

МОЛБИОЛ 2014-лекции / ООФ / Л05_РЕПАРАЦИЯ
.pdfЛекция 5
ДНК-лигаза репарирует дефект хромосомы
Репарация ДНК

Репарация,
или исправление повреждений в ДНК
Репарация генетических повреждений - способность живых организмов восстанавливать повреждения, возникшие в ДНК уже после ее синтеза
•От того, как клетки справляются с повреждениями ДНК, зависят такие кардинальные процессы, как появление наследственных болезней и раковых опухолей, а также старение.
•Ферменты репарации исправляют как ошибки, возникшие при репликации, так и не зависящие от репликации.
•Ферментативные системы репарации по сложности приближаются к репликационному ферментативному аппарату.
•Системы репарации и репликации имеют много общих компонентов.
•Принципы репарации ДНК у различных организмов сходны.

Дефекты ДНК после репликации
Этапы контроля последовательности дочерней цепи ДНК при репликации:
•Первичный отбор нуклеотидов идет по принципу комплементарности. Способностью к этому виду
отбора обладают все ДНК-полимеразы благодаря полимеризующей 5' 3' активности.
•Редактирующий отбор. Его проводят все ДНК-
полимеразы благодаря экзонуклеазной активности 3'
5'.
•Первичный и редактирующий отбор допускают примерно 1 ошибку на 109 включенных в ДНК нуклеотидов.
•Исправление ошибок в уже синтезированной ДНК. Этим занимаются ферменты репарации.

Повреждающие изменения в ДНК
Точечные мутации
Тип повреждения |
Последствия |
|||||
|
|
|
|
|
|
|
Нарушается правильное |
Влияние на будущие поколения |
|||||
спаривание оснований |
||||||
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
||||
|
Структурные нарушения |
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Тип повреждения |
|
Последствия |
|
|||
|
|
|
|
|
|
|
Введение одноцепочечных |
|
Нарушение матричных свойств |
|
|||
разрывов |
|
ДНК |
|
|||
|
|
|
|
|
|
|
Удаление основания или |
|
«---» |
|
|||
размыкание пуринового кольца |
|
|
|
|
||
|
|
|
|
|
|
|
Введение ковалентных связей |
|
Остановка полимеразы, брешь |
|
|||
между основаниями одной цепи |
|
|
|
|
||
|
|
|
|
|
|
|
Введение ковалентных связей |
|
Невозможность разделения цепей, |
|
|||
между основаниями на |
|
остановка полимеразы |
|
|||
антипараллельных цепях |
|
|
|
|
||
|
|
|
|
|
|
|

Как возникают повреждения ДНК, не зависящие от репликации?
1.Спонтанно.
2.При индукции мутагенами (химические вещества окружающей среды, многие углеводороды, реагенты, используемые в противоопухолевой химиотерапии) и
даже клеточными метаболитами (например, активные радикалы кислорода, образуемые во время нормального клеточного дыхания в различных биохимических путях).
3.Под действием ионизирующей радиации (гамма-лучи,
рентгеновские лучи).
4.Под действием ультрафиолетового излучения
(особенно ~260 нм, именно в этой области происходит максимальное поглощение ДНК).

1а. Спонтанные повреждения ДНК: апуринизация
•Происходит разрыв N-гликозидной связи между пуриновым основанием и дезоксирибозой.
•ДНК каждой клетки человека теряет за сутки 5 000-10 000 пуринов (!). В ДНК образуются апуриновые сайты (АР-сайты).
•Причины апуринизации: изменение рН, ионизирующее
излучение, повышение температуры и т.д.
•Пиримидины тоже могут отщепляться, но скорость этого процесса на два порядка ниже.
•Под AP-сайтом сейчас понимается апуриновый/апиримидиновый сайт.

6-оксипурин
2,6-диоксипурин
2,6- диоксипиримидин
1b. Спонтанные повреждения ДНК: дезаминирование
•Дезаминирование цитозина до урацила – наиболее частый случай дезаминирования (в ДНК каждой клетки человека происходит около 100 раз за сутки).
•Образование продуктов спонтанного дезаминирования может приводить к мутациям при последующей репликации ДНК, поскольку:
гипоксантин образует пару с C (А-Т -> HX-C),
ксантин – с Т (G-C -> X-T), урацил – с А (C-G -> U-A).
•Продукты дезаминирования не характерны для ДНК, их можно отличить от обычных оснований и удалить.
•Вероято, поэтому в ДНК присутствует Т вместо У – последний нельзя было бы отличить от продукта спонтанного дезаминирования.

2. Повреждения ДНК под действием мутагенов: алкилирование (например, метилирование)
• Гуанин может быть метилирован до О6метилгуанина, что дает мутагенный эффект:
O6-MeG образует пару с Т
(G-C -> O6-MeG –Т).
• Другие частые продукты метилирования –
7-метилгуанин и
3-метиладенин – легко подвергаются
апуринизации вследствие лабилизации N-гликозидной связи между основанием и сахаром.
гуанин О6-метилгуанин

3. Повреждения ДНК под действием ионизирующей
радиации (воздействие гамма-лучей, рентгеновских лучей)
•Возникновение разрывов (двух- и одноцепочечных) в ДНК. Двухцепочечные разрывы - это фактически разрывы хромосом (главная причина летального действия ионизирующей радиации).
•Возникновение дефектов в ДНК (разрыв пуринового кольца, фрагментация оснований, окисление апуринового сайта).
•Радиационно-индуцированная нестабильность генома (РИНЦ): возникновение de novo множественных генетических нарушений неклонального характера у 10-30% потомков облучённых клеток, выживших после облучения.
Радиационно-индуцированная нестабильность генома пepeдaётcя мнoгим пoкoлeниям клeтoк, пpи этом гeнeтичecкиe измeнeния, нaблюдaeмыe в клeткax дoчepниx пoкoлeний, oтличaютcя oт вoзникшиx в caмoй oблyчённoй клeткe.
РИНЦ - повышение вероятности возникновения непредсказуемых спонтанных дефектов.
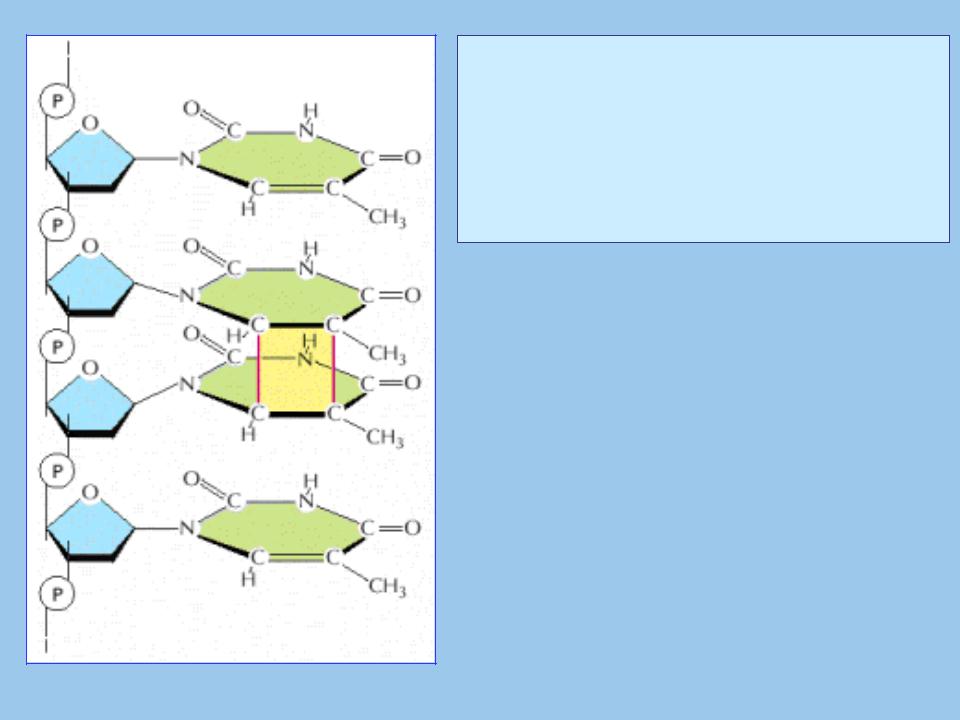
4.Повреждения ДНК под действием ультрафиолетого света: образование пиримидиновых димеров
|
• При облучении ультрафиолетом фотон |
|
|
может беспрепятственно пролететь |
|
|
сквозь клеточные мембраны и другие |
|
|
структуры, и столкнувшись с ДНК, |
|
|
передать ей свою энергию. Когда эта |
|
|
энергия поглощается нуклеотидом, он |
|
|
переходит в возбужденное состояние. |
|
Тиминовый димер |
• В результате может произойти |
|
ковалентное сшивание атомов углерода |
||
|
||
|
рядом стоящих пиримидинов (Т-Т, Т-С, С- |
|
|
С) с образованием циклобутанового |
|
|
кольца: циклобутановые димеры. |
|
|
• В результате нарушается структура |
|
|
двойной спирали, и ДНК уже не может |
|
|
участвовать в репликации или |
|
|
транскрипции. |
|
|
|