

Класс I. Мшанки (bryozoa)
Класс мшанок содержит водных, преимущественно морских, реже пресноводных животных, ведущих сидячий образ жизни и образующих в результате бесполого размножения колонии. Чаще всего колонии бывают древовидными, ветвистыми. В некоторых случаях (Flustra) ветви колонии сплющиваются в одной плоскости и принимают характер пластинок, а сама колония становится похожей на пучок листьев. Если же колонии стелются по субстрату, то образуют на дне или на стеблях водорослей тонкие корочки. Каждая колония объединяет большое количество особей. Таким образом, по внешнему виду колонии мшанок больше всего напоминают гидроидных полипов. Размеры мшанок невелики: колонии измеряются несколькими сантиметрами, отдельные особи большей частью длиной не более 1 мм. Современных видов мшанок около 4000, ископаемых еще больше.
Для понимания природы мшанок следует рассмотреть строение отдельной особи.
Строение и физиология. Отдельные особи колонии мшанок (рис. 496) отчетливо подразделяются на два отдела, которые, однако, не являются сегментами. Передний конец особи свободно торчит в окружающую воду и несет на себе рот с венчиком щупалец вокруг. Задняя половина животного одета утолщенной кутикулой, производимой наружным эпителием тела, и имеет вид глубокой чашечки или мешочка. Эту часть животного нередко обозначают особым термином — цистид. Передняя половина мшанки лишена плотной кутикулы, нежная и при раздражении втягивается внутрь цистида; ее называют полипидом (рис. 497). Втягивание совершается при помощи двух особых мускулов-ретракторов. Они находятся по бокам кишечника и передним концом прикрепляются изнутри к стенке тела полипида по сторонам рта, а задним концом — к основанию цистида. При втягивании передней половины тела в цистид отдельные щупальца не вворачиваются, а весь венчик их уходит в глубь образующегося впячивання. Щупальца имеют вид длинных нежных выростов, в которые заходят каналы целома, и покрыты мерцательным эпителием;
502

Рис. 495. Колонии морских мшанок подкласса Gymnolaemata. А — Idmonea tu-mida; Б — Crisia eburnea; В — Alcyonidium mamillatum; Г — Dendrobaenia flustrot-des; Д — участок колонии Dendrobaenia flustroides; E — Porella saccata: 1 — ГОНОЗОИД. 2 — авикулярия

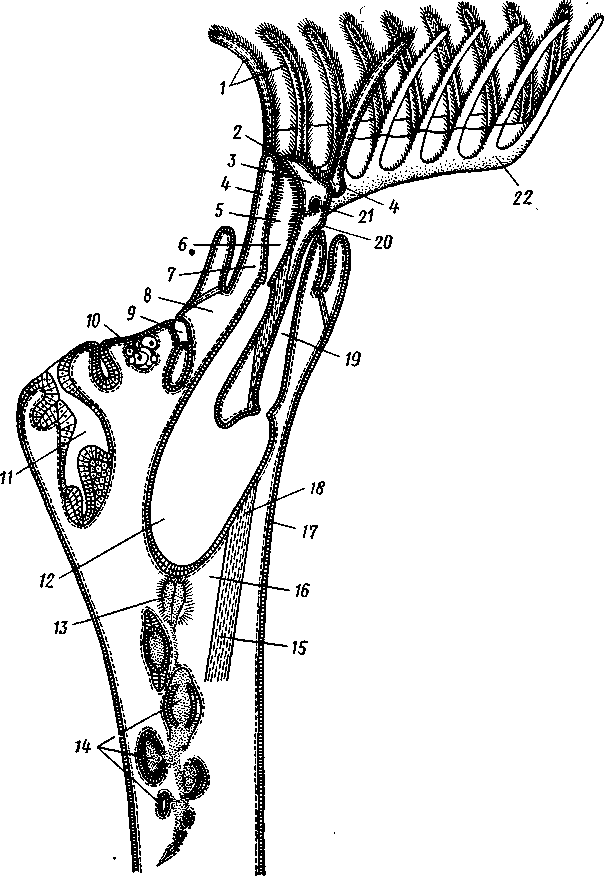
Рис. 496. Участок колонии Plumatella repens (из Матвеева)?
/ — полипид с расправленным лофофором, 1 — передний отдел кишечника, 3 — задняя кишка, 4 — желудок, 5 — стенка цистида, 6 — канатик со статобластамн.
7 — втянутый полипид
503
они служат, с одной стороны, для собирания пищевых частиц, с другой — в качестве органов дыхания.
У пресноводных мшанок, образующих подкласс Phylactolaemata (Покрыторотые), щупальца в два ряда располагаются на особом щу-пальценосце — лофофоре, имеющем сверху, со стороны рта, форму под-ко.вы, у вершины которой и располагается ротовое отверстие (рис. 496, 497). Последнее прикрыто небольшим язычковидным выростом — эпи-стомом — предротовым отделом тела (рис. 497).
Представители подкласса Gymnolaemata (см. рис. 495) (Голоротые мшанки) вторично утратили эпистом, а их щупальце вокруг ротового отверстия образуют простой венчик (рис. 498).
Кутикула цистида может быть различной консистенции: в виде тонкой хитиновой пленки или остуденяется и превращается в толстый желеобразный слой, содержащий до 99% воды. В других случаях кутикула
Рис.
497. Строение особи пресноводной
мшанки из подкл.
Phylactolaemata
(из
Ремане):
I
—
щупальца, 2
— эпистом.
3
—* целом
эпистома, 4
— целом
ло-фофора,
S—глотка,
6
— пищевод,
7 — полипид, 8
— цистид,
9
— оэций, 10
— яичник,
11
—
почка,
12
— желудок,
13
— семенник,
14
— статобласт,
15
—
канатик,
16
— туловищный
целом,
17
— целомический
эпителий,
18
— ретрактор,
19
— задняя
кишка, 20
— анальное
отверстие,
21
—
ганглий, 22
—
ло* фофор
504
ID
11
12

505
I
/ — отверстие цистида, 2 и 3 — части устьевой области цистида, 4 — венчик щупалец, втянутый внутрь цистида, 5 — анус, б — задняя кишка, 7 — мускульные волокна, пересекающие целом, 8, 9 — желудок, 10 — пищевод, // — канатик, 12 — перитонеаль-ный эпителий, 13 — стенка тела, 14 — кутикула, 15 — мускул-ретрактор, 16 — семенники, 17 — яичник, 18 — глотка, 19 — нервный узел, 20 — щупальца
утолщается и пропитывается углекислой известью, становясь очень твердой.
Наружный эпителий мшанок однослоен и у пресноводных форм (подкл. Phylactolaemata) подстилается двумя слабо развитыми слоями мышц: наружным из кольцевых и внутренним из продольных мышечных волокон. У морских голоротых мшанок (подкл. Gymnolaemata) мышечные слои отсутствуют. Слабое развитие кожно-мускульного мешка объясняется неподвижностью мшанок. Перитонеальный эпителий ограничивает обширную вторичную полость тела, в которой помещаются все внутренности.
Полость тела (целом) тонкими перегородками делится на три отдела. Передний обладает небольшими размерами и залегает в эпистоме. Средний — кольцевой канал — окружает глотку и посылает слепые ответвления в щупальца. Задний, наиболее обширный, занимает почти все тело и называется туловищным целомом. Как уже говорилось, у голоротых мшанок эпистом вместе с целомом редуцируется.
Пищеварительная система. Пищеварительный канал имеет характерную для многих сидячих животных подковообразную форму (см. рис. 496, 497, 498). Рот, лежащий внутри венчика щупалец (у подкл. Phylactotaemata— в средней части подковы лофофора, между двумя рядами ее щупалец), ведет сначала в небольшую глотку, а затем в длинную узкую трубку — пищевод. Пищевод переходит в мешковидный, V-образно изогнутый желудок; он впадает в одно из колен желудка, тогда как от другого колена отходит поднимающаяся кпереди тонкая кишка, открывающаяся наружу порошицей непосредственно позади венчика щупалец.
Весь кишечник снаружи одет перитонеальным эпителием. От дна желудка отходит продолжение его перитонеальной выстилки в виде тонкой брыжейки, или канатика, тянущегося через полость тела назад и переходящего затем в наружный, пристенный слой перитонеума.
Пища — мелкие одноклеточные водоросли, простейшие, коловратки и т. п. — подгоняется ко рту мерцанием ресниц на щупальцах. В глотке пища сбивается в пищевые комки и поступает через пищевод в желудок.
Нервная система состоит из единственного надглоточного ганглия (см. рис. 497, 498), лежащего между глоткой и задней кишкой, от которого расходятся в разные стороны, но главным образом к щупальцам, периферические нервы. Единственными органами чувств являются особые осязательные волоски на наружной стороне щупалец.
Интересно, что во время почкования ганглий почки возникает в виде ясного впячивания эктодермы, отшнуровывающегося от покровов; полость впячивания позднее исчезает, и ганглий становится массивным.
Дыхание происходит через всю поверхность тела, но главным образом через щупальца.
Кровеносной системы у мшанок нет. Отсутствие ее, вероятно, объясняется редукцией, вызванной малыми размерами тела, что, в свою очередь, связано с тем, что мшанки — животные колониальные.
Выделительные органы у мшанок отсутствуют. Удаление продуктов обмена из организма осуществляется с помощью клеток-фагоцитов через стенки щупалец или стенку кишечника. Из последнего они выводятся вместе с экскрементами.
У покрыторотых мшанок полости целомов соединяются трубчатыми образованиями, которые рассматриваются как видоизмененные целомо-дукты. Долгое время их считали органами выделения покрыторотых мшанок, но сейчас эта точка зрения оставлена.
Половая система. Мшанки — гермафродиты. Половые клетки раз-
506
виваются под перитонеальным эпителием, на стенке тела или на канатике. Живчики образуют на канатике довольно большое скопление — семенник. Яйца формируются на внутренней стенке тела (рис. 498) в небольшом количестве, иногда по одному. Оплодотворение внутреннее — подвижные живчики выходят из одной колонии мшанок и активно проникают в другую, где в это время имеются созревшие яйца. Выход оплодотворенных яиц наружу совершается разными способами. У одних видов для этого используется специальная целомическая пора, расположенная между щупальцами и ротовым отверстием. Значительно чаще развивается специальный ресничный канал, который открывается наружу непосредственно между щупальцами. Наконец, как полагают, у ряда видов зрелые яйца обладают активным амебоидным движением и сами прокладывают себе дорогу через стенки родительской особи.
У очень немногих видов оплодотворенные яйца выбрасываются в воду, где и осуществляется их дальнейшее развитие. Более обычно проявление у мшанок своеобразной «заботы о потомстве», когда яйца приклеиваются к покровам или даже поступают в особые образования — оэции (см. ниже), в которых протекает их развитие до полного формирования личинки.
Колонии. У мшанок, как и у многих других колониальных организмов, наблюдается морфологическая и функциональная дифференци-ровка особей, образующих колонию. У сохранивших большое количество примитивных признаков покрыторотых мшанок в колониях представлены особи только двух типов. Основную массу составляют обычные индивиды. Кроме них имеются еще оэции, которые образуются за счет молодых почек, развитие которых рано останавливается. Почка у Phy-lactolaemata сначала представляет небольшое впячивание покровов, в полость которого и поступает яйцо. Оэции с развивающимся зародышем до конца сохраняет вид простого мешочка. В колониях подкл. Gymnolae-mata дифференциация особей выражена значительно сильнее. Оэции го-лоротых мшанок имеют вид выступающих бугорков, несущих полость, сообщающуюся с внешней средой. В этой полости протекает формирование личинки. У некоторых видов отдельные особи колонии резко увеличиваются в размерах и принимают характерную кувшинообразную форму. Они называются гонозоидами (см. рис. 495). В полости гонозоидов яйца претерпевают сложное развитие, включающее полиэмбрионию (с. 509).
Ряд особей колонии специализируется для выполнения защитных функций. Это в первую очередь авикулярии (рис. 499). Последние имеют вид, сходный с птичьей головой (с шеей и мощным клювом). «Голова» представляет собой цистид, измененный и вытянутый в неподвижный отросток, отвечающий верхней части «клюва». Кроме того, на «голове» сидит подвижный кутикулярный крючок, отвечающий нижней части клюва; он способен то прижиматься к неподвижному отростку, то отводиться от него. Подвижный кутикулярный крючок есть результат видоизменения крышечки, имеющейся у многих мшанок на переднем крае цистида и захлопывающейся при втягивании полипида. Авикулярии захватывают мелких животных, заползающих на колонию, и убивают или прогоняют их. Разновидностью авикулярии являются вибракулы, которые отличаются отсутствием неподвижного выроста цистида, тогда как кутикулярный крючок вытягивается в длинный подвижный жгут.
Размножение. Мшанки имеют половое размножение, но, кроме того, отличаются большим разнообразием бесполых способов размножения. Прежде всего образование колоний — результат не доведенного до конца бесполого размножения посредством почкования. Детали процесса
507

В результате такого наружного почкования происходит постепенный рост колонии.

Рис. 499. Авикулярия (А) и вибракула (Б) мшанки (из
Догеля):
/ — неподвижная ветвь, 2 — подвижная ветвь, 3, 4—мышцы, раскрывающие и закрывающие «клюв», 5 — подвижный бич вибракулы, приводимый в действие мышцами (6)
Рис. 500. Статобласт Crista-' tella mucedo. Вокруг цент-' ральной массы видно плава-• тельное кольцо, состоящее > из хитиновых камер, наполненных газом, и хитиноид-ные крючочки (по Догелю)
Наряду с наружным почкованием у пресноводных мшанок (отр. Phylactolaemata) имеется еще особый способ внутреннего почкования при помощи образования статобластов (см. рис. 497; 500). Статоблас-ты — особые чечевицеобразные многоклеточные тельца, одетые плотной оболочкой; образуются преимущественно осенью. Зачаток статобласта появляется внутри канатика в виде кучки мезодермальных клеток. Одновременно с этим группа эктодермальных клеток мигрирует с поверхности тела внутрь канатика. Здесь за их счет формируется двухслойная эпителиальная оболочка, окружающая группу мезодермальных клеток. Эпителий выделяет на своей поверхности тонкую, но плотную скорлупу, состоящую из двух слоев, между которыми располагается слой воздухоносных камер. У некоторых видов по краю чечевицы образуется хитиновое кольцо с крючками, или зацепками. Статобласты освобождаются при разрушении материнского тела (например, зимой при отмирании колонии) и попадают наружу. Воздухоносные камеры позволяют им плавать в воде, а своими крючками они зацепляются за водоросли, неровности дна и т. п. Статобласты остаются зимой в покоящемся состоянии,
508
весной же оболочки их лопаются, клеточное содержимое прикрепляется ко дну и образует при помощи наружного почкования новую колонию.
Легко видеть, что внутреннее почкование мшанок имеет большое сходство и одинаковое биологическое значение с образованием геммул у пресноводных губок (с. 108), Наконец, у некоторых мшанок, относящихся к подкл. Gymnolaemata, зимуют особые «зимние» наружные почки: они представляют собой одетые плотной оболочкой отдельные цистиды. От статобластов они легко отличаются по присутствию в них зачатка кишечника, мышц и половых клеток, тогда как внутренность статобласта занята сплошной массой мезодермальных клеток.
Развитие. Все мшанки обладают половым размножением. Дробление яйца полное и во многих случаях почти равномерное. В результате возникает бластула, часто сплющенная в одном направлении. В этом случае она сначала имеет вид пластинки, а позднее становится похожей на чечевицу.
Часть клеток на одном полюсе бластулы внедряется в бластоцель и дает начало неразделенному зачатку энтодермы и мезодермы.
У многих видов, развитие эмбрионов которых протекает в оэции или гонозоиде, между формирующимся зародышем и материнской колонией устанавливается тесная связь в виде своеобразной «плаценты». Таким способом эмбрион получает необходимые для развития питательные вещества.
Процессы эмбрионального развития части мшанок из подкл. Gymnolaemata усложняются за счет появления полиэмбрионии. Оплодотворенное яйцо, развивающееся в гонозоиде, в результате неравномерного дробления дает сначала крупного первичного зародыша, от которого позднее отделяются более мелкие вторичные зародыши. Количество последних может достигать ста и более. В результате развития получаются свободноплавающие личинки неодинакового вида у разных мшанок (рис. 501). Наиболее типичная личинка, сохранившая ряд примитивных признаков,— цифонаут; тело ее заключено в двустворчатую раковину. Передвигается личинка с помощью венчика ресничек. Имеется хорошо развитый кишечник; на переднем конце перед ротовым отверстием располагается особый грушевидный орган, выполняющий функции органа чувств, а на брюшной поверхности — присоска, с помощью которой личинка прикрепляется к субстрату во время метаморфоза.
У многих мшанок наблюдается вторичное упрощение личинок, связанное с утратой раковины и кишечника. Самостоятельный энтодермальный зачаток в этом случае не обособляется.
После периода свободного плавания личинка садится на дно, прикрепляется и испытывает превращение. Тело осевшей личинки принимает вид овального мешочка, и большинство личиночных органов подвергается распаду, которому содействуют амебоидные клетки фагоцитарного характера. В частности, у цифонаута полностью разрушается энтодермальный кишечник. Новый кишечник формируется за счет экто-дермальных клеток. На свободной стороне личинки, противолежащей месту прикрепления, начинается закладка одной или двух первых особей будущей колонии. Они образуются в виде утолщений эктодермы, которые впячиваются внутрь и дают особь мшанки по способу, напоминающему наружное почкование. Путем дальнейшего почкования первых особей и их потомков постепенно вырастает целая колония.
У пресноводных мшанок (подкл. Phylactolaemata) закладка отдельных особей происходит очень рано — еще во время развития личинки. У Cristatella личинки, выходящие из оэция в воду, несут до 20 особей, находящихся на разных стадиях формирования.
509
Экология. Мшанки живут преимущественно в море, где встречаются на различных глубинах, начиная с прибрежных камней и до больших глубин.
В пресных водах встречаются почти исключительно снабженные щупальценосцем Phylactolaemata. Наиболее обыкновенны из них виды рода Plumatella, образующие стелющиеся по субстрату, ветвящиеся колонии или массивные наросты на подводных предметах, и Cristatella mucedo (рис. 502). Последняя особенно интересна тем, что у нее проявляется высокая степень интеграции колонии, выражающаяся в слиянии цистидов. Колонии Cristatella имеют вид студенистой колбаски, на верхней поверхности которой торчат полипиды. Подошва не прикреплена к субстрату, и вся колония может очень медленно ползать.
Срок жизни пресноводных колоний обычно не больше 5—6 месяцев; колонии редко перезимовывают. Жизнь отдельных особей колоний значительно короче; на колонии всегда можно встретить дегенерирующие особи, места которых впоследствии занимаются индивидами, вновь развивающимися из почек.

Рис. 501. Личинки мшанок (из разных авторов). А—личинка типа цифонаут; Б — личинка Bugula sp.; В — личинка Crisia sp.; Г — личинка Plumatella до выворачивания полипидов:
1 — энтодермальная средняя кишка, 2—анальное отверстие, 3 — створки, 4 — присоска, 5 — грушевидный орган, 6 — глотка, 7 — втянутые полипиды
510
Древнейшие морские мшанки известны уже из силурийских отложений.
Практическое значение мшанок мало. Наряду с другими беспозвоночными животными мшанки составляют важный компонент подводных обрастаний. Кроме того, в водопроводах, не снабженных хорошими песчаными фильтрами, мшанки могут причинять существенными вред, заку-

Рис. 502, Общий вид колонии Cristatella mucedo (из Деляжа)
поривая трубы и давая приют разным мелким организмам, загрязняющим воду.
Классификация. Класс Bryozoa, делится на 2 подкласса.
Подкласс 1. Покрыторотые (Phylactolaemata) — пресноводные мшанки с подковообразным лофофором, эпистомом и парой целомодуктов. Представители: Cristatella, Pluma-tella (см. рис. 496, 497) и др.
Подкласс 2. Голоротые (Gymnolaemata)— преимущественно морские мшанки с щупальцами, расположенными по кругу, без эпистома. Иногда имеется рудиментарный цело-
модукт в виде трубочки или поры на спинной стороне. Представители: Crisia, Alcyonidi-
ит, Dendjobaenia, Porella и др. (см. рис. 495, 498).
КЛАСС II. ПЛЕЧЕНОГИЕ (BRACHIOPODA)
Плеченогие образуют небольшой класс одиночных морских животных, ведущих прикрепленный, неподвижный образ жизни. В настоящее время весь класс содержит всего около 280 видов, но ископаемых плече-ногих, главным образом палеозойских, насчитывается более 10000 видов. Размеры большинства современных плеченогих измеряются несколькими миллиметрами или сантиметрами, длина створок раковины самой крупной формы Magellania venosa достигает 8 см.
Строение и физиология. Все тело скрыто в двустворчатой раковине, похожей на таковую пластинчатожаберных моллюсков (рис. 503). Створки раковины выделяются особыми кожными складками, которые называют мантийными складками. Существенное отличие от пластинчатожаберных моллюсков состоит в том, что у них створки охватывают тело с боков, у плеченогих же они прикрывают его со спинной и брюшной сторон. Брюшная створка обыкновенно более крупная и выпуклая, спинная более плоская, иногда прилегает к брюшной в виде крышечки. Каждая створка билатерально симметрична и имеет приблизительно
511
яйцевидную форму (рис. 504). Заостренными задними концами створки обращены книзу, к субстрату; они соединяются друг с другом только при помощи мускулов (рис. 505) или же, кроме того, при помощи замка— совокупности нескольких мелких зубчиков, имеющихся на заднем крае брюшной створки и входящих в соответствующие им ямки на крае противоположной створки.
Соответственно отсутствию или присутствию замка плеченогие делятся на два подкласса: беззамковых (Ecardines, или Inarticulata) и

Рис. 503. Плеченогие в естественной обстановке. Видны стебельки, которыми животные прикрепляются к субстрату (из Джордана и Хет)
Рис. 504. Раковина Magellania flavescens. A —
спинная створка с внутренней стороны; Б—
брюшная створка с внутренней стороны (по Да-
видсопу):
/ — скелет руки, 2 — замочная пластинка, 3 — ямка замка, 4 — отпечатки мускулов, закрывающих и открывающих раковину, 5 — зубец замка, 6 — отверстие для стебелька
Рис.
505. Мускулатура и аппарат рук Magellania
flavescens (по
Дэвидсону):
/
— брюшная мантийная
складка,
1
— щупальца,
J
— рука,
4
— передняя
стенка тела, 5
— мускулы-открыватели
раковины. 6
—
мускулы-замыкатели раковины, 7
— целом. 8
— брюшная
створка раковины. 9 — отверстие
для стебелька. 10
— основание
скелета рук, //
— спинная створка раковины, 12
— эпнстом

Раковина состоит из углекислой извести и из роговидного органического ве-
512
щества. Структура раковины пленочных характеризуется наличием глубоко вдающихся в раковину канальцев, в которые входят выросты мантии (в раковине пластинчатожаберных моллюсков подобные канальцы отсутствуют). Непосредственно под створками лежит наружный эпителий мантийных складок, выделяющих створки.

/7
16
1
Рис. 500. Схема строения плеченогого. Руки изображены в упрощенном виде. Щупальца частично обрезаны у основания (по Кестнеру):
/ — передняя стенка тела, 2 — яичник, 3 — пе-ритонсальпый эпителий, 4 — кожный эпителий, ,5 — стебелек, в — раковина, 7—• средняя кишка,. 8 — целом, 9 — задняя складка перитоне-ального эпителия, 10 — воронка целомодукта, // — цсломодукт, 12 — желудок, 13—передняя складка перитонеального эпителия, 14 — основание руки, 15 — пищевая бороздка между основаниями щупалец, 15 — спинная мантийная складка, 17 — мантийная полость
Тело животного скрыто внутри раковины и занимает не всю полость раковины, а лишь его заднюю треть (рис. 506). На брюшной и спинной сторонах тело продолжается в мантийные складки, подстилающие раковину. Передняя стенка тела, как бы натянутая между створками, плоская или несколько вогнутая. На этой передней поверхности тела, несколько ближе к спинной створке, помещается рот, по бокам которого у большинства плеченогих отходят две руки, служащие для собирания пищи (см. рис. 504, 506). Это два спирально закрученных длинных выроста тела, усаженных на нсем протяжении двойным рядом мелких мерцательных щупалец. Ряд щупалец переходит с рук на поверхность тела и окаймляет рот с брюшной стороны. Вдоль внешней стороны рук идет ресничный желобок, по которому ко рту проходят мелкие частицы пищи. Щупальца плеченогих соответствуют таковым мшанок, а руки Brachi-opoda можно приравнять более сильно развитому лофофору мшанок.
Известную прочность и эластичность рукам многих представителей класса Brachiopoda придает особый скелет, проходящий по оси рук в виде известкового стволика. Выходя из основания
руки, скелетный стержень направляется к спинной створке и прирастает к ней (см. рис. 505). Иногда между обеими руками имеются мясистые соединительные мостики; в таких случаях и оба известковых стержня обнаруживают между собой соединения. Скелет рук бывает различного вида, хорошо сохраняется в ископаемом состоянии и поэтому служит отличным систематическим признаком. У некоторых ископаемых родов (Spirifer) руки, судя по их скелету, отличались большой длиной и были скручены в сложную спираль (рис. 507).
Со спинной стороны ротовое отверстие покрыто небольшой складкой— эпистомом, соответствующей аналогичному образованию мшанок.
Тело одето однослойным кожным эпителием, под которым находится тонкий слой соединительной ткани, а затем мерцательный псритонеаль-ный эпителий, выстилающий обширную вторичную полость тела.
513
17—763
Сплошной кожно-мускульный мешок отсутствует вследствие развития раковины, которая не позволяет телу сокращаться и изменять*форму, но сильно развиты отдельные мышечные пучки, закрывающие и открывающие раковину; они идут через полость тела от внутренней поверхности одной створки к другой (см. рис. 504). Часть пучков прикреп-

Рис. 507. Ископаемое плеченогое Spirifer. Раковина взломана так, что виден известковый спиральный скелет рук (из Яковлева)
лена к пожкс, и среди них различают приводящие, отводящие и вращающие мышцы.
Целом плеченогих представлен обширной полостью тела, содержащей кишечник, и пересекаемой мышцами раковины. Мезентерий делит целом на правую и левую половины; целом дает выросты, проникающие в толщу мантийных складок, а также узкие каналы, проходящие внутри рук. От этих каналов отходят тонкие веточки в каждое щупальце.
В полости тела плеченогих кроме продольных мезентериев имеются и неполные поперечные перегородки. Иногда их рассматривают как остатки настоящих диссепиментов, разделяющих целом на три отдела: передний — эпистомальный; средний, посылающий целомическис каналы в лофофор и щупальца, и задний — туловищный. Однако многие исследователи считают эти перегородки вторичным образованием и подчеркивают, что целом взрослых Brachiopoda не расчленен.
Полость тела содержит бесцветную жидкость с рассеянными в ней амебоидными клетками.
Пищеварительная система (см. рис. 506). Рот имеет форму поперечной щели и прикрыт эпистомом. Рот ведет в короткий эктодермальный пищевод, направляющийся вниз и переходящий в энтодермальную среднюю кишку; начальное расширение ее образует желудок. В желудок впадают с боков протоки двух крупных двухлопастных желез; они выделяют пищеварительные соки и называются печенью. У большинства плеченогих, а именно у замковых (подкл. Testicardincs), средняя кишка заканчивается слепо. Беззамкйвые (подкл. Ecardines)—более примитивные и менее измененные формы, имеют порошицу. Кишка их загибается несколько кверху и открывается наружу на правой стороне тела.
Кишечник прикреплен к стенкам тела при помощи мезентериев, или брыжеек, спинно-брюшной и двух поперечных. Как уже отмечалось, поперечные брыжейки не сплошные и имеют скорее характер нежных продырявленных пленок.
514
Нервная система слагается из очень небольшого парного надглоточного ганглия, тонких окологлоточных коннективов и лежащего под пищеводом, на брюшной стороне, более крупного подглоточного узла; в результате получается окологлоточное нервное кольцо. От надглоточного ганглия нервы идут к рукам, от подглоточного — к остальному телу.
Дифференцированные органы чувств у взрослых плеченогих отсутствуют.
Функции органов дыхания выполняют в значительной мере руки. Они обладают мерцательным эпителием и, благодаря присутствию щупалец, большой поверхностью для газообмена. Внутрь рук заходит не только продолжение полости тела, но и сосуды кровеносной системы, которая у плеченогих довольно хорошо развита.
Кровеносная система. На спинной стороне желудка помещается небольшой сократимый мешочек — сердце. От него отходит аорта, вскоре распадающаяся сначала на две, а затем на большее число артерий, главные из которых направляются в руки, складки мантии и к половым железам. Артерии ветвятся и переходят в щелевидные пространства в тканях, или кровеносные лакуны.

Органы выделения представлены каналами с широкой ресничной воронкой, открывающейся в полость тела (см. рис. 506). Их часто называют метаиефридиями, хотя по способу закладки они скорее должны рассматриваться как типичные целомодукты (их одна или две пары). Наружные отверстия лежат на верхней стороне тела, по бокам и вентрально от рта. Кроме выделительной функции отверстия служат и для выведения половых продуктов.
В
Половая система. Большинство плеченогих раздельнополы. Половых желез обыкновенно две пары и лежат они в це-ломических полостях обеих складок мантии. Половые продукты закладываются под пе-ритонсальным эпителием, по созревании поступают в целом и выводятся наружу целомо-дуктами. Оплодотворение внутреннее. Живчики проникают в полость тела самок и часть развития зародыш проходит в теле матери.
Рис. 508. Развитие плеченогого Argiopc. A — стадия отшнуровывания от кишечника целоми-ческих мешков; Б — дифференциация трех отделов тела; В — сформированная личинка с головным отделом, обрамленным ресничками и несущим глаза; туловищный отдел образовал две мантийные складки с пучками щетинок, свешивающихся книзу и частично прикрывающих стебельковый отдел; Г и Д — схема метаморфоза Brachiopoda Testicardines (по Ковалевскому) :
1 —зачаток эпистома, 2 — рот, 3 — стебелек, 4, 5««» спинная и брюшная лопасти мантии
515
Развитие. Размножение плеченогих исключительно половое. Их эмбриональное развитие впервые было выяснено А. О. Ковалевским (1873).
В результате полного, почти равномерного дробления образуется бластула, а затем инва-гинационная гаструла. В дальнейшем у многих видов бласто-пор замыкается и личинка превращается в слепо замкнутый
17*
двухслойный мешок из экто- и энтодермы. Мезодерма плеченогих закладывается иначе, чем у червей. Обычно энтодермальный кишечник дает от. себя два боковых выпячивания, которые затем отшнуровываются и ложатся по бокам кишки в виде двух целомических мешков (рис. 508, А). Эти процессы скорее напоминают развитие мезодермы у вторичноротых (с. 555).
У замковых плеченогих личинка, имевшая сначала вид овального двухслойного мешочка, несколько удлиняется и подразделяется двумя поперечными кольцевыми бороздками на 3 участка: головной, туловищный и стебельковый (рис. 508, Б). Однако существует мнение, что это деление не соответствует настоящей сегментации. Многие зоологи считают, что границы сегментов у брахиопод проходят так же, как и у других Tentaculata: предротовая лопасть, или эпистом, второй сегмент несет ротовое отверстие и лофофор (руки со щупальцами) и, наконец, общий туловищный отдел, подразделяющийся у плеченогих на собственно туловище и стебелек. Анализ сегментации целома в этом отношении мало что дает. Целомические пузырьки, отделяющиеся от кишечника, очень скоро утрачивают просвет, который заново возникает значительно позднее за счет вторичного расхождения клеток. У ряда замковых плеченогих целомические мешочки до этого успевают разделиться пополам, так что зародыш некоторое время обладает двумя парами целомов.
Головной отдел тела разрастается в виде зонтика, обрамленного ресничками, с теменной пластинкой на темени и 4 глазками. На туловищном участке образуются две свешивающиеся книзу кожные складки, спинная и брюшная, с двумя группами длинных тонких щетинок в каждой. Стебельковый отдел в виде простого сосочка. На этой стадии личинка плавает в планктоне, причем щетинки увеличивают ее поверхность и помогают ей держаться в воде. Плавающий образ жизни длится у личинок разных Brachiopoda от 10 до 30 дней. Позднее личинка оседает на дно, прикрепляется своим стебельковым отделом, а обе складки мантии заворачиваются кверху и охватывают туловище личинки (рис. 508, Д). При этом любопытном процессе внутренняя поверхность личиночной мантии становится наружной и, наоборот, наружная поверхность— внутренней. У основания головного отдела образуется впячи-вание, дающее рот и переднюю кишку, последняя соединяется с энтодер-мальным зачатком средней кишки. Далее щетинки исчезают, головной отдел редуцируется, оставляя после себя лишь небольшую складочку, эпистом, на спинном крае ртз; складками мантии выделяется раковина, стебельковый отдел вырастает в стебелек. Еще ранее этого, дорзально ото рта, появляются зачатки рук сначала в виде двух простых бугров; позднее они испытывают сложное спиральное закручивание, характерное для взрослого животного. Во время развития руки проходят стадию простых ушковидных выростов, очень напоминающих две половины ло-фофора мшанок.
Итак, развитие представителей подкл. Testicardines сопровождается глубоким метаморфозом, во время которого формируется личинка, несколько напоминающая трохофору.
У беззамковых (подкл. Ecardines) личинка выходит из яйца, уже заключенная в двустворчатую раковинку, и напоминает взрослое животное. Она ведет планктонный образ жизни, плавая благодаря работе ресничек на лофофоре, который может высовываться из раковины.
Экология. Все плеченогие — обитатели морей. Они широко распространены и встречаются на различных глубинах. Питаются Brachiopoda различными мелкими животными и органическими остатками, взвешенными в воде. Большинство совершенно неподвижно прикреплено к суб-
516
страху: Lingula (из Ecardines) имеет длинный мясистый свободный стебелек, погружающийся в мягкий грунт. Плеченогих следует считать вымирающей группой. Как уже сказано, число известных науке вымерших видов превышает 10000. Раковины Brachiopoda появляются с до-кембрийских отложений. Попадаясь в большом количестве, начиная с самых древних эпох, плеченогие имеют очень большое значение в палеонтологии, ибо служат «руководящими окаменелостями» для многих пластов. Интересно, что некоторые роды плеченогих обнаруживают в течение огромных промежутков времени крайне малую изменчивость и большую стойкость организации. Так, Lingula, встречающаяся в настоящее время очень часто у берегов Японии, сохранила все родовые признаки с силурийского периода. Среди ископаемых плеченогих особенно любопытны виды, жившие в сильно подвижной воде и прикреплявшиеся брюшной створкой. Последняя принимает вид высокого толстостенного конуса, в котором помещается животное, тогда как спинная створка низводится до степени небольшой крышечки, замыкающей вход в конус (Richthofenia).
Классификация. Класс Brachiopoda делится на два подкласса.
ПОДКЛАСС I. БЕЗЗАМКОВЫЕ (ECARDINES, ИЛИ INARTICULATA)
Наиболее древние плеченогие, известные еще из докембрийских отложений. Замковое соединение створок отсутствует. Стебелек иногда преобразуется в сократимую ногу, служащую для рытья. Кишечник изогнут и открывается наружу анальным отверстием. Выходящая в воду личинка заключена в двустворчатую раковину.
Представители: Lingula, ведущая роющий образ жизни, и др.
ПОДКЛАСС II. ЗАМКОВЫЕ (TESTICARDINES, ИЛИ ARTICLJLATA)
Филогенетически более молодые плеченогие, появляющиеся в нижнем кембрии. Створки раковины соединяются с помощью замка. Пищеварительная система слепо замкнута, анальное отверстие отсутствует. Раковина появляется только в процессе метаморфоза личинки после прикрепления последней к субстрату.
Представители: Rhynchonella, обитающая в Белом море; Magellania (см. рис. 504, 505); ископаемый Spirifer (см. рис. 507) и др.
КЛАСС III. ФОРОНИДЫ (PHORONIDEA)
Очень небольшая, состоящая всего из 18 видов группа морских животных, имеющих вытянутую, колбасообразную форму и прячущихся внутри выделяемой ими трубки, из которой высовывается только передний конец тела (рис. 509), снабженный, как и у мшанок, подковообразным лофофором с венчиком мерцательных щупалец (рис. 510), подгоняющих пищу ко рту. Последний прикрыт эпистомом. Немного позади рта лежит порошица, так что кишечник образует петлю. Дыхание совершается преимущественно через щупальца, выделение — посредством пары трубчатых органов, снабженных ресничной воронкой. Предполагается, что по своей природе это нефромиксии (см. с. 258). Через эти же органы наружу выводятся и половые продукты.
Половая система гермафродитна и состоит из яичника и семенника, лежащих у заднего конца тела. Нервная система состоит из околоротового кольца и продольного ствола, проходящего асимметрично по левой
517
стороне тела. Органы чувств представлены особым лофофоральным органом, имеющим вид язычка и расположенным позади эпистома.
Полость тела вторичная и состоит из трех отделов: переднего — эпи-стомального, среднего — кольцевидного, посылающего каналы в лофо-фор, и заднего — большого, занимающего все тело.
Кровеносная система состоит из околоротового кольца и двух продольных сосудов, переходящих один в другой на заднем конце тела.

Рис. 509. Форонис — Phoronis psammopluta в своей трубке, состоящей из зерен песка (по Сели Лоншану)
Рис. 510. Схема строения Phoronis (по Дсляжу и Эроуару):
1 — внутренний и наружный ряды щупалец лофофора, 2 — лофофораль-ный орган, выполняющий, вероятно, чувствительную функцию, 3 — выделительное отверстие, 4 — нервное кольцо, 5—анус, 6 — нервный ганглии, 7 — нефромиксии, £ — спинной кровеносный сосуд, 9 — средняя кишка, 10 — семенник, // — яичник. 12—пищевод, 13 — лофофоральный кровеносный сосуд, 14 — ротовое отверстие, 15 — эпистом
Развитие Phoronis ведет к образованию плавающей личинки актинотро-хи, похожей некоторыми признаками организации на трохофору, и сопровождается метаморфозом. Туловище «червя» образуется за счет разрастания брюшной стороны тела личинки (рис. 511).
Форониды обладают и бесполым размножением, которое осуществляется в виде поперечного деления. Образующиеся особи позднее достраивают недостающие участки тела.
У нас представители Phoronidea найдены в Черном море и в морях Дальнего Востока.
518


Рис. 511. Развитие Phoronis. A — личинка актинотроха сбоку; Б и В — стадии метаморфоза (по Маркусу):
I — теменная пластинка, 2 — рот, 3 — личиночные щупальца, 4 — анус, 5 — кишка. 6 — передняя кишка, 7 — брюшной вырост тела — будущее туловище «червя», 8 —
дефинитивные щупальца