший биологический фактор, обусловливающий огромную наследственную изменчивость организмов, что дает материал для эволюции.
Мутациями называются внезапные, стойкие, ненаправленные, скачкообразные изменения хромосом или единиц наследственности – генов, влекущие за собой изменения наследственных признаков. Мутации обязательно вызывают изменения генотипа, которые наследуются потомством и не связаны со скрещиванием и рекомбинацией генов. Сходство между комбинативной и мутационной изменчивостью заключается в том, что в обоих случаях потомство получает набор генов каждого из родителей.
Мутационная изменчивость является одним из главных факторов эволюционного процесса. В результате мутаций могут возникать полезные признаки, которые под действием естественного отбора дадут начало новым видам и подвидам. Подавляющее число мутаций неблагоприятно или даже смертельно для организма, так как они разрушают отрегулированный на протяжении миллионов лет естественным отбором целостный генотип.
Способностью к мутированию обладают все живые организмы. У каждой конкретной мутации есть какая-то причина, хотя в большинстве случаев она неизвестна. Однако общее количество мутаций можно резко увеличить, используя различные способы воздействия на организм.
Факторы, вызывающие мутации, получили название мутагенных факторов (рис. 17).
Рис. 17. Классификация мутагенных факторов.
Во-первых, сильнейшим мутагенным действием обладает ионизирующее излучение. Радиация увеличивает число мутаций в сотни раз. Во-вторых, мутации вызывают вещества, которые действуют, например, на ДНК, разрывая цепочку нуклеотидов. Есть вещества, действующие и на другие молекулы, но также дающие мутации. Например, колхицин, приводящий к одному из видов мутаций – полиплоидии. В-третьих, к мутациям приводят и различные физические воздействия, например, повышение температуры окружающей среды.
40.Комбинативная изменчивость. Ее источники, значение. Системы браков в популяциях человека.
Комбинативная изменчивость основывается на возникновении новых комбинаций генов родителей. При комбинативной изменчивости в результате слияния родительских гамет возникают новые комбинации генов, однако сами гены и хромосомы остаются неизменными (пример, каждый новый организм является новой комбинацией генов родителей).
Механизмы комбинативной изменчивости:
1)кроссинговер в профазу-І мейоза (рис. 14);
2)независимое расхождение гомологичных хромосом в анафазу-І первого мейотического деления;
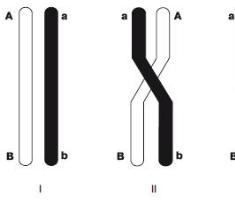
3)независимое расхождение хроматид (дочерних хромосом) в анафазу-ІІ второго мейотического деления;
4)случайное сочетание гамет в момент оплодотворения;
5)случайный подбор родительских пар.
Рис. 14. Упрощенная схема кроссинговера.
Две гомологичные хромосомы разрываются в точке контакта (II), и участки
их воссоединяются в ином сочетании, вследствие чего образуются две хромосомы (III), каждая из которых содержит участки обоих исходных хромосом (I).
В основе комбинативной изменчивости лежит половое размножение организмов, вследствие которого возникает огромное разнообразие генотипов.
Независимое расхождение гомологичных хромосом в первом мейотическом делении. Именно независимое комбинирование хромосом при мейозе является основой третьего закона Менделя. Появление зеленых гладких и желтых морщинистых семян гороха во втором поколении от скрещивания растений с желтыми гладкими и зелеными морщинистыми семенами – пример комбинативной изменчивости.
Взаимный обмен участками гомологичных хромосом, или кроссинговер. Он создает новые группы сцепления, т.е. служит важным источником генетической рекомбинации аллелей. Рекомбинантные хромосомы, оказавшись в зиготе, способствуют появлению признаков, нетипичных для каждого из родителей.
Эти источники комбинативной изменчивости действуют независимо и одновременно, обеспечивая при этом постоянную «перетасовку» генов, что приводит к появлению организмов с другими генотипом и фенотипом. Однако новые комбинации генов довольно легко распадаются при передаче из поколения в поколение.
Любая особь генетически уникальна (за исключением однояйцевых близнецов и особей, возникших за счет бесполого размножения клона, имеющего родоначальником одну клетку). Так, если допустить, что в каждой паре гомологичных хромосом имеется только одна пара

аллельных генов, то для человека, у которого гаплоидный набор хромосом равен 23, число возможных генотипов составит 2 в 23 степени. Такое огромное количество генотипов в 20 раз превышает численность всех людей на Земле. Однако в действительности гомологичные хромосомы различаются по нескольким генам и в расчете не учтено явление кроссинговера. Поэтому количество возможных генотипов выражается астрономическим числом, и можно с уверенностью утверждать, что возникновение двух одинаковых людей практически невероятно.
Комбинативная изменчивость является важнейшим источником всего колоссального наследственного разнообразия, характерного для живых организмов. Однако перечисленные источники изменчивости не порождают существенных для выживания стабильных изменений в генотипе, которые необходимы, согласно эволюционной теории, для возникновения новых видов. Такие изменения возникают в результате мутаций.
Благодаря комбинативной изменчивости создаётся разнообразие генотипов в потомстве, что имеет значение для эволюционного процесса в связи с тем, что:
увеличивается разнообразие материала для эволюционного процесса без снижения жизнеспособности особей;
расширяются возможности приспособления организмов к изменяющимся условиям среды и тем самым обеспечивается выживание группы организмов (популяции, вида) в целом.
Рис. 15. Гетерозис по продуктивности гибрида (в центре), полученного от скрещивания двух различных линий кукурузы (справа и слева).
Комбинативная изменчивость используется в селекции с целью получения более ценного в хозяйственном отношении сочетания наследственных признаков. В частности, применяется явление гетерозиса,
повышения жизнеспособности, интенсивности роста и других показателей при гибридизации между представителями различных подвидов или сортов. Ярко выражено оно, например, у кукурузы (рис. 15), обусловливая значительный экономический эффект.
Противоположный эффект даёт явление инбридинга или близкородственного скрещивания – скрещивания организмов, имеющих общих предков. Общность происхождения скрещиваемых организмов увеличивает у них вероятность наличия одних и тех же аллелей любых генов, а, следовательно, – вероятность появления гомозиготных организмов. Наибольшая степень инбридинга достигается при самоопылении у растений и самооплодотворении у животных. Гомозиготность увеличивает возможность проявления рецессивных аллельных генов, мутагенные изменения которых приводят к появлению организмов с наследственными аномалиями.
Результаты изучения явления комбинативной изменчивости используются в медико-генетическом консультировании, особенно на его втором и третьем этапах: прогноз потомства, формирование заключения и объяснение смысла генетического риска. В консультировании будущих супружеских пар используется установление вероятности наличия у каждого из двух индивидуумов аллелей, полученных от общего предка и идентичных по происхождению. Для этого используют коэффициент родства, выражаемый в долях единицы. У монозиготных близнецов он равен 1, у родителей и детей, братьев и сестёр – 1/2, у деда и внука, дяди и племянника – 1/4, у двоюродных сибсов (братьев и сестёр) – 1/8, у троюродных сибсов – 1/32 и т.д.
Подбор родительских пар в популяции людей осуществляется через систему браков.
Системы браков
В человеческих популяциях комбинативная изменчивость реализуется через системы браков.
Панмиксические браки происходят в больших по численности популяции людей, где осуществляется свободный подбор супружеских пар.
Панмикси́я – свободное скрещивание разнополых особей в популяции, при котором достигается случайное комбинирование гамет и равновесное распределение частот гомо- и гетерозиготных особей. Полная панмиксия возможна лишь в идеальных популяциях. Согласно
закону Харди-Вайнберга (1908 г.) в идеальных популяциях частота
аллелей и генотипов будет оставаться постоянной из поколения в поколение при выполнении следующих условий:
1)численность особей популяции достаточно велика (в идеале – бесконечно велика),
2)спаривание происходит случайным образом (т. е. осуществляется панмиксия),
3)мутационный процесс отсутствует,
4)отсутствует обмен генами с другими популяциями (полная изоляция),
5)естественный отбор отсутствует, т. е. особи с разными генотипами одинаково плодовиты и жизнеспособны.
Строгой панмиксии в популяциях людей не существует. Предпочтения в выборе брачного партнёра и социальные факторы препятствуют подлинно случайному заключению браков. Тем не менее, существуют гены (например, определяющие группы крови), которые обычно редко принимаются во внимание при подборе брачных пар и распределяются среди жителей популяции так, как если бы подбор брачных пар происходил более или менее случайно.
Инбридинг – кровнородственные браки. Инбридинг приводит к
гомозиготизации генотипов и снижению адаптивной ценности, т.е. приспособленности. Это кровнородственные браки внутри изолятов или кровнородственные браки в большой популяции из-за социальной изоляции по национальным или религиозным признакам (например, евреи). У евреев, в силу изоляции и национально-религиозных особенностей, редко проходило заражение сифилисом, поэтому у них это заболевание протекает особенно тяжело, с высокой частотой прогрессивных параличей. Однако другое заболевание (болезнь ТеяСакса), напротив, намного чаще регистрируется в еврейских семьях, особенно, среди евреев ашкенази.
Инцест – запретные кровнородственные браки (1 или 2 степень родства). Генетические эффекты у потомства: увеличение частоты мертворождений и врожденных уродств, ранней смертности и наследственных болезней.
С самого начала существования человеческого рода есть табуирование, запрет на кровосмешение в пределах семьи первого колена. Табу на инцест существует только в мире людей, причем еще с древних времен. Так, например, в племени зулусов отец, который насиловал дочь, должен был быть обезглавлен. По библейским, Моисеевым традициям родители,
которые замечены в сексуальной связи с детьми, подвергаются остракизму, то есть отлучению от церкви.
Аутбридинг – браки неродственных людей в больших популяциях или браки далеко неродственных людей из разных больших и малых рас. Они увеличивают степень гетерозиготности.
Генетические эффекты у потомства: снижение частот рецессивных заболеваний, гетерозис – гибридная сила.
Ассортативные браки (фр. "assorti" – подобранный) – «избирательные браки». Самая часто встречающаяся форма брака в популяции людей.
Ассортативность – предпочтительность в выборе фенотипически сходного брачного партнера (положительная ассортативность) или брак между фенотипически различными лицами (отрицательная ассортативность), приводящая к отклонению от панмиксии, то есть случайного относительно фенотипических признаков и равновероятного вступления в брак.
Примером отрицательных ассортативных браков является заключение союза между двумя абсолютно рыжими людьми. В данном случае вероятность образования пары между фенотипически похожими особями ниже ожидаемой. Если вероятность образования пар между похожими особями выше, чем ожидается, речь идет о положительных
ассортативных браках.
Ассортативные браки между людьми со сходным фенотипом по генетическому эффекту приравниваются к кровнородственным бракам. Практически это служит причиной того, что частота ряда психических заболеваний и состояний (шизофрения, неврозы, алкоголизм, олигофрения) выше среди брачных партнеров – больных с соответствующими нозологическими формами, чем можно было бы ожидать, исходя из их популяционной частоты. Формируются, таким образом, субпопуляции с высокой генотипической корреляцией внутри них и повышенным риском заболеваемости у потомства. Так, пациенты с дебильностью вступают в брак обычно с равными между собой по интеллекту и нередко имеют значительно больше детей, в основном, повидимому, также дебилов, чем ответственные за своих детей и обладающие нормальным интеллектом индивиды. Приводятся также сведения о том, что пациенты с шизофренией объединяются в браки в 4 раза чаще, нежели это происходило бы случайно.
Таким образом, ассортативные браки ведут к изменению частоты некоторых генов в популяции, что может привести либо к ухудшению,
либо к улучшению жизнеспособности данной популяции в зависимости от направления предпочтений.
Закон гомологических рядов наследственной изменчивости
В селекции важное значение имеет закон гомологических рядов наследственной изменчивости, сформулированный советским ученым
Н.И. Вавиловым. Он гласит: «Генетически близкие роды и виды
характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других родственных видов и родов».
Значение закона: зная характер наследственных изменений у одних сортов, можно предвидеть сходные изменения у родственных видов и, воздействуя на них мутагенами, вызывать у них подобные полезные изменения, что значительно облегчает получение хозяйственно ценных форм.
Н. И. Вавилов исследовал изменчивость признаков у растений из семейства злаковых (табл. 2). Из 38 признаков, которые характерны для различных видов этого семейства (окраска колосковых чешуй и зерна, остистость и безостость, форма зерна, строение листьев, окраска всходов, озимость и яровость, холодостойкость и т. д.), у ржи Н. И. Вавилов обнаружил 37 признаков, у пшеницы – 37, у овса и ячменя – по 35, у кукурузы и риса – по 32 и т. д.
Исходно закон касался изменчивости у растений, однако Н. И. Вавилов указывал на применимость его и к животным. Известны многие примеры гомологической изменчивости и у человека. Например, альбинизм (дефект синтеза клетками красящего вещества) обнаружен у европейцев, негров и индейцев; среди млекопитающих – у грызунов, хищных, приматов (рис. 16).

Рис. 16. Альбинизм у человека и животных.
Малорослые темнокожие люди – пигмеи – встречаются в тропических лесах экваториальной Африки, на Филиппинских островах и в джунглях полуострова Малакки; некоторые наследственные дефекты и уродства, присущие человеку, отмечены и у животных. Таких животных используют в качестве модели для изучения аналогичных дефектов у человека.
Например, катаракта глаза бывает у мыши, крысы, собаки, лошади; гемофилия – у мыши и кошки, диабет – у крысы; врожденная глухота – у морской свинки, мыши, собаки; заячья губа – у мыши, собаки, свиньи и т.д. Эти наследственные дефекты – убедительное подтверждение закона гомологических рядов наследственной изменчивости Н.И. Вавилова.
41.Модификационная изменчивость. Норма реакции. Фенокопии.
Ненаследственная (фенотипическая, модификационная) изменчивость – это фенотипические различия у генетически одинаковых особей, способность организмов изменять свой фенотип под влиянием различных факторов. Фенотипическая изменчивость не связана с изменением генетического материала. Она является ответной реакцией организма на конкретные изменения окружающей среды. Большую роль в формировании признаков организмов играет среда его обитания. Каждый организм развивается и обитает в определенной среде, испытывая на себе действие ее факторов, способных изменять морфологические и физиологические свойства организмов, т.е. их фенотип.
Причиной модификационной изменчивости являются изменения внешней среды обитания организма или его внутренней среды, не затрагивающие гены организма, наследственный материал и не вызывающие изменений генотипа особи. Следовательно, эти изменения не наследуются (ненаследственная). Так, если получить потомство от тритонов, выращенных на темном грунте, и поместить их на светлый, то все они будут иметь светлую окраску, а не темную, как их родители. То есть данный вид изменчивости не затрагивает генотип и поэтому не передается потомкам.
Изучение влияния новых условий на человека показало, что такие признаки, как тип обмена веществ, предрасположенность к некоторым заболеваниям, группа крови, узоры кожи на пальцах, и другие определяются генотипом, и их выражение мало зависит от факторов окружающей среды. Другие признаки, такие как уровень интеллекта, вес, рост и т.п., обладают широким диапазоном изменений, и их проявление в значительной степени определяется окружающей средой. Те внешние различия, которые обусловлены средой, получили название модификаций. Модификации не связаны с изменением генетических структур особи, а являются лишь частной реакцией генотипа на конкретные изменения окружающей среды (температуры, содержания кислорода во вдыхаемом воздухе, характера питания, воспитания, обучения и т.д.).
Модификационная изменчивость носит групповой характер (групповая), то есть все особи одного вида, помещенные в одинаковые условия, приобретают сходные признаки. Например, если сосуд с эвгленами зелеными поместить в темноту, то все они утратят зеленую окраску, если же вновь выставить на свет – все опять станут зелеными.
Модификационная изменчивость является определенной, то есть всегда соответствует факторам, которые ее вызывают. Так, усиленные физические нагрузки влияют на степень развития мышц. Под действием ультрафиолетовых лучей у людей (если они не альбиносы) возникает загар в результате накопления в коже меланина, причем у разных людей интенсивность окраски кожи различна.
В оптимальных условиях выявляется максимум возможностей, присущих данному генотипу. Так, продуктивность беспородных животных в условиях улучшенного содержания и ухода повышается (надои молока, нагул мяса). В этом случае все особи с одинаковым генотипом отвечают на внешние условия одинаково, поэтому Ч. Дарвин этот тип изменчивости назвал определенной или групповой изменчивостью. Однако другой
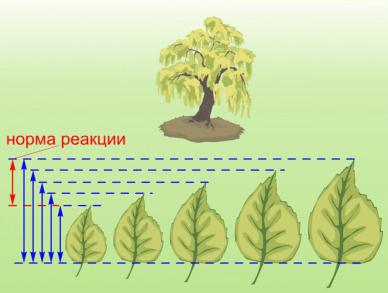
признак – жирность молока – слабо подвержен изменениям условий среды, а масть животного – еще более устойчивый признак. Интенсивность модификационной изменчивости можно регулировать. Модификационная изменчивость направлена.
Рис. 4. Норма реакции.
Модификационная изменчивость признака может быть очень велика, но она всегда контролируется генотипом организма и обычно колеблется в определенных пределах. Конкретные изменения не наследуются, они формируются в процессе жизнедеятельности особи. Наследуется генотип с его специфической нормой реакции на изменение среды. Внешние воздействия могут вызывать у особи или группы особей изменения, которые бывают для них вредными, безразличными или полезными, т.е. приспособительными.
Границы фенотипической изменчивости, контролируемые генотипом организма, называют нормой реакции (рис. 4).
Широкая норма реакции приводит к повышению выживаемости. Широкая норма реакции свойственна количественным признакам, таким как удои молока, размеры листьев, окраска у некоторых бабочек; узкая норма реакции для качественных признаков – жирности молока, яйценоскости у кур, интенсивности окраски венчиков у цветков и др.
Одним из первых исследователей, изучавших модификационную изменчивость, был К. Нэгели (1865), который сообщил, что если альпийские формы растений, например ястребинки, перенести на богатую почву Мюнхенского ботанического сада, то у них
обнаруживаются увеличение мощности, обильное цветение, а некоторые растения изменяются до неузнаваемости. Если же формы вновь перенести на бедные каменистые почвы, то они возвращаются к исходной форме. Несмотря на полученные результаты, К. Нэгели оставался сторонником наследования приобретенных свойств.
Впервые строгий количественный подход к исследованию модификационной изменчивости с позиций генетики применил В. Иогансен. Он изучал наследование массы и размера семян фасоли – признаков, в значительной степени меняющихся под влиянием как генетических факторов, так и условий выращивания растений.
Убежденным противником наследования свойств, приобретенных в онтогенезе, был А. Вейсман (1833-1914). Последовательно отстаивая дарвиновский принцип естественного отбора как движущую силу эволюции, он предложил разделить понятия соматогенных и бластогенных изменений, т.е. изменения свойств соматических клеток и органов, с одной стороны, и изменения свойств генеративных клеток – с другой. А. Вейсман указал на невозможность существования механизма, который передавал бы изменения соматических клеток половым таким образом, чтобы в следующем поколении организмы изменялись адекватно тем модификациям, которые претерпели родители во время своего онтогенеза.
Иллюстрируя это положение, А. Вейсман поставил следующий эксперимент, доказывавший ненаследование приобретенных признаков. На протяжении 22 поколений он отрубал хвосты белым мышам и скрещивал их между собой. В общей сложности он обследовал 1592 особи и ни разу не обнаружил укорочения хвоста у новорожденных мышат.
Типы модификационной изменчивости
Различают следующие типы фенотипической изменчивости: модификации, морфозы и фенокопии.
Модификации – это ненаследственные изменения, которые возникают под действием фактора среды, носят адаптивный характер и чаще всего обратимы (например, увеличение эритроцитов в крови при недостатке кислорода).
Свойства модификаций:
1)ненаследуемость;
2)групповой характер изменений;
3)соотнесение изменений действию определенного фактора среды;
4)обусловленность пределов изменчивости генотипом.

При интенсивном действии многих агентов наблюдаются ненаследуемые изменения, случайные (по своему проявлению) по отношению к воздействию. Такие изменения называют морфозами. Очень часто они напоминают фенотипическое проявление известных мутаций. Тогда их называют фенокопиями этих мутаций.
В конце 30-х – начале 40-х годов И.А. Рапопорт исследовал действия на дрозофилу многих химических соединений, показав, что, например, соединения сурьмы – brown (коричневые глаза); мышьяковистая кислота и некоторые другие соединения – изменения крыльев, пигментации тела; соединения бора – eyeless (безглазие), aristopedia (превращение арист в ноги), соединения серебра – yellow (желтое тело) и т.д. При этом некоторые морфозы при воздействии на определенную стадию развития индуцировались с высокой частотой (до 100%).
Морфозы – это ненаследственные изменения, вызванные экстремальными или необычными факторами среды (рентгеноморфозы, хемоморфозы), которые изменяют соматические клетки и необратимы (например, ожоги, шрамы). Морфозы рассматривают как «уродства», которые не наследуются и не носят адаптивного характера. Например, при облучении личинок дрозофилы получают имаго с вырезками в различных частях крыла, которые являются следствием гибели части клеток имагинальных дисков крыла вследствие облучения.
Фенокопии – это ненаследственные изменения, сходные с известными мутациями (вариант эмбриопатии). Фенокопии являются результатом действия физических и химических агентов на генетически нормальный организм. Например, при использовании талидомида часто рождались дети с фокомелией (рис. 5) – укороченными ластовидными руками, которую могут вызвать и мутантные аллели.
Рис.5. Фокомелия (греч. phoke – тюлень, melos – часть тела, конечность).
Фенотип формируется в результате взаимодействий генотипа и факторов среды. Фенотипические признаки не передаются от родителей потомкам, наследуется лишь норма реакции, т. е. характер реагирования на изменение окружающих условий. У гетерозиготных организмов при изменении условий среды можно вызвать различные проявления данного признака.
Для оценки степени выраженности изучаемого признака используют понятие: экспрессивность – степень фенотипического проявления гена. Этот показатель зависит от взаимодействия гена с другими генами или от воздействия внешних условий. Наличие данного гена не всегда означает, что он проявится в фенотипе. Для оценки количества особей, у которых этот признак фенотипически проявился, используют термин «пенетрантность».
Пенетрантность – это частота фенотипического проявления признака у особей с одинаковым генотипом по этому гену. Например, пенетрантность врожденного вывиха бедра составляет 20%, сахарного диабета – 65%.
Виды модификаций
Различают возрастные, сезонные и экологические модификации. Они сводятся к изменению лишь степени выраженности признака; нарушения структуры генотипа при них не происходит. Следует отметить, что четкую границу между возрастными, сезонными и экологическими модификациями провести невозможно.
Возрастные, или онтогенетические, модификации выражаются в виде постоянной смены признаков в процессе развития особи. Это наглядно демонстрируется на примере онтогенеза земноводных (головастики, сеголетки, взрослые особи), насекомых (личинка, куколка, имаго) и других животных, а также растений. У человека в процессе развития наблюдаются модификации морфофизиологических и психических признаков. Например, ребенок не сможет правильно развиваться и физически, и интеллектуально, если в раннем детстве на него не будут оказывать влияние нормальные внешние, в том числе социальные, факторы. Например, долгое пребывание ребенка в социально неблагополучной среде может вызвать необратимый дефект его интеллекта.
Онтогенетическая изменчивость, как и сам онтогенез, детерминируется генотипом, где закодирована программа развития особи. Однако особенности формирования фенотипа в онтогенезе обусловлены

взаимодействием генотипа и среды. Под влиянием необычных внешних факторов могут происходить отклонения в формировании нормального фенотипа.
Таким образом, совокупность признаков особи (ее фенотип) является результатом реализации генетической информации в конкретных условиях окружающей среды. Формируется фенотип в процессе индивидуального развития, начиная с момента оплодотворения. Физическое, психическое и умственное здоровье человека – это результат взаимодействия унаследованных человеком особенностей с факторами окружающей среды, воздействующими на него на протяжении всей жизни. Ни наследственность, ни окружающая человека среда не являются неизменными. Этот важный принцип лежит в основе современного понимания процессов изменчивости и наследственности. В мире нельзя найти двух людей, за исключением однояйцовых близнецов (развившихся из одной оплодотворенной яйцеклетки), обладающих одинаковым набором генов. Нельзя также найти двух людей, проживших жизнь в одинаковых условиях. Наследственность и среда не противопоставляются друг другу: они едины и немыслимы одна без другой.
Рис. 6. Заяц-беляк летом и зимой.
Сезонные модификации особей или целых популяций проявляются в виде генетически детерминированной смены признаков (например, изменение окраски шерсти (рис. 6), появление подпушка у животных), происходящей в результате сезонных изменений климатических условий.
Ярким примером такой изменчивости является опыт с горностаевым кроликом. У горностаевого кролика на спине
выбривают наголо определенный участок (спина горностаевого кролика покрыта белой шерстью), и затем кролика
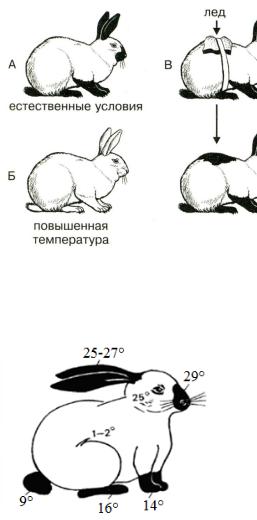
помещают на холод. Оказывается, что в таком случае на оголенном месте, подвергшемся влиянию низкой температуры, появляется темнопигментированный волос и в результате на спине – темное пятно (рис. 7).
Рис. 7. Опыт с горностаевым кроликом
.
Очевидно, что развитие того или иного признака кролика – его фенотип, в данном случае – горностаевая окраска, зависит не только от его генотипа, но и от всей совокупности условий, в которых происходит это развитие.
Советский биолог Ильин показал, что температура окружающей среды имеет большое значение в развитии пигмента у горностаевого кролика, причем для каждой области тела есть свой порог температуры, выше
Экологические модификации
которого вырастает белая шерсть, а ниже – черная (рис. 8).
Рис 8. Карта температурных порогов пигментации шерсти у горностаевого кролика (из Ильина по С.М. Гершензону)
представляют собой адаптивные
изменения фенотипа в ответ на изменение условий среды. Сезонные модификации можно отнести к группе экологических модификаций. Экологические модификации фенотипически проявляются в изменении степени выраженности признака. Они могут возникать на ранних стадиях развития и сохраняться в течение всей жизни. Классическим примером экологических модификаций могут служить различные формы листовой

пластины у стрелолиста, обусловленные влиянием среды: погруженные в воду листья имеют лентовидную форму, листья, плавающие на поверхности воды, – округлую, а находящиеся в воздушной среде, – стреловидные. Если же все растение оказывается полностью погруженным в воду, его листья только лентовидные (рис. 9).
листья в воздушной |
|
плавающие листья – |
|
погруженные в |
|
|
воду листья – |
||
среде – стреловидные |
|
округлые |
|
|
|
|
лентовидные |
||
|
|
|
|
|
|
|
|
|
|
Рис. 9. Растение стрелолиста, образующее три типа листьев: надводные, плавающие и подводные.
Экологические модификации затрагивают количественные (количество лепестков в цветке, потомства у животных, масса животных, высота растений, размер листа и т.д.) и качественные (окраска цветков у медуницы, чины лесной, примулы; цвет кожи у человека под влиянием ультрафиолетовых лучей и др.) признаки.
Рис.10. Листья водяного лютика.
Так, например, Леваковский при выращивании в воде ветки ежевики вплоть до ее распускания обнаружил существенные изменения в

анатомическом строении ее ткани. В аналогичном эксперименте Константен выявил фенотипические различия в строении надводной и
подводной частей листа у лютика (рис.10). |
|
|||||
В 1895 г. французский ботаник Г. |
|
|||||
Боннье провел опыт, ставший |
|
|||||
классическим |
|
примером |
|
|||
экологической модификации. Он |
|
|||||
разделил |
одно |
|
растение |
|
||
одуванчика на две части и |
|
|||||
выращивал |
их |
в |
разных |
|
||
условиях: на равнине и высоко в |
|
|||||
горах. Первое растение достигло |
|
|||||
нормальной высоты, а второе |
|
|||||
оказалось карликовым (рис.11). |
|
|||||
Такого рода изменения бывают и |
|
|||||
у животных. Например, Р. |
|
|||||
Вольтерк в 1909 г. наблюдал |
|
|||||
изменения |
высоты |
|
шлема |
у |
Рис. 11. Опыт Г. Боннье. |
|
дафний в |
зависимости |
от |
||||
|
||||||
условий питания.
Экологические модификации, как правило, обратимы и со сменой поколений при условии изменения внешней среды могут проявиться. Например, потомство низкорослых растений на хорошо удобренных почвах будет нормальной высоты; определенное количество лепестков в цветке какого-либо растения в потомстве может не повториться; у человека с кривыми ногами вследствие рахита бывает вполне нормальное потомство. Если же в ряду поколений условия не меняются, степень выраженности признака в потомстве сохраняются, ее нередко принимают за стойкий наследственный признак (длительные модификации).
Статистические закономерности модификационной изменчивости
К статистическим закономерностям модификационной изменчивости относятся варианты, вариационный ряд изменчивости признака и вариационная кривая.
Модификационная изменчивость многих признаков растений, животных и человека подчиняется общим закономерностям. Эти закономерности выявляются на основании анализа проявления признака у группы особей (n). Степень выраженности изучаемого признака у членов выборочной
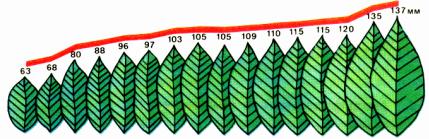
совокупности различна. Каждое конкретное значение изучаемого признака называют вариантой и обозначают буквой ν. При изучении изменчивости признака в выборочной совокупности составляется вариационный ряд, в котором особи располагаются по возрастанию показателя изучаемого признака.
Вариационный ряд представляет ряд вариант, расположенных в порядке убывания или возрастания (например, если собрать листья с одного и того же дерева и расположить их по мере увеличения длины листовой пластинки, то получается вариационный ряд (рис. 12) изменчивости данного признака).
Рис. 12. Вариационный ряд листьев лавровишни (цифрами показана длина листьев).
На основании вариационного ряда строится вариационная кривая – это графическое изображение зависимости между размахом изменчивости признака и частотой встречаемости отдельных вариант данного признака (рис. 13). Частота встречаемости отдельных вариант обозначается буквой p. Например, если взять 100 колосьев пшеницы (n) и подсчитать число колосков в колосе, то это количество будет от 14 до 20 – это численное значение вариант (ν). Вариационный ряд: ν = 14-15-16-17-18-19-20. Частота встречаемости каждой варианты p = 2-7-22-32-24-8-5. Среднее значение признака встречается чаще, а вариации, значительно отличающиеся от него, – намного реже. Это называется нормальным распределением. Кривая на графике бывает, как правило, симметричной. Вариации, как большие, чем средние, так и меньшие, встречаются одинаково часто.
Наиболее типичный показатель признака – это его средняя величина, то есть среднее арифметическое вариационного ряда.
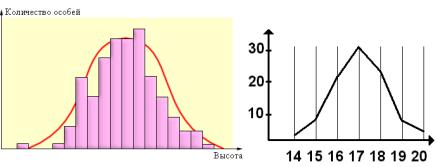
Рис. 13. Вариационная кривая.
Среднее значение данного признака легко рассчитать. Для этого используют формулу: М = Ʃ (ν х p) ∕ n, где М – средняя величина признака, в числителе сумма произведений вариант на их частоту встречаемости, в знаменателе – количество вариант (n). Для данного признака среднее значение равно 17,1.
Знание закономерностей модификационной изменчивости имеет большое практическое значение, поскольку позволяет предвидеть и заранее планировать степень выраженности многих признаков организмов в зависимости от условий внешней среды.
Характеристики модификационной изменчивости
1.Адаптивные изменения (например, стрелолист).
2.Приспособительный характер. Это означает, что в ответ на изменившиеся условия среды у особи проявляются такие фенотипические изменения, которые способствуют их выживанию. Примером служит изменение содержания влаги в листьях растений в засушливых и влажных районах, окраски у хамелеона, формы листа у стрелолиста в зависимости от условий среды.
3.Обратимость в пределах одного поколения, т.е. со сменой внешних условий у взрослых особей меняется степень выраженности тех или иных признаков. Например, у крупного рогатого скота в зависимости от условий содержания может колебаться удой и жирность молока, у кур – яйценоскость.
4.Модификации адекватны, т.е. степень выраженности признака находится в прямой зависимости от вида и продолжительности действия того или иного фактора. Так, улучшение содержания скота способствует увеличению живой массы животных, плодовитости, удоя и жирности молока; на удобренных почвах при оптимальных климатических условиях повышается урожайность зерновых культур и т.д.
5.Массовый характер. Массовость обусловливается тем, что один и тот же фактор вызывает примерно одинаковое изменение у особей, сходных генотипически.
6.Длительные модификации. Впервые были описаны в 1913 г. нашим соотечественником В. Иоллосом. Путем раздражения инфузорий туфелек он вызвал у них появление ряда морфологических особенностей, которые сохранялись в течение большого числа поколений, до тех пор, пока размножение было бесполым. При изменении условий развития длительные модификации не наследуются. Поэтому ошибочно мнение, что воспитанием и внешним воздействием можно закрепить в потомстве новый признак. Например, предполагалось, что от хорошо дрессированных животных потомство получается с лучшими «актерскими» данными, чем от недрессированных. Потомство дрессированных животных действительно легче поддается воспитанию, но объясняется это тем, что оно наследует не приобретенные родительскими особями навыки, а способность к дрессировке, обусловленную наследуемым типом нервной деятельности.
7.Норма реакций (предел модификации). Именно норма реакции, а не сами модификации, наследуются, т.е. наследуется способность к развитию того или иного признака, а форма его проявления зависит от условий внешней среды. Норма реакции – это конкретная количественная и качественная характеристика генотипа, т.е. определенное сочетание генов в генотипе и характер их взаимодействия.
Примеры модификационной изменчивости
У человека:
увеличение уровня эритроцитов при подъеме в горы; увеличение пигментации кожи при интенсивном воздействии ультрафиолетовых лучей;
развитие костно-мышечной системы в результате тренировок; шрамы (пример морфоза).
У насекомых и других животных:
изменение окраски у колорадского жука вследствие длительного влияния на их куколки высоких или низких температур;
смена окраски шерсти у некоторых млекопитающих при изменении погодных условий (например, у зайца);
различная окраска бабочек-нимфалид (например, Araschnia levana), развивавшихся при разной температуре.
У растений:
различное строение подводных и надводных листьев у водяного лютика, стрелолиста и др.; развитие низкорослых форм из семян равнинных растений, выращенных в горах.
У бактерий:
работа генов лактозного оперона кишечной палочки (при отсутствии глюкозы и в присутствии лактозы они синтезируют ферменты для переработки этого углевода).
42. Определение понятия "ген". Классификация генов. Современное состояние теории гена.
Ген - это информационная структура, состоящая из нуклеотидов ДНК (а у вирусов и РНК), неделимая в функциональном отношении, способная к неограниченной репликации и направляющая развитие и функционирование организма, обеспечивая в том числе и транскрипцию других генов.
Обширная многолетняя (1990-2000 гг.) программа "Геном человека", заключающаяся в последовательном секвенировании участков генома человека, установила, что генотип человека - это всего лишь 30000 генов. Сотни генов получены человеком, вероятно, в результате горизонтальной передачи, начиная от бактерий, – у человека и бактерий одинаковы более 600 генов; геном мышей и человека совпадает на 90%; шимпанзе и человека - различается на 1% нуклеотидных последовательностей.
Вгеноме каждый ген может быть представлен несколькими формами – аллелями: в гаплоидном геноме - одним аллелем (любым), в
диплоидном – двумя (доминантным и рецессивным), в генофонде популяции – несколькими (более, чем двумя). Если у гетерозигот оба аллеля одинаково активны и каждый участвует в синтезе 50% продукта, считается, что оба имеют равные дозы. Однако дозы гена могут быть и разные. В крови у гетерозигот по гену серповидноклеточной анемии содержится 65% нормального гемоглобина и 35% аномального (НЬS).Молекулярный механизм разных доз аллельных генов до сих пор неясен.
Взависимости от выполняемой функции различают гены: структурные –
транскрибируются во все виды РНК; регуляторные – регулируют процесс транскрипции; модуляторы – изменяют активность структурных генов.
Согласно хромосомной теории, каждый ген занимает в хромосоме свое определенное место - локус. Однако существуют прыгающие или блуждающие гены, которые называются транспозоны (мобильные элементы геномов). У прокариот плазмиды способны переносить генетическую информацию между бактериями, частично или целиком встраиваясь в геном клетки-хозяина. К таким плазмидам относятся факторы F (от английского fertility-плодовитость) и гены устойчивости к лекарственным препаратам в плазмидах R.
Но если интеграция плазмиды F в геном клетки-хозяина происходит в определенных местах генома, то гены устойчивости могут встраиваться во многие места хромосомы - такие гены и называются транспозонами. Впервые явление транспозиции было описано у эукариот много лет назад Барбарой Мак-Клинток, проводившей генетические исследования на кукурузе. Сейчас известно несколько систем блуждающих контролирующих элементов эукариот. В частности, контролирующие системы у кукурузы, которые отличаются по своему воздействию на структурные гены, влияя на экспрессию генов, контролирующих окраску семян кукурузы (транспозиция регуляторных элементов осуществляется автономно); транспозоны Drosophila melanogaster, встраивающиеся в Х- хромосому и вызывающие делеции в соседних генах, что приводит к изменению цвета глаз у плодовой мушки; описаны транспозирующие элементы дрожжей.
Транспозоны про - и эукариот, по-видимому, не могут существовать вне генома. Неизвестно, существует ли взаимосвязь между транспозонами разных видов и сохраняются ли функции транспозиции при перемещении мобильных элементов из одного вида в другие.
Присутствие транспозонов в геноме может индуцировать перестройки,
обусловливает высокую частоту обмена генетической информацией. Эти элементы способствуют быстрому распространению генов в популяции, управляют процессами дифференцировки, ускоряют эволюцию про - и эукариот. Перемещение определенных последовательностей из одного специфического сайта в другой имеет регуляторный эффект. В случае иммуноглобулиновых генов в результате рекомбинации меняется содержание генома, создаются активные гены в соответствующих соматических клетках.
В нуклеотидной последовательности структурных генов, транскрибируемых в и-РНК, должно быть достаточно кодирующих единиц, чтобы зашифровать 20 аминокислот. Генетический код триплетен, поэтому кодирующих единиц более, чем достаточно - их 64. Последовательность из трех нуклеотидов (триплет), соответствующая
одной аминокислоте, называется кодоном, их 61. Три триплета из 64 (УАГ, УАА, УГА) - это кодоны-терминаторы, которые располагаются в окончании структурного гена и и-РНК, они транскрибируются, но никогда не транслируются - на них синтез белка останавливается.
Так как кодирующих единиц больше, чем кодируемых, то почти каждой аминокислоте соответствует несколько кодонов-синонимов (кроме триптофана и метионина). Эта особенность или свойство триплетного кода называется избыточностью или вырожденностью кода.
При этом генетический код универсален, т.е. любая аминокислота в любой форме жизни зашифрована одинаково; генетический код специфичен, т.е. каждый триплет кодирует только свою аминокислоту. В
пределах одной рамки считывания, что означает транскрипцию информации об одном полипептиде, генетический код неперекрываем - каждый нуклеотид входит в состав только своего триплета.
Ген как единица функции (с этой позиции он называется цистрон) неделим. Однако в нем выделяют разные составляющие: структурная составляющая - нуклеотид; информационная - триплет (кодон); кроме того, в гене различают единицы мутации (мутон) и единицы рекомбинации (рекон), соответствующие любой паре комплементарных нуклеотидов.
В 1977 г. было обнаружено, что гены эукариот состоят из последовательностей ДНК двух типов: экзонов - информативных участков и интронов – неинформативных. Транскрибируется весь ген, но транслируется только его экзонная часть. У эукариот в процессе биосинтеза белка между транскрипцией и трансляцией происходит ряд событий, которые объединяются термином процессинг. Во время процессинга происходит преобразование проинформационной РНК (первичного транскрипта) в информационную: специфические ферменты (рестриктазы) вырезают интронные участки, оставшиеся экзоны сшиваются другими ферментами (лигазами) – это событие называется сплайсингом; в передней части (5'-конец) про-и-РНК формируется группа нуклеотидов, получивших название "колпачок" и предназначенных для узнавания рибосомой и-РНК; в хвостовой части (З'-конец) про-и-РНК формируется полиадениновый "хвост" (100-200 адениновых нуклеотидов), назначение которого пока что неизвестно.
После всех этих событий формируется и-РНК, готовая к трансляции. Благодаря экзонно-интронной структуре гена у эукариот в пределах одной полинуклеотидной последовательности структурного гена может быть закодирована информация не об одной молекуле белка, а более. Извлекается эта разная информация из одного гена в результате
альтернативного сплайсинга, когда некоторые участки про-и-РНК в одном случае ведут себя как экзоны, а при другом типе сплайсинга оказываются интронами.
Ген, являясь единицей функции, сам входит в состав единицы транскрипции, которой после открытия Жакоба и Моно (1961) является оперон. Точнее, единицей транскрипции является последовательность нуклеотидов в опероне от промотора до терминатора включительно. Осуществление транскрипции называется экспрессией гена или его генетической активностью.
Таким образом, современное состояние теории гена включает следую-
щие положения:
Ген - материальная единица хранения и передачи наследственной информации, является частью молекулы ДНК или у вирусов РНК.
Ген в хромосоме занимает определенный локус. Существуют гены с непостоянной локализацией - транспозоны.
Гены делятся на структурные, регуляторные, гены модуляторы.
Структурно-функциональной единицей гена является триплет.
Триплеты в гене расположены колинеарно аминокислотам в
белке.
Гены эукариот имеют интронно-экзонную природу.
Единицей транскрипции в геноме является оперон.
Гены способны к рекомбинации (как межгенной, так и внутригенной) и мутации.
Не всякое повреждение гена ведет к мутации, т.к. гены способны к репарации.
Дискретные единицы - гены формируют целостную систему взаимодействующих генов – генотип.
43.Регуляция генной активности (экспрессия генов) у про- и эукариот.
Организмы, обладающие способностью регулировать свою генетическую активность, хорошо адаптируются к изменяющимся условиям окружающей среды.
Наличие таких регуляторных систем характерно для всех эукариотических и прокариотических клеток. Специфические механизмы регуляции активности (экспрессии) генов были установлены французскими исследователями Франсуа Жакобом и Жаком Моно в 1961 году. Они предложили гипотезу "оперона", которая впоследствии была названа классической, и ее авторы были удостоены Нобелевской премии. На базе этого исследования была впервые разработана модель структурно-
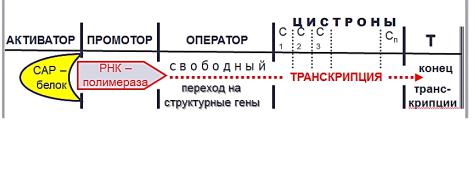
функциональной организации оперона. В настоящее время оперонная теория получила экспериментальное подтверждение.
Строение оперона Оперон (рис. 13) - это блок генов, участвующих в обеспечении
транскрипции генов, ответственных за синтез определенного генопродукта.
Рисунок 13. Схема строения оперона
Схема оперона: Регуляторная часть оперона:
А - активатор, часть промотора, к которому присоединяется белокактиватор (САР - белок или catabolite activator protein), что активирует присоединение РНК - полимеразы к промотору; это "положительно" контролирующий элемент, который есть не в каждом опероне.
П - ген-промотор - это участок ДНК, который распознается ферментом РНК - полимеразой и указывает место, где должна начинаться транскрипция.
О - ген-оператор, управляющий работой структурных генов; "негативно" контролирующий элемент - присутствие на нем белка-репрессора прекращает транскрипцию.
Т - ген-терминатор - это участок, после которого прекращается транскрипция и перед которым прекращается трансляция. В состав этого участка входит один из трех кодонов терминаторов (стоп-кодонов). В некоторых оперонах между оператором и структурными генами расположен участок(16 пар оснований), частью которого является аттенуатор, служащий барьером для транскрипции. Подобная структура есть в триптофановом опероне кишечной палочки (Escherichia coli).
Цистронная часть оперона: С1, С2, С3 – структурные гены, кодирующие соответствующие белки; структурные гены одного оперона включаются и выключаются одновременно.
Транскрипция группы структурных генов (цистронов) контролируется
геном-регулятором и геном-оператором. Оператор состоит приблизительно из 30 нуклеотидов. Генетические дефекты в операторе
приводят к непрерывному синтезу ферментов, т.е. регуляция синтеза генопродукта нарушается. Ген-регулятор контролирует синтез белка репрессора, не входит в состав оперона и может находиться на разном расстоянии от оперона.
Регуляторный белок репрессор определяет активность оперона. Он имеет два функциональных центра: 1) место связывания с опероном; 2) место связывания с индуктором или корепрессором. Большее сродство белок репрессор имеет ко второй группе веществ, которые высокоспецифичны.
Оперон активен, если оператор свободен от репрессора. Этот белок с оператора снимается, если к его второму активному центру присоединяется вещество, называемое индуктором (по химической природе оно может быть различным). Следовательно, регуляторные белки либо запускают, либо блокируют транскрипцию цистронной части оперона.
Таким образом, анализируя механизмы регуляции экспрессии генов у прокариот, можно выделить три типа регуляторных элементов.
1.Регуляторные белки – белки, влияющие на активность РНКполимеразы, т.к. или позволяют ей связываться с промотором или нет; или открывают ей доступ к следующим после промотора нуклеотидам ДНК, или закрывают, соединяясь с оператором. Активность регуляторных белков изменяется с помощью специфического связывания с низкомолекулярными эффекторами (индукторами, корепрессорами).
2.Эффекторы - небольшие небелковые молекулы, концентрация которых в клетке отражает её состояние. В качестве эффектора могут выступать циклический аденозинмонофосфат, триптофан, лактоза и др.
3.Регуляторные нуклеотидные последовательности оперона
(промоторы, операторы, терминаторы, аттенуаторы), действуя на которые, регуляторные белки, влияют на уровень синтеза соответствующих и-РНК.
Регуляция активности генов у прокариот
В процессе синтеза катаболических ферментов (расщепляющих субстраты) у прокариот происходит индуцируемый синтез ферментов. Это дает клетке возможность приспосабливаться к условиям окружающей среды и экономить энергию, прекращая синтез соответствующего фермента, если потребность в нем исчезает.
Для индукции синтеза катаболических ферментов обязательны следующие условия:
1. Фермент синтезируется только тогда, когда расщепление соответствующего субстрата необходимо для клетки.

2. Концентрация субстрата в среде должна превысить определенный уровень, прежде чем соответствующий фермент сможет образоваться. Наиболее хорошо изучен механизм регуляции экспрессии генов у кишечной палочки на примере lac-оперона, контролирующего синтез трех катаболических ферментов, расщепляющих лактозу (рис.14) . Если в клетке много глюкозы и мало лактозы, промотор остается неактивным, а на операторе находится белок репрессор - блокируется транскрипция lacоперона.
Когда количество глюкозы в среде, а следовательно и в клетке, уменьшается, а лактозы увеличивается, происходят следующие события: количество циклического аденозинмонофосфата увеличивается, он связывается с САР -белком - этот комплекс активирует промотор, с которым соединяется РНК-полимераза ; в это же время избыток лактозы соединяется с белком-репрессором и освобождает от него оператор - путь для РНК-полимеразы открыт, начинается транскрипция структурных генов lac -оперона. Лактоза выступает в качестве индуктора синтеза тех ферментов, которые её расщепляют.
Лактозный оперон будет находиться в состоянии экспрессии до тех пор, пока в клетке уровень индуктора - лактозы не будет доведен до определенного уровня, характерного для данной клетки (принцип обратной связи). Тогда белок-репрессор освободится от лактозы, займет свое место на операторе, и транскрипция оперона прекратится.
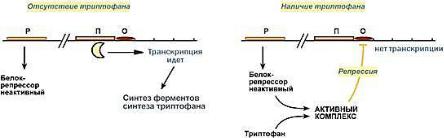
Рисунок 14. Механизм работы лактозного оперона.
Такая регуляция синтеза катаболических ферментов получила название негативной индукции, т.к. сам белок-репрессор осуществляет негативный контроль работы оперона (его присутствие на операторе выключает транскрипцию), а снимается блок транскрипции благодаря индуктору, который инактивирует белок-репрессор.
В настоящее время изучена работа многих оперонов, в том числе и оперонов анаболического ряда. Примером такого оперона у кишечной палочки может быть триптофановый оперон, контролирующий синтез пяти ферментов, необходимых для образования аминокислоты триптофана (рис.15).
Для триптофанового оперона синтезируется неактивный репрессор, который активируется лишь под действием корепрессора (триптофана). Здесь наблюдается особая форма ингибирования конечным продуктом: оперон становится активным в случае недостатка триптофана в среде, а высокое содержание в среде данной аминокислоты подавляет выработку фермента, необходимого для синтеза триптофана, т.к. избыток
триптофана активирует белок-репрессор, который соединяется с оператором, и транскрипция прекращаетсяоперон репрессируется. Такая система регуляции называется негативной репрессией. Она позволяет не синтезировать вещество в избытке.
Особенности регуляции экспрессии генов у эукариот
Регуляция экспрессии генов у эукариот протекает намного сложнее. Различные типы клеток многоклеточного эукариотического организма синтезируют ряд одинаковых белков, и в то же время они отличаются друг от друга набором белков, специфичных для клеток данного типа. Уровень продукции зависит от типа клеток, а также от стадии развития
организма. Регуляция экспрессии генов осуществляется на уровне клетки и на уровне организма.
Гены эукариотических клеток делятся на два основных вида: первый определяет универсальность клеточных функций, второй – детерминирует (определяет) специализированные клеточные функции.
Функции генов первой группы проявляются во всех клетках. Для осуществления дифференцированных функций специализированные клетки должны экспрессировать определенный набор генов.
Хромосомы, гены и опероны эукариотических клеток имеют ряд структурно-функциональных особенностей, что объясняет сложность экспрессии генов.
1.Опероны эукариотических клеток имеют несколько генов - регуляторов, которые могут располагаться в разных хромосомах.
2.Структурные гены, контролирующие синтез ферментов одного биохимического процесса, могут быть сосредоточены в нескольких оперонах, расположенных не только в одной молекуле ДНК, но и в нескольких.
3.Сложная последовательность молекулы ДНК. Имеются информативные и неинформативные участки, уникальные и многократно повторяющиеся информативные последовательности нуклеотидов.
4.Эукариотические гены состоят из экзонов и интронов, причем созревание и-РНК сопровождается вырезанием интронов из соответствующих первичных РНК-транскриптов (про-и-РНК), т.е. сплайсингом.
5. Процесс транскрипции генов зависит от состояния хроматина. Локальная компактизация ДНК полностью блокирует синтез РНК.
6.Транскрипция в эукариотических клетках не всегда сопряжена с трансляцией. Синтезированная и-РНК может длительное время сохраняться в виде информосом. Транскрипция и трансляция происходят
в разных компартментах.
7.Некоторые гены эукариот имеют непостоянную локализацию
(лабильные гены или транспозоны).
8.Методы молекулярной биологии выявили тормозящее действие белков-гистонов на синтез и-РНК.
9.В процессе развития и дифференцировки органов активность генов зависит от гормонов, циркулирующих в организме и вызывающих специфические реакции в определенных клетках. У млекопитающих важное значение имеет действие половых гормонов.
10.У эукариот на каждом этапе онтогенеза экспрессировано 5-10% генов, остальные должны быть заблокированы
44.Репарация генетического материала - виды репараций. Цитоплазматическая наследственность.
Важное значение для ограничения неблагоприятных последствий генных мутаций имеют естественные антимутационные барьеры. Одним из них является парность хромосом в диплоидных наборах хромосом эука-риот, которая препятствует проявлению рецессивных мутаций у гетерозиготных особей. Главным антимутационным барьером рассматривается выработавшая в процессе эволюции способность к репарации наследственного материала. Её сущность - в устранении из наследственного материала клетки изменённого участка. , Различают 3 системы репарации генетического материала: эксци-зионная репарация (репарация путём «вырезания»), фоторепарация и пострепликативная репарация.
Механизм эксцизионной репарации заключается в ферментативном разрушении изменённого участка молекулы ДНК с последующим восстановлением на этом отрезке нормальной последовательности нуклеотидов. Такой механизм включает следующие этапы (рис. 81): а) разрыв спирали ДНК у места повреждения при участии эндонуклеаз; б) удаление поврежденного участка с запасом в обе стороны с помощью эндонуклеаз; в) синтез при участии ДНК-полимеразы на месте дефекта нормального участка ДНК; г) «сшивание» последнего с образовававшимися концами спирали ДНК при помощи фермента ДНК-лигазы (восстановление непрерывности ДНК).
Например, под действием УФ-лучей у человека нарушается комплементар-ность пар нуклеотидов в двойной спирали ДНК (появляются пары Т-Т, Ц-Ц и т.п.). Они устраняются вышеописанным способом. Однако у различных индивидуумов наблюдаются генетические различия в активности репаративньгх ферментов и надёжности функционирования механизма ферментативного разрушения изменённого участка молекулы ДНК в целом. У ряда людей наблюдается изменение ДНК и, как следствие, возникновение заболевания «пигментная ксеродерма». В клетках эукариот обнаружены два вида репарации «путём вырезания»: 1) более продолжительная репарация (длительность процесса - от 1 до 24 часов), восстанавливающая большой фрагмент ДНК (около 100 нуклеотидов); 2) быстродействующая репарация (продолжается от 5 минут до 2 часов), восстанавливающая 3-4 нуклеотида.
Пострепликативная репарация «включается» тогда, когда эксци-зионная репарация «не справляется» с устранением всех повреждений, возникших в ДНК до её репликации. При репликации во второй спирали ДНК возникают бреши - однонитевые пробелы, соответствующие изменённым нуклеотидам первой спирали. Бреши заполняются участками цепи с нормальной последовательностью нуклеотидов уже в ходе пострешгикативной репарации при участии ДНК-полимеразы. Фоторепарация заключается в расщеплении ферментом (дезоксирибопиримидинфотолиазой), активируемым видимым светом, циклобутановых димеров, возникающих в ДНК под действием ультрафиолетового излучения.
Механизмам репарации свойственны нарушения и «сбои», которые приводят к повышению чистоты мутаций. Известны специфические мутации, блокирующие механизмы репарации и вызывающие наследственные заболевания (пигментная ксеродерма и др.). Биологическое значение репарации ДНК заключается в резком снижении частоты мутаций, большинство которых оказываются летальными и полулетальными или же снижающими жизнеспособность организмов, вызывающими аномалии и обусловливающими тератогенез. Благодаря репарации ДНК повышается устойчивость генотипа организма к повреждающим агентам (мутагенам).
Фотореактивация уменьшение повреждающего действия ультрафиолетового излучения на живые клетки при последующем воздействии на них ярким видимым светом. Ф. открыта в 1948 И. Ф. Ковалевым (СССР), А. Келнером и Р. Дульбекко (США) в результате опытов, проведённых на инфузориях парамециях, коловратках, конидиях грибов, бактериях и бактериофагах. В основе Ф. лежит ферментативное расщепление на мономеры пиримидиновых димеров, образующихся в ДНК под влиянием ультрафиолетового излучения. Ф. возникла в процессе эволюции как защитное приспособление от губительного действия УФкомпонента солнечного излучения и является одной из важнейших форм Репарации живых организмов от повреждений их генетического аппарата. Позднее была обнаружена и темновая репарация, т. е. свойство клеток ликвидировать повреждения ДНК без участия видимого света. Темновая репарация осуществляется комплексом из пяти ферментов:
узнающего химические изменения на участке цепи ДНК; осуществляющего вырезание поврежденного участка; удаляющего этот участок;
синтезирующего новый участок по принципу комплементарности взамен удаленного фрагмента;