Мега Шпора
.doc
|
№ 67 Щитовидная и паращитовидная железы: гормоны, их функции и регуляция деятельности этих желёз.
Щитовидная железа Тироксин (Т4), трийодтиронин, йодсодержащие гликопротеиды.. Обеспечение роста, умственного и физического развития. Стимуляция энергетического обмена, синтеза белка и окислительного катаболизма жиров и углеводов, поглощения кислорода и метаболизма всех клеток. Повышение чувствительности клеток к катехоламинам. Активация натриевого насоса. Стимуляция водного и электролитного обмена. Повышение возбудимости ЦНС. К-клетки щитовидной железы, вилочковой и околощитовидной железы. Тирокальцитонин - Регуляция метаболизма кальция и фосфора; гипокальциемический фактор: подавление активности остеокластов и поступления кальция и фосфора в кровь. Околощитовидная железа Паратгормон - Регуляция метаболизма кальция и фосфора; гиперкальциемический фактор: стимулирует поступление кальция и фосфора из костной ткани в кровь, усиливает реабсорбцию кальция в почке и его всасывание в кишечнике. Щитовидная железа относится к гипофиззависимым от передней доли гипофиза железам. Околощитовидные железы относятся к гипофизнезависимым. |
№ 68 Эндокринная функция поджелудочной железы, ее гормоны и их значение для организма. Регуляция внутрисекреторной деятельности поджелудочной железы.
Поджелудочная железа В-клетки. Инсулин; белок - Регуляция обмена углеводов, липогенез, гликогенез, активация утилизации клетками глюкозы, стимуляция синтеза белка, подавление липолиза, усиление образования жиров. А-клетки. Глюкагон; белок - Стимуляция гликогенолиза и липолиза в печени; усиление секреции адреналина; регуляция секреции инсулина; выраженный катаболический эффект. Поджелудочная железа натощак выделяет небольшое количество панкреатического секрета. Поджелудочный сок богат ферментами, которые синтезируются в ацинозных панкреоцитах. Ферменты поджелудочного сока переваривают все виды питательных веществ. Амилаза, липаза и нуклеазы секретируются поджелудочной железой в активном состоянии, а протеазы – в виде зимогенов. Секреция поджелудочной железы регулируется нервными и гуморальными механизмами. Парасимпатическая регуляция. Раздражение блуждающего нерва вызывает выделение поджелудочного сока, богатого ферментами. Холинергические волокна блуждающих нервов посредством ацетилхолина действуют на М-холинорецепторы панкреацитов. Симпатические волокна, иннервирующие поджелудочную железу, через посредство а-адренорецепторов тормозят поджелудочную секрецию. Адренергические эффекты снижения секреции обеспечиваются также уменьшением кровоснабжения поджелудочной железы путем сужения кровеносных сосудов через их а-адренорецепторы. Гуморальная регуляция. Секретин - стимулирует обильное сокоотделение и секрецию бикарбонатов. Секретин образуется в двенадцатиперстной кишке, а его высвобождение в кровь дуоденальными клетками происходит при переходе в нее кислого желудочного содержимого. Чем больше свободных ионов Н+ в двенадцатиперстной кишке, тем больше высвобождается секретина и тем выше объем панкреатического сока и секреция гидрокарбонатов. Холецистокинин - усиливает секрецию поджелудочной железы, высвобождаясь в кровь из ССК-клеток слизистой оболочки 12-перстной кишки. Секрецию поджелудочной железы усиливают также гастрин, серотонин, инсулин, бомбезин, соли желчных кислот. Тормозят выделение поджелудочного сока глюкагон, соматостатин, вазопрессин, энкефалин, кальцитонин. Вазоинтестинальный пептид может как возбуждать, так и тормозить панкреатическую секрецию. |
№ 69 Эндокринная функция надпочечников. Гормоны мозгового и коркового слоя надпочечников, их значение для функций организма. Участие гормонов надпочечников в приспособительных реакциях организма.
Надпочечники Корковое вещество: гидрокортизон (кортизол), выработка регулируется АКТГ - Регуляция обмена углеводов, белков, жиров, глюконеогенез, катаболическое действие, липолиз, противовоспалительное действие, повышение устойчивости к инфекции. Альдостерон - Регуляция минерального обмена и водно-солевого равновесия, увеличение активного транспорта натрия через клеточные мембраны, повышение реабсорбции натрия и воды в канальцах нефрона:задерживает в огранизме Na и Cl и усиливает выведение калия и аммония. Аналогичное влияние на клетки потовых, слюнных и кишечных желез. Участие в адаптации организма к повышенной температуре окружающей среды. Андрогены; стероиды. Мозговое вещество: адреналин, НА, катехоламины - Стимуляция всех видов обмена веществ, гликонеогенеза, липолиза, термогенное действие. Учащение, усиление сокращений сердца, сужение кровеносных сосудов, расширение бронхов, зрачков. Увеличение вентиляции легких, доставки кислорода к мышцам, сердцу и мозгу. Надпочечники (корковое вещество) относятся к гипофиззависимым от передней доли гипофиза. Тропные гормоны аденогипофиза активируют выделение гормонов железам, которые воздействуют на аденогипофиз и тормозят его активность. Мозговое вещество – гипофизнезависимые. Участие гормонов надпочечников в приспособительных реакциях. Важная роль в регуляции содержания глюкозы в крови в условиях гипогликемии принадлежит гормонам надпочечников. В ответ на пониженное содержание глюкозы в крови в мозговом веществе надпочечников усиливается выработка адреналина. Это является следствием первичного влияния гипогликемии на гипоталамус и гипофиз. Роль гипоталамуса Раздражение рецепторных клеток гипоталамуса приводит к повышению тонуса симпатико-адреналовой системы, что вызывает повышенную секреторную активность мозгового вещества надпочечников и как следствие — увеличенный выброс в кровь адреналина. Последний вместе с глюкагоном активирует фосфорилазу печени и усиливает распад печеночного гликогена. Одновременно усиливается распад гликогена мышц, поэтому после введения адреналина или избыточного его образования увеличивается концентрация глюкозы и молочной кислоты в крови. Роль гипофиза Стимуляция «гипогликемической кровью» гипофиза приводит к дополнительной выработке адренокортикотропного гормона. Избыточное образование адренокортикотропного гормона способствует выделению корковым веществом надпочечников глюкокортикоидов. Повышение концентрации в крови последних приводит к усилению гликонеогенеза — новообразованию глюкозы из неуглеводов, в частности из продуктов расщепления белков и жиров, что, естественно, сопровождается увеличением концентрации глюкозы в крови и содержания гликогена в печени. В мышцах и других тканях одновременно происходит усиленный распад белков, а освобождающиеся аминокислоты используются затем как исходный материал для гликонеогенеза. |
|
|
||
|
|
||
|
№ 70 Эндокринная функция половых желёз. Место образования половых гормонов и регуляция их продукции. Формирование и механизм половой мотивации.
Яичники Эстрогены (эстрадиол, стероиды) - Половая дифференцировка у эмбриона, развитие половых органов, вторичных половых признаков. Обеспечение пролиферативной фазы эпителия слизистой оболочки матки. Анаболическое действие, особенно в периоде полового созревания. Прогестерон; стероид - Подготовка слизистой оболочки матки к имплантации зародыша. Блокирует действие эстрогенов на матку, обеспечивая сохранение беременности. Торможение секреции пролактина. Катаболическое действие. Яички Андрогены (тестростерон, андростерон, стероиды) - Половая дифференцировка эмбриона по мужскому типу, развитие вторичных половых признаков, мужское половое поведение и агрессивность. К гипофиззависимым относят половые железы (яички, яичники). Тропные гормоны аденогипофиза активируют выделение гормонов соответствующими железами, которые в свою очередь в определенных концентрациях воздействуют на аденогипофиз и тормозят его активность. Женские половые органы – яичники – состоят из коркового и мозгового веществ. Гормональную функцию несет корковое вещество. С момента образования полости в фолликуле он начинает синтезировать половые гормоны. Из фолликулярной жидкости выделены три эстрогена: эстрон, эстрадиол, эстриол. Регуляция женских половых желез осуществляется совместным действием ФСГ и ЛГ. Если ФСГ подготавливает морфологические структуры фолликула к синтезу половых гормонов, то ЛГ стимулирует образование из холестерина прегненолона — основного предшественника всех половых гормонов. В женском организме первая фаза менструального цикла, всецело связанная с развитием фолликула в яичниках, регулируется ФСГ. Этот гормон ответствен за формирование гранулезной оболочки фолликула, стимулирует гиперплазию гранулезных клеток и биосинтез ими эстрогенов. В сложном процессе разрыва созревшего фолликула доминирующую роль играет ЛГ. Пролактин вместе с гормоном желтого тела — прогестероном — подавляет в яичниках развитие новых фолликулов. Если произошло оплодотворение, они обеспечивают имплантацию яйцеклетки и формирование плаценты. Эти гормоны ответственны за подготовку репродуктивных органов к родам, они способствуют развитию молочных желез, регулируют лактацию. Основным источником мужских половых гормонов являются яички. Сперматогенез осуществляется в извитых канальцах. Сперматогенез регулируется в основном фолликулостимулирующим гормоном аденогипофиза. Основными гормонами, секретируемыми яичками человека, являются тестостерон, андроапендион и дегидроэпиандростерон. Основная физиологическая роль андрогенов в мужском организме состоит в стимуляции сперматогенеза и развитии вторичных половых признаков. Андрогены образуются не только яичками, но и надпочечниками. В репродуктивном возрасте тестостерон в мужском организме секретируется только семенниками. Андростендион образуется и яичками, и надпочечниками. Эстрадиол образуется только яичками, а эстрон образуется из предшественников, секретируемых корковым веществом надпочечников. В мужском организмк сперматогенез регулируется ФСГ, а биосинтез андрогенов – ЛГ. До периода полового созревания половые гормоны вырабатываются надпочечниками. По достижении полового созревания основную роль по выработке половых гормонов берут на себя половые железы. Гормональный фон создает основу, обеспечивающую половую функцию, направленную на воспроизведение. Гормоны влияют на тканевой метаболизм, на функциональное состояние нейронов в определенных структурах мозга. |
№ 71 Функциональная система, поддерживающая оптимальный для метаболизма уровень глюкозы и крови. Характеристика её узловых механизмов.
Углеводы играют ведущую роль в энергетическом обмене организма. Деятельность практически всех без исключения органов находится в большей или меньшей зависимости от содержания углеводов в притекающей к ним крови. Местные резервы углеводов в разных тканях неодинаковы, поэтому степень зависимости скорости обменных процессов в органах и их функции определяются концентрацией глюкозы в крови. Особенно большое значение глюкоза крови имеет для работы мышц, занимающих в количественном отношении преобладающее место в организме, и деятельности нервной системы благодаря ее ведущей, регулирующей роли в организме. Содержание глюкозы в артериальной крови взрослого человека составляет 4,2—6,4 ммоль/л. В венозной крови содержание глюкозы обычно несколько ниже, чем в артериальной, так как часть ее переходит из крови в ткани в процессе обмена. Артериовенозная разница зависит от специфики органа и уровня его активности: в период активной работы поглощение глюкозы возрастает и артериовенозная разница увеличивается, что косвенно говорит о функциональной активности того или иного органа. Периодические изменения уровня глюкозы в крови у человека и животных обусловлены суточными и сезонными колебаниями, приемом пищи, эмоциональным состоянием и возрастными особенностями. Нормальный уровень глюкозы в крови, так же как и его изменения, воспринимается специальными хеморецепторами, чувствительными к изменению концентрации глюкозы крови. Глюкозорецепторы расположены в печени, сосудах, желудочно-кишечном тракте, центральной нервной системе. Центральные глюкозорецепторы расположены в вентромедиальном отделе гипоталамуса и через рилизинг-факторы оказывают активирующее влияние на передний отдел гипофиза, а через него на деятельность таких желез внутренней секреции, как щитовидная железа, надпочечники и поджелудочная железа. |
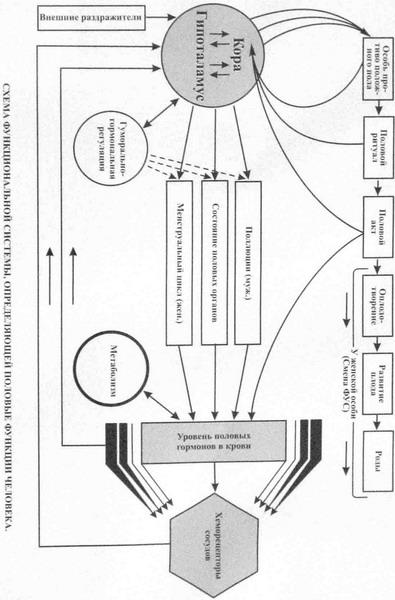
№ 72 Функциональная система, определяющая половые функции организма. Роль биологических и социальных факторов в формировании полового поведения.
Внутренний результат - определенный уровень половых гормонов в крови. К половым гормонам относятся прежде всего мужские половые гормоны — андрогены и женские половые гормоны — эстрогены и прогестины, вырабатываемые половыми железами. По достижении полового созревания основную роль по выработке половых гормонов берут на себя половые железы: семенники у мужских (у человека — яички) и яичники у женских особей. Половые гормоны, кроме специфического участия в организации половых функций, оказывают на организм широкое воздействие, влияя, в частности, на процессы тканевого метаболизма, на функциональное состояние нейронов в определенных структурах мозга и т.д. Выработка половых гормонов в организме определяется не только половыми железами, но находится в тесной зависимости от внешних влияний: в женском организме в форме менструального цикла. Гормональный фон создает как бы основу, обеспечивающую половую функцию, направленную на воспроизведение. Особенностью функциональной системы воспроизведения является то, что она строится на различных гормональных механизмах саморегуляции в женском и мужском организмах. Внешнее звено саморегуляции - механизмы активного взаимодействия особей противоположного пола в плане достижения биологического и социального результата, осуществления полового акта, воспроизведения и продления вида. Результатом деятельности данной системы является поддержание уровня половых гормонов, оптимального для каждого возрастного периода особей. Гормональная и репродуктивная функции мужского и женского организма находятся под контролем сложной нейроэндокринной организации, включающей в себя гипоталамус, гипофиз, периферические железы внутренней секреции. Первым уровнем, где реализуются гормональные эффекты, являются различные органы и ткани. Их клетки, дифференцируясь, приобретают специфические рецепторы к широкому спектру гормонов. Среди межклеточных тканевых регуляторов ведущую роль играют простагландины. Их действие опосредуется через циклический аденозинмонофосфат. Вторым уровнем нейроэндокринной регуляции являются периферические железы внутренней секреции. Продуцируемые ими гормоны, поступая в кровь, обладают дистантным и пролонгированным во времени действием. Воздействуя на территориально разобщенные органы, гормоны способствуют объединению их специфической деятельности. Например, молочная железа и матка обладают чрезвычайно высокой чувствительностью к половым гормонам. Периферические эндокринные железы в свою очередь регулируются тройными гормонами гипофиза — третий уровень. Четвертый уровень составляют гипоталамические центры, которые посредством гормонов контролируют тропные функции аденогипофиза. Пятый уровень — экстрагипоталамические влияния, непосредственно участвующие в регуляции нейросекреторных функций гипоталамических центров. |
|
|
||
|
|
||
|
№ 73 Вегетативная нервная система и ее роль и регуляции различных видов деятельности организма. Особенности строения рефлекторной дуги вегетативного рефлекса. Вегетативные ганглии: локализация, классификация, механизмов передачи возбуждения в ганглиях. Вегетативные рефлексы и их виды, классификация. Адаптационно-трофическая роль вегетативных рефлексов /И.П.Павлов, Л.А.Орбели, А.Г. Гинецинский/.
Вегетативная нервная система — часть общей нервной системы, имеющая отношение преимущественно к регуляции вегетативных (растительных) функций: сокращению гладких мышц, секреции железистого аппарата, функционированию соединительной ткани внутренних органов, тонусу кровеносных и лимфатических сосудов. ВНС отчасти оказывает влияние на деятельность скелетных мышц и наружных половых органов. ВНС часто рассматривается как автономная, так как ее деятельность в слабой степени регулируется произвольно. ВНС характеризуется: • меньшей возбудимостью; • меньшей функциональной лабильностью (не более 5—15 имп/с); • меньшей скоростью проведения возбуждения; • пролонгированным до 150 мс (за счет следовых фаз) потенциалом действия; • временной и пространственной суммацией возбуждений в ганглиях. Эффекторные волокна ВНС иннервируют практически все ткани и внутренние органы. Часть органов имеет «двойную» и «тройную» иннервацию: симпатическую, парасимпатическую и метасимпатическую. В отличие от соматической нервной системы, которая построена по сегментарному принципу, вегетативная нервная система не имеет сегментарной организации. Нейроны вегетативной нервной системы располагаются в различных отделах центральной нервной системы, образуя так называемые вегетативные центры. Вегетативная рефлекторная дуга состоит из трех звеньев, имеющих более сложное строение, чем рефлекторная дуга соматической нервной системы. Рефлекторная дуга ВНС состоит из чувствительного (афферентного, сенсорного), ассоциативного (вставочного) и эффекторного (эфферентного) звеньев. Чувствительное (первое) звено образовано клетками спинномозговых или периферических ганглиев. Ассоциативное (второе) звено представлено преганглионарными нейронами, расположенными в боковых рогах спинного мозга, в продолговатом и в среднем мозге. Отростки вставочных преганглионарных нейронов выходят из спинного мозга в составе вентральных корешков, вступают в соматические нервные стволы и отсюда достигают узлов пограничного симпатического ствола или парасимпатических узлов в органах, где переключаются на эффекторные клетки. Эффекторное (третье) звено образовано эффекторными клетками периферических ганглиев. Периферическая (исполнительная) часть ВНС представлена преганглионарными и ганглионарными нейронами. Смотри №80. Особенности строения симпатических и парасимпатических нервов. Симпатические нервы сформированы в основном преганглионарными, а парасимпатические — постганглионарными нервными волокнами. Преганглионарные волокна симпатической и парасимпатической нервной системы представлены тонкими миелиновыми волокнами типа В, постганглионарые — тонкими безмиелиновыми волокнами типа С. К метасимпатической нервной системе относится комплекс микроганглионарных образований, расположенных в стенках внутренних органов, обладающих двигательной активностью — сердца, бронхов, мочевого пузыря, пищеварительного тракта, матки, желчного пузыря и желчных путей. Нейроны ВНС участвуют в рефлекторных реакциях, называемых вегетативными рефлексами. Некоторые из них используются в качестве функциональных проб — глазосердечный рефлекс, дыхательно-сердечный рефлекс, ортостатическая реакция, кожный дермографизм. Л.А.Орбели, А.Г. Гинецинский обнаружили, что раздражение постганглионарных волокон симпатического нерва восстанавливает сокращение поперечнополосатой мышцы, утомленной раздражением двигательного нерва. Этот эффект обусловлен прямым действием симпатической нервной системы на обмен веществ мышечной ткани и не связан с сосудистыми влияниями. |
№ 74 Симпатический отдел вегетативной нервной системы. Функциональная организация симпатической нервной системы и ее роль в регуляции функций организма. Особенности строения рефлекторной дуги симпатической нервной системы, (расположение центров, локализации ганглиев, виды нервных волокон: медиаторы и рецепторы). Влияние парасимпатической нервной системы на функции организма.
Симпатический отдел. Тела первых нейронов симпатического отдела ВНС расположены преимущественно в задних ядрах гипоталамуса, среднем и продолговатом мозге и в передних рогах спинного мозга, начиная с 1-го грудного и кончая 3—4-м сегментом поясничного ее отдела. Периферическая (исполнительная) часть ВНС представлена преганглионарными и ганглионарными нейронами. Симпатический отдел. Тела преганглионарных нейронов располагаются в сером веществе боковых рогов спинного мозга, начиная от 1-го грудного сегмента до 2—3-го поясничного сегмента, и в двигательных ядрах черепных нервов. Миелиновые аксоны этих нейронов представлены медленнопроводящими возбуждение В-волокнами. Аксоны преганглионарных нейронов через синапсы взаимодействуют с нейронами, расположенными в периферических ганглиях. Околопозвоночные ганглии расположены по обе стороны вдоль позвоночника в виде цепочек, составляющих правый и левый симпатические стволы. Продольно ганглии соединяются межузловыми соединительными ветвями, состоящими из миелиновых и безмиелиновых нервных волокон. Имеются и поперечные связи между ганглиями правой и левой стороны. Кроме этого, ганглии симпатического ствола соединяются со спинномозговыми нервами, благодаря чему они получают возможность иннервировать скелетную мускулатуру. Другая часть постганглионарных волокон направляется к внутренним органам. Особенности строения симпатических и парасимпатических нервов. Симпатические нервы сформированы в основном преганглионарными, а парасимпатические — постганглионарными нервными волокнами. Преганглионарные волокна симпатической и парасимпатической нервной системы представлены тонкими миелиновыми волокнами типа В, постганглионарые — тонкими безмиелиновыми волокнами типа С. Локализация ядер – заднее ядро гипоталамуса, нейроны боковых рогов грудных и поясничных сегментов спинного мозга. Характеристика отростков – прерываются в ганглиях, преганглионарные короткие, тип В, постганглионарные длинные, тип С. Зона иннервации – иннервирует все внутренние органы Локализация – парабертебральные (пограничный столб), превертебральные (чревное, солнечное сплетение, брыжеечные узлы). Медиатор – ацетилхолин, норадреналин. Физиологические свойства симпатической нервной системы: 1.благодаря мультипликации в симпатических ганглиях распространяющееся из них возбуждение широко охватывает сразу несколько различных органов, т.е. является генерализованным. Эти влияния наиболее отчетливо прослеживаются при эмоциональных реакциях. 2.оказывает активирующие влияния на функции иннервируемых органов: усиливает катаболические реакции, силу и частоту сокращений сердца, повышает артериальное давление, улучшает оксигенацию тканей, увеличивает содержания глюкозы в крови, скорость проведения возбуждения в скелетных мышцах и их тонус, расширяет бронхи, увеличивает объем легочной вентиляции; расширяет зрачки, увеличивает секрецию катехоламинов надпочечниками. При этом одновременно снижается тонус пищеварительного тракта, ослабляются процессы всасывания и ферментативного расщепления в кишечнике. 3. участвует в формировании целостных состояний, как агрессия, стресс, болевые реакции. 4. передача с пре- на ганглионарные нейроны осуществляется с помощью ацетилхолина, а на эффекторы — норадреналина. 5. представлена на периферии на значительном протяжении постганглионарными волокнами типа С, возбуждение на пути от ганглиев к органам распространяется медленнее, чем в парасимпатической системе. 6. Эффекты действия более продолжительны. 7. Электрические потенциалы в симпатических ганглиях характеризуются продолжительными следовыми явлениями. При стимуляции преганглионарных волокон регистрируется быстрый возбуждающий постсинаптический потенциал, который сменяется медленным тормозным постсинаптическим потенциалом. 8. начальная деполяризация обусловлена действием ацетилхолина на никотиновые рецепторы. |
№ 75 Парасимпатический отдел вегетативной нервной системы. Структурно-функциональная организация парасимпатической нервной системы и ее роль в регуляции функций в организме Особенности строения рефлекторной дуги парасимпатической нервной системы (расположение центров в ЦНС, локализация ганглиев, медиаторы и рецепторы). Влияние парасимпатической нервной системы на функции различных органов.
Парасимпатический отдел Центральные нейроны расположены в передних отделах гипоталамуса, среднем и продолговатом мозге, во 2-4-м сегментах крестцового отдела спинного мозга. Периферическая (исполнительная) часть ВНС представлена преганглионарными и ганглионарными нейронами. Парасимпатический отдел. Тела преганглионарных нейронов расположены в среднем и продолговатом мозге среди двигательных нейронов черепных нервов: глазодвигательного, лицевого, языкоглоточного и блуждающего, а также в сером веществе боковых рогов крестцового отдела спинного мозга. Вместе с черепными нервами преганглионарные волокна распространяются к ганглионарным нейронам, расположенным в крылонебном, подчелюстном и ушном ганглиях. Отходящие от них постганглионарные волокна иннервируют глазное яблоко, аккомодационную мышцу и сфинктер зрачка, поднижнечелюстную, подъязычную и околоушную слюнные железы. Преганглионарные волокна блуждающего нерва распространяются к нейронам, расположенным в ганглиях органов грудной и верхней части брюшной полости. Аксоны нейронов крестцового отдела спинного мозга через тазовые нервы адресуются к нейронам, располагающимся в ганглиях органов нижней части брюшной полости и таза, иннервируя нижнюю часть толстой кишки, прямую кишку, мочевой пузырь, нижнюю часть мочеточника и наружные половые органы. Особенности строения симпатических и парасимпатических нервов. Симпатические нервы сформированы в основном преганглионарными, а парасимпатические — постганглионарными нервными волокнами. Преганглионарные волокна симпатической и парасимпатической нервной системы представлены тонкими миелиновыми волокнами типа В, постганглионарые — тонкими безмиелиновыми волокнами типа С. Локализация ядер – передний гипоталамус, средний мозг, мост, продолговатый мозг, боковые рога I-V крестцовых сегментов спинного мозга. Характеристика отростков - прерываются в ганглиях, преганглионарные длинные, тип В, постганглионарные короткие, тип С. Зона иннервации – ограничена (нет в надпочечниках и стенках большинства сосудов) Локализация –в иннервируемых органах (интрамурально) или рядом с ними. Медиатор - ацетилхолин Физиологические свойства парасимпатической нервной системы: 1. влияния направлены и локальны. Оказывает ограниченное воздействие в пределах иннервируемого органа. 2. оказывает успокаивающее, расслабляющее действие на большинство функций организма; снижается возбудимость ЦНС и миокарда, уменьшаются интенсивность метаболизма, сила и частота сердечных сокращений, кровяное давление, объем легочной вентиляции, температура тела; увеличивается секреция инсулина и как следствие снижается концентрация глюкозы в крови и увеличивается внутриклеточное депонирование ее в виде гликогена. При этом одновременно усиливаются моторная, секреторная и всасывательная функции желудочно-кишечного тракта. 3. усиливает анаболические реакции. 4.Парасимпатические влияния доминируют в формировании сна и психологического субъективного чувства удовлетворения. 5.Медиатором в преганглионарных и постганглионарных волокнах служит ацетилхолин. 6.Парасимпатическая нервная система представлена на значительном расстоянии преганглионарными волокнами типа В. Вследствие этого возбуждения от центра до органа доходят быстрее, чем по симпатическим нервам. 7.Эффекты действия парасимпатических нервов менее продолжительны. |
|
|
|
|
|
№ 76 Нервный центр. Понятие об анатомическом и функциональном нервном центре. Физиологические свойства и принципы распространения возбуждения по нейронным цепям нервного центра.
Функционально связанная совокупность нейронов, расположенных в одной или нескольких структурах ЦНС и обеспечивающих регуляцию той или иной функции или осуществление целостной реакции организма, называется центром нервной системы. Физиологическое понятие центра нервной системы отличается от анатомического представления о ядре, где близко расположенные нейроны объединяются общими морфологическими особенностями. Классификация нервных центров. По локализации в структурах нервной системы различают корковые, подкорковые и спинальные центры. В головном мозге также выделяют центры диэнцефальные, мезэнцефальные, бульбарные, гипоталамические, таламические. На функциональной основе центры нервной системы разделяют по регулируемой функции (например, сосудодвигательный центр, центр теплообразования, дыхательный центр и др.) или по афферентному восприятию (например, центры зрения, слуха, обоняния и др.). Выделяют также центры нервной системы, которые формируют мотивационные состояния организма, являясь пейсмекерами мотивационных возбуждений (центры голода, жажды, насыщения и др.). Существуют центры нервной системы, которые на основе интеграции возбуждений формируют целостные реакции организма (например, центры глотания, чиханья, дефекации, половой центр и т.п.). Тонус нервных центров. От центров нервной системы идет больший или меньший поток импульсов к эффекторам, обусловливая их постоянное тоническое возбуждение. Тоническое состояние центра нервной системы определяется соотношением активированных и так называемых молчащих нервных клеток, что обеспечивается не только поступающими от периферических рецепторов афферентными импульсами, но и гуморальными влияниями (гормоны, метаболиты, биологически активные вещества). Доминанта. При высоком уровне тонического возбуждения центр может доминировать в деятельности ЦНС (принцип доминанты А.А. Ухтомского). В доминантном центре нервной системы нейроны характеризуются длительной повышенной возбудимостью, возрастает эффективность временной и пространственной суммации возбуждений. Доминирующий центр, подавляя деятельность других центров, может стать главенствующим в формировании целенаправленного поведенческого акта животных и человека. Примером доминанты служит классический опыт А.А. Ухтомского, в котором раздражение сенсомоторной коры на фоне механического раздувания прямой кишки вызывало реакцию дефекации вместо сокращения лапы животного. Линейное распространение возбуждения. Наиболее жесткая линейная связь осуществляется афферентными проекционными путями, обеспечивающими передачу возбуждений от периферических специализированных рецепторов через различные ядра головного мозга вплоть до коры большого мозга. Возбуждение от рецепторов поступает в задние столбы спинного мозга, затем через ядра продолговатого мозга и вентробазальный комплекс таламуса достигает соматосенсорной области коры большого мозга. Генерализованное распространение возбуждения. - обеспечивает функциональное взаимодействие между корой большого мозга и подкорковыми структурами. Эффект восходящих генерализованных активирующих влияний ретикулярных образований проявляется в возбуждении всех областей коры большого мозга, что находит отражение в десинхронизации суммарной биоэлектрической активности мозга. В головном мозге существуют нисходящие, главным образом кортико-фугальные влияния на подкорковые структуры. Эти структуры могут вновь активировать кору мозга восходящими потоками импульсов, создавая эффект возвратной генерализации возбуждений. Взаимодействие восходящих и нисходящих влияний обусловливает двустороннюю связь между структурами головного мозга, особенно между корой мозга и подкорковыми образованиями — реверберацию возбуждений. |
№ 77 Нейрон, его строение и функции. Морфологическая и функциональная классификация нейронов. Представление об интегральной деятельности нейронов (П.К. Анохин). Методы изучения отдельных нейронов.
Нейрон – основная структурная и функциональная единица центральной нервной системы. С позиции об анатомическом, функциональном и генетическом единстве нервной клетки нейрон с его отростками – дендритами и аксоном – является основной структурной единицей нервной системы. Основной функцией нейронов является их 1.способность к возбуждению. Возбуждение может возникать как в результате синаптических влияний на нейрон других нервных клеток, так и за счет эндогенных цитоплазматических процессов. Внешним выражением возбуждения нейрона является колебание электрического потенциала на его мембране. В невозбужденном нейроне регистрируется мембранный потенциал, или потенциал покоя, около —70 мВ. 2. синтез БАВ 3. воспроизведение информации 4. хранение и интеграция информации в пресинаптических окончаниях. 5. в аксоне: аксонный транспорт, генерация электрических импульсов, выделение медиатора. Каждый нейрон синтезирует в своем теле и затем выделяет во всех своих синапсах один и тот же медиатор, поэтому нейроны и ацетилхолиновой передачей возбуждения называются холинергическими, с адреналиновой – адренергическими. Дофаминергические нейроны у млекопитающих находятся в гипоталамусе. Норадренергические нейроны обнаружены в составе среднего мозга, моста и продолговатого мозга. В состав дорсального и медиального ядер продолговатого мозга, моста и среднего мозга входят серотонические нейроны. Интегративная деятельность нейрона: наличие многочисленных специфических хеморецептивных участков на постсинаптических мембранах нейронов позволило сформулировать химическую теорию работы нервных клеток. Электрические импульсы, приходящие к синапсам нейрона через медиаторы, трансформируются в химические процессы на постсинаптической мембране, которые в свою очередь вовлекают в биохимические процессы цитоплазматические и ядерные структуры клетки. Внутриклеточные молекулярные преобразования приходящих к нейрону гетерогенных возбуждений обозначаются как интегративная деятельность нервной клетки. В основе химической теории интегративной деятельности нейрона лежит утверждение о том, что метаболический процесс, развертывающийся в цитоплазме нейрона, закреплен генетически и является специфичным по отношению к отдельным постсинаптическим структурам. Внутринейронная функциональная связь хеморецептивной части постсинаптической мембраны с цитоплазматическими процессами обеспечивается целой группой биологически активных веществ, выполняющих функции универсальных регуляторов клеточного метаболизма. К таким веществам относят циклические пуриновые нуклеотиды, простагландины, гормональные вещества, ионы металлов. Такие медиаторы, как норадреналин, адреналин, дофамин, серотонин, гистамин, специфически активируют мембраносвязанный фермент аденилатциклазу, которая катализирует синтез цАМФ из АТФ. Медиатор ацетилхолин активирует гуанилатциклазу — фермент, катализирующий образование цГМФ из гуанозинтрифосфата. Повышение активности гуанилатциклазы обеспечивается окисью азота (N0). В свою очередь образование окиси азота из аргинина катализируется синтазой окиси азота, которая активируется Са2+, связанным с кальмодулином (регуляторный белок). Наличие кальция в нервной клетке имеет отношение к перераспределению ионов Na+ и К+ в клетке, синтезу и секреции медиаторов, синтезу белка и РНК, аксоплазматическому транспорту. При синаптической активации постсинаптических мембран из них выделяются простагландины, которые изменяют энергетический метаболизм нейронов, участвуют в регуляции возбудимости клетки, секреции медиаторов и гормонов. В молекулярных механизмах интегративной деятельности нейронов большая роль принадлежит эндогенным нейропептидам и так называемым мозгоспецифическим белкам. К эндогенным нейропептидам относятся: тиролиберин, холецистокинин, ангиотензин II, пролактин, вазопрессин. Они могут выступать не только в роли нейромедиаторов, но и в роли нейромодуляторов, т.е. оказывать влияние на высвобождение медиаторов из пресинаптических окончаний и постсинаптическую реакцию. |
№ 78 Торможение в центральной нервной системе. Классификация торможения. Опыты И.М. Сеченова. Тормозные нейроны, их синапсы и медиаторы,
Механизмы торможения проявляются в прекращении или уменьшении активности нервных клеток. В отличие от возбуждения торможение — локальный нераспространяющийся процесс, возникающий на клеточной мембране. Сеченовское торможение. Наличие процесса торможения в ЦНС впервые было показано Сеченовым в 1862 г. в экспериментах на лягушке. Выполняли разрез головного мозга лягушки на уровне зрительных бугров и измеряли время рефлекса отдергивания задней лапы при погружении ее в раствор серной кислоты (метод Тюрка). При наложении на разрез зрительных бугров кристаллика поваренной соли время рефлекса увеличивалось. Прекращение воздействия соли на зрительные бугры приводило к восстановлению исходного времени рефлекторной реакции. Рефлекс отдергивания лапки обусловлен возбуждением спинальных центров. Кристаллик соли, раздражая зрительные бугры, вызывает возбуждение, которое распространяется к спинальным центрам и тормозит их деятельность. И.М. Сеченов пришел к выводу, что торможение является следствием взаимодействия двух и более возбуждений на нейронах ЦНС. В этом случае одно возбуждение неизбежно становится тормозимым, а другое — тормозящим. Подавление одним возбуждением другого происходит как на уровне постсинаптических мембран (постсинаптическое торможение), так и за счет уменьшения эффективности действия возбуждающих синапсов на пресинаптическом уровне (пресинаптическое торможение). Пресинаптическое торможение. Пресинаптическое торможение развивается в пресинаптической части синапса за счет воздействия на его мембрану аксо-аксональных синапсов. В результате как деполяризующего, так и гиперполяризующего воздействия происходит блокирование проведения импульсов возбуждения по пресинаптическим путям к постсинаптической нервной клетке. Постсинаптическое торможение. Наибольшее распространение в ЦНС имеет механизм постсинаптического торможения, которое осуществляется специальными тормозными вставочными нервными клетками (например, клетки Реншоу в спинном мозге или клетки Пуркинье (грушевидные нейроны) в коре мозжечка). Особенность тормозных нервных клеток состоит в том, что в их синапсах имеются медиаторы, вызывающие на постсинаптической мембране нейрона ТПСП (тормозящие постсинаптические потенциалы), т.е. кратковременную гиперполяризацию. Например, для мотонейронов спинного мозга гиперполяризующим медиатором является аминокислота глицин, а для многих нейронов коры большого мозга таким медиатором служит гамма-аминомасляная кислота — ГАМК. Частным случаем постсинаптического является возвратное торможение. Реципрокное торможение. Механизм постсинаптического торможения лежит в основе таких видов торможения, как реципрокное и латеральное. Реципрокное торможение является одним из физиологических механизмов координации деятельности нервных центров. Так, попеременно реципрокно тормозятся в продолговатом мозге центры вдоха и выдоха, прессорный и депрессорный сосудодвигательные центры. Реципрокное торможение проявляется на уровне спинного мозга при осуществлении строго координированных двигательных актов (ходьба, бег, чесание). На уровне сегментов спинного мозга возбуждение группы мотонейронов, вызывающих сокращение мышц-сгибателей, сопровождается реципрокным торможением другой группы мотонейронов, приводящих к расслаблению мышц-разгибателей. Латеральное торможение. Активность нейронов или рецепторов, расположенных рядом с возбужденными нейронами или рецепторами, прекращается. Механизм латерального торможения обеспечивает дискриминаторную способность анализаторов. Так, в слуховом анализаторе латеральное торможение обеспечивает различение частоты звуков, в зрительном анализаторе латеральное торможение резко увеличивает контрастность контуров воспринимаемого изображения, а в тактильном анализаторе способствует дифференцировке двух точек прикосновения. При поступлении возбуждений к синапсам нервной клетки на постси-наптических мембранах могут возникать процессы гиперполяризации. Гиперполяризация приводит к возрастанию критического уровня деполяризации мембраны, следовательно, затрудняет возникновение возбуждения. Такие постсинаптические потенциалы получили название «тормозящие постсинаптические потенциалы» (ТПСП); они возникают в синапсах, где медиатор вызывает гиперполяризацию постсинаптической мембраны. Каждый нейрон синтезирует в своем теле и затем выделяет во всех своих синапсах один и тот же медиатор, поэтому нейроны и ацетилхолиновой передачей возбуждения называются холинергическими, с адреналиновой – адренергическими. К гиперполяризующим медиаторам относят ГАМК глицин. Эти медиаторы взаимодействуя с хеморецепторами постсинаптической мембраны, приводят к развитию ТПСП. |
|
|
||
|
|
||
|
№ 79 Электрофизиологические методы исследований центральной нервной системы. Электроэнцефалография: методы регистрации ЭЭГ, характеристика ритмов ЭЭГ. Значение ЭЭГ для оценки функционального состояния центральной нервной системы. Микроэлектродная техника и микроинофорез, стереотаксическая техника.
Методы исследования функций ЦНС делятся на две группы: 1) непосредственное изучение и 2) опосредованное (косвенное) изучение. Методы непосредственного изучения функций ЦНС подразделяют на морфологические и функциональные. Морфологические методы. К морфологическим методам относятся мак-роанатомическое и микроскопическое исследования строения мозга. Этот принцип лежит в основе метода генетического картирования мозга, позволяющего выявлять функции генов в метаболизме нейронов. К морфологическим методам относят и метод меченых атомов. Сущность его заключается в том, что вводимые в организм радиоактивные вещества интенсивнее проникают в те нервные клетки мозга, которые в данный момент наиболее функционально активны. Функциональные методы: разрушение и раздражение структур ЦНС, стереотаксический метод, электрофизиологические методы. Метод разрушения. Разрушение структур мозга является довольно грубым методом исследования, поскольку повреждаются обширные участки мозговой ткани. Так, повреждение в области продолговатого мозга приводит к нарушению дыхания, глотания, деятельности сердца и изменению тонуса сосудов. В клинике для диагностики повреждений мозга различного происхождения (опухоли, инсульт и др.) у человека используют методы компьютерной рентгенотомографии, эхоэнцефалографии, ядерного магнитного резонанса. Метод раздражения структур мозга позволяет установить пути распространения возбуждения от места раздражения к органу или ткани, функция которых при этом изменяется. В качестве раздражающего фактора чаще всего применяют электрический ток. В эксперименте на животных применяют метод самораздражения различных участков мозга: животное получает возможность посылать раздражение в мозг, замыкая цепь электрического тока и прекращать раздражение, размыкая цепь. Стереотаксический метод введения электродов. Стереотаксические атласы, которые имеют три координатных значения для всех структур мозга, помещенного в пространство трех взаимно перпендикулярных плоскостей — горизонтальной, сагиттальной и фронтальной. Данный метод позволяет не только с высокой точностью вводить электроды в мозг с экспериментальной и диагностической целями, но и направленно воздействовать на отдельные структуры ультразвуком, лазерными или рентгеновскими лучами с лечебной целью, а также проводить нейрохирургические операции. Электрофизиологические методы исследования ЦНС включают анализ как пассивных, так и активных электрических свойств мозга. Электроэнцефалография. Метод регистрации суммарной электрической активности мозга называется электроэнцефалографией, а кривая изменений биопотенциалов мозга — электроэнцефалограммой (ЭЭГ). ЭЭГ регистрируют с помощью электродов, располагаемых на поверхности головы человека. Используют два способа регистрации биопотенциалов: биполярный и монополярный. При биполярном способе регистрируют разность электрических потенциалов между двумя близко расположенными точками на поверхности головы. При монополярном способе регистрируют разность электрических потенциалов между любой точкой на поверхности головы и индифферентной точкой на голове, собственный потенциал которой близок к нулю. Такими точками являются мочки уха, кончик носа, а также поверхность щек. Основными показателями, характеризующими ЭЭГ, являются частота и амплитуда колебаний биопотенциалов, а также фаза и форма колебаний. По частоте и амплитуде колебаний различают несколько видов ритмов в ЭЭГ. Гамма >35 Гц, эмоциональное возбуждение, умственная и физическая деятельность, при нанесении раздражения. Бета 13-30 Гц , эмоциональное возбуждение, умственная и физическая деятельность, при нанесении раздражения. Альфа 8-13 Гц состояние умственного и физического покоя, с закрытыми глазами. Тета 4-8 Гц, сон, умеренные гипоксии, наркоз. Дельта 0,5 – 3,5 глубокий сон, наркоз, гипоксия. Основным и наиболее характерным ритмом является альфа-ритм. В состоянии относительного покоя альфа-ритм наиболее выражен в затылочных, затылочно-височных и затылочно-теменных областях головного мозга. При кратковременном действии раздражителей, например света или звука, появляется бета-ритм. Бета- и гамма-ритмы отражают активированное состояние структур головного мозга, тета-ритм чаще связан с эмоциональным состоянием организма. Дельта-ритм указывает на снижение функционального уровня коры большого мозга, связанное, например, с состоянием легкого сна или утомлением. Локальное появление дельта-ритма в какой-либо области коры мозга указывает на наличие в ней патологического очага. Микроэлектродный метод. Регистрация электрических процессов в отдельных нервных клетках. Микроэлектроды - стеклянные или металлические. Стеклянные микропипетки заполняют раствором электролита, чаще всего концентрированным раствором хлорида натрия или калия. Существуют два способа регистрации клеточной электрической активности: внутриклеточный и внеклеточный. При внутриклеточном расположении микроэлектрода регистрируют мембранный потенциал, или потенциал покоя нейрона, постсинаптические потенциалы — возбуждающий и тормозящий, а также потенциал действия. Внеклеточный микроэлектрод регистрирует только положительную часть потенциала действия. |
№ 80 Мышечный тонус. Роль различных отделов центральной нервной системы в механизме поддержании и регуляции мышечного тонуса.
Скелетные мышцы всегда находятся в состоянии некоторого напряжения. Постоянное незначительное напряжение мышц, не сопровождающееся признаками утомления, называется мышечным тонусом. Односторонняя перерезка у спинальной лягушки, подвешенной на крючке штатива, чувствительных (задних) корешков спинного мозга, в которых проходят афферентные нервные волокна, иннервирующие соответствующую заднюю лапку, приводит к исчезновению мышечного тонуса этой лапки и она распрямляется. К аналогичному эффекту приводит перерезка передних (двигательных) корешков или разрушение спинного мозга. Эти опыты свидетельствуют о том, что при разрушении основных звеньев рефлекторного кольца (афферентных и эфферентных путей, нервных центров) мышечный тонус исчезает. Следовательно, мышечный тонус имеет рефлекторную природу. Источником возбуждений, поддерживающих мышечный тонус, являются проприорецепторы. В скелетных мышцах имеются три вида проприорецепторов: • мышечные веретена, расположенные среди мышечных волокон; • сухожильные рецепторы Гольджи, расположенные в сухожилиях; • пачиниевы тельца, расположенные в фасциях, сухожилиях, связках. Особое значение в регуляции мышечного тонуса имеют мышечные веретена и сухожильные рецепторы Гольджи. Мышечные веретена представляют собой небольшие продолговатые образования, напоминающие своим внешним видом прядильные капсулы мышечного веретена находится пучок мышечных волокон, которые называются интрафузальными, т. к. они расположенными внутри веретена в отличие от обычных мышечных волокон, которые называются зкстрафузальными. Каждое интрафузальное волокно состоит из трех частей: • его центральная часть называется ядерной сумкой, в которой находятся ядра мышечной клетки; • два периферических участка, которые имеют поперечную исчерченность и обладают способностью сокращаться; • миотрубки, расположенные между ядерной сумкой и периферическими участками. Ядерную сумку в виде спирали окружают нервные волокна чувствительного нейрона-первичные рецепторные окончания. В области миотрубок нервные окончания афферентных нейронов гроздевидно ветвятся, образуя вторичные рецепторные окончания. В мышце мышечное веретено одним концом прикрепляется к экстрафузальному мышечному волокну, а другим - к сухожилию этого волокна. Таким образом, мышечное веретено расположено в мышце параллельно экстрафузальным мышечным волокнам. При снижении тонуса экстрафузального волокна увеличивается его длина, что приводит к растяжению и раздражению первичных и вторичных рецепторных окончаний, для которых растяжение является адекватным раздражителем. Возбуждение от рецепторных окончаний по афферентным волокнам поступает в спинной мозг к мотонейронам, расположенным в передних рогах. Мотонейроны спинного мозга принято подразделять на альфа- и гамма-мотонейроны (так как их аксоны относятся к А-альфа и А-гамма нервным волокнам). Возбуждение от альфа-мотонейронов поступает к экстрафузальным мышечным волокнам, вызывая их сокращение - тонус восстанавливается. Избыточное сокращение экстрафузальных мышечных волокон приводит к растяжению сухожильных рецепторов Гольджи, так как они прикрепляются к мышце последовательно. В них возникает возбуждение, которое поступает к тормозным вставочным нейронам спинного мозга, а от них к альфа-мотонейронам. Активность альфа-мотонейронов при этом снижается, уменьшается импульсация, идущая от них к экстрафузальным мышечным волокнам, тонус несколько снижается. Рассмотренные выше механизмы поддержания мышечного тонуса осуществляются на уровне спинного мозга, поэтому такой тонус называется спинальным или простейшим. Спинальный тонус характеризуется очень слабой выраженностью тонического напряжения. Такой тонус не может обеспечить поддержание позы животного и акт ходьбы, но он достаточен для осуществления простейших спинальных рефлексов. Большое значение в регуляции мышечного тонуса имеют базальные ядра - бледный шар и полосатое тело, которые образуют стриопаллидарную систему. Эти структуры регулируют активность всех нижележащих отделов ЦНС, участвующих в регуляции мышечного тонуса, обеспечивая адекватное перераспределение тонуса мышц при различных видах деятельности. При поражении экстрапирамидной системы, составной частью которой являются базальные ядра, возникают нарушения регуляций тонуса мускулатуры, что приводит к развитию так называемых дрожательных параличей. Главную роль в приспособительной регуляции мышечного тонуса выполняет кора головного мозга. С ее участием и участием других структур, регулирующих мышечный тонус, формируется нормальный или корковый тонус. Регуляция тонуса скелетной мускулатуры осуществляется экстрапирамидной системой головного мозга. Быстрые (фазные) движения обеспечиваются активностью пирамидной системы. |
№ 81 Спинной мозг. Его строение и функции. Проводящие пути спинного мозга Рефлексы спинного мозга, их виды и строение рефлекторных дуг. Нисходящий контроль деятельности спинного мозга.
Спинной мозг располагается в позвоночном канале и на уровне нижнего края большого затылочного отверстия переходит в головной мозг. В этом месте из спинного мозга (верхняя его граница) выходят корешки, образующие правый и левый спинномозговые нервы. Нижняя граница спинного мозга соответствует уровню I—II поясничных позвонков. Интегративная деятельность спинного мозга связана со структурной организацией его сегментов. Каждый сегмент спинного мозга получает афферентную импульсацию через задние (чувствительные) корешки и посылает возбуждение к эффекторам через передние (двигательные) корешки. Таким образом, уже в пределах каждого сегмента спинного мозга возможна интеграция простейших двигательных реакций. Миотатический рефлекс — одна из простых реакций на растяжение мышцы, осуществляемая на уровне спинного мозга по самому короткому пути — от проприорецепторов до мотонейронов и далее к двигательным мышечным волокнам. Реакция растяжения является основой регуляции длины мышцы, изменение которой возникает при поднятии груза или при сокращении мышцы-антагониста. Сухожильные рефлексы в противоположность реакции на растяжение мышцы способствуют быстрому ее расслаблению. Они защищают ее от повреждения при сильных сокращениях и участвуют в регуляции напряжения в мышце. Когда напряжение достигает значения, при котором возбуждаются сухожильные рецепторы Гольджи, альфа-мотонейроны, иннервирующие сокращающуюся мышцу, тормозятся, что приводит к ее расслаблению и падению напряжения в ней. Стабилизируя подвижные звенья скелета, миотатические и сухожильные рефлексы создают основу для осуществления непроизвольных и произвольных движений. Защитные реакции спинного мозга. На уровне сегментов спинного мозга, возникают при раздражении кожной поверхности повреждающими раздражителями. При раздражении рецепторов кожи происходит отдергивание конечности от раздражителя (сгибательный рефлекс). При этом сокращение мышц – сгибателей сопровождается расслаблением антагонистов мышц – разгибателей. Эти реакции осуществляются сегментарными механизмами реципрокного торможения. При возбуждении мотонейронов какой-либо мышцы двигательные нейроны мышцы-антагониста тормозятся за счет влияния на них вставочных нейронов (клетки Реншоу), выделяющих в синаптических окончаниях тормозной медиатор. Влияние вышележащих отделов ЦНС. Спинальные двигательные центры находятся под контролирующим влиянием вышележащих отделов ЦНС. Ретикулоспинальный и вестибулоспинальные тракты моносинаптически возбуждают мотонейроны проксимальных мышц конечностей и туловища. Мотонейроны дистальных мышц конечностей (особенно мышц пальцев кисти и стопы) находятся под нисходящим влиянием кортико-спинального и руброспинального трактов. Восходящие пути. В сегментах спинного мозга имеются нейроны, дающие начало восходящим афферентным (чувствительным) путям. Среди них в первую очередь выделяют путь Голля и Бурдаха (главным образом тактильное чувство), спинномозжечковые пути (мышечно-суставное чувство) и спиноталамический путь (болевая и температурная чувствительность). Рефлексы делятся на: 1) экстероцептивные (возникают при раздражении агентами внешней среды сенсорных раздражителей) 2) интероцептивные (возникают при раздражении механо-, хемо-, терморецепторов): висцеро-висцеральные—рефлексы с одного внутреннего органа на другой, висцеро-мышечные — рефлексы с внутренних органов на скелетную мускулатуру; 3) проприоцептивные (собственные) рефлексы с самой мышцы и связанных с ней образований. Они имеют моносинаптическую рефлекторную дугу. Проприоцептивные рефлексы регулируют двигательную активность за счет сухожильных и позотонических рефлексов. Сухожильные рефлексы (коленный, ахиллов, с трехглавой мышцы плеча и т. д.) возникают при растяжении мышц и вызывают расслабление или сокращение мышцы, возникают при каждом мышечном движении; 4) позотонические рефлексы (возникают при возбуждении вестибулярных рецепторов при изменении скорости движения и положения головы по отношению к туловищу, что приводит к перераспределению тонуса мышц (повышению тонуса разгибателей и уменьшению сгибателей) и обеспечивает равновесие тела). |
|
|
||
|
|