

231 Рис. 198. А—схема организации Kinorhyncha (по Ремане); б — личинка
волосатика Gordius (по Мюльдорфу); В — личинка приапулиды Hali-
cryptus spinulosus (по Хаммарстену):
/ — рот, 2 — ротовой бугорок, 3 — головные шипы, 4 — глотка, 5 — слюнные железы, 6 — пищевод, 7 — средняя кишка, 8 — брюшные пластинки, 9 — половая железа, 10 — брюшной нервный ствол, // — половое отверстие, 12 — аналыюе отверстие, 13 — задняя кишка, 14 — дорзо-вентральные мышцы, 15 — спинные пластинки, 16 — шейные пластинки, 17 — окологлоточное нервное кольцо, 18 — крючья, 19 — хоботная железа, 20 — зачаток брюшного нервного ствола, 21 — кишка
ствие чего животное приобретает до известной степени членистый характер, состоя из 13 участков (голова, шея и 11 члеников, или зенитов туловища). Наружная расчлененность мало отражается на внутреннем строении животного, и членистость киноринх отнюдь нельзя приравнивать к настоящей сегментации, или метамерии, кольчатых червей. Голова усажена несколькими венчиками направленных назад шипов, или крючков; на остальном теле также, особенно на заднем конце, довольно многочисленны хитиновые шипы и щетинки.
Покровы состоят из кутикулы, подостланной синцитиалыюй гиподермой. Общий кожно-мускульный мешок отсутствует, и мускулатура состоит из отдельных мышечных пучков более специального назначения. Вся мускулатура поперечнополосатая. Этим класс Kinorhyncha выделяется среди всех низших червей.
Нервная система образована окологлоточным нервным кольцом и брюшным продольным нервным стволом; членистость тела сказывается на нервном стволе тем, что ганглиозные клетки собраны на нем кучками, соответственно зопитам. Органы чувств представлены осязательными волосками и парой мелких, просто устроенных глаз, лежащих над окологлоточным нервным кольцом.
Кишечник, состоит из прямой трубки со ртом на переднем и порошицей на заднем полюсах тела. Кишечник помещается в обширной первичной полости тела.
Выделительная система состоит из одной пары коротких неразветвленных протонефридиев с одним мерцательным пламенем внутри каждого.
Половая система. Киноринхи раздельнополы. Половые железы парны, причем половые протоки их открываются на заднем членике тела.
Развитие на ранних стадиях не изучено; имеются личинки, обладающие неполным числом зенитов. Рост животного сопровождается периодической линькой, причем старая кутикула сбрасывается, заменяясь новым панцирем.
Филогения. Происхождение класса Kinorhyncha не вполне ясно; сначала их считали членистоногими с атрофировавшимися конечностями, но наличие протонефридиев (у членистоногих никогда не бывает) опровергло это предположение.
Общий план строения киноринх свидетельствует о принадлежности их к первичнополостным червям. По кутикуляризакии покровов, росту с помощью линек, строению глотки они напоминают нематод, но имеют некоторое сходство с брюхоресничными (наличие протонефридиев). Однако этого недостаточно, чтобы сближать класс Kinorhyncha с какой-либо конкретной группой типа Nemathelminthes.
Некоторые исследователи отмечают, кроме того, известное сходство киноринх со скребнями и с личинками волосатиков и приапулид.
КЛАСС IV. ВОЛОСАТИКИ (NEMATOMORPHA)
Сюда принадлежит небольшая (около 225 видов) группа паразитических червей, которые отличаются от Nematoda рядом существенных признаков. Волосатики паразитируют в различных членистоногих, главным образом в насекомых.
Строение. Цилиндрическое, очень тонкое и длинное волосовидное тело (рис. 199, А), может быть от нескольких сантиметров до 1,5 м длины. Молодые паразитические стадии волосатиков беловатого цвета, взрослые черви имеют темную бурую окраску. На переднем конце тела лежит рот, на заднем — отверстие клоаки.
232
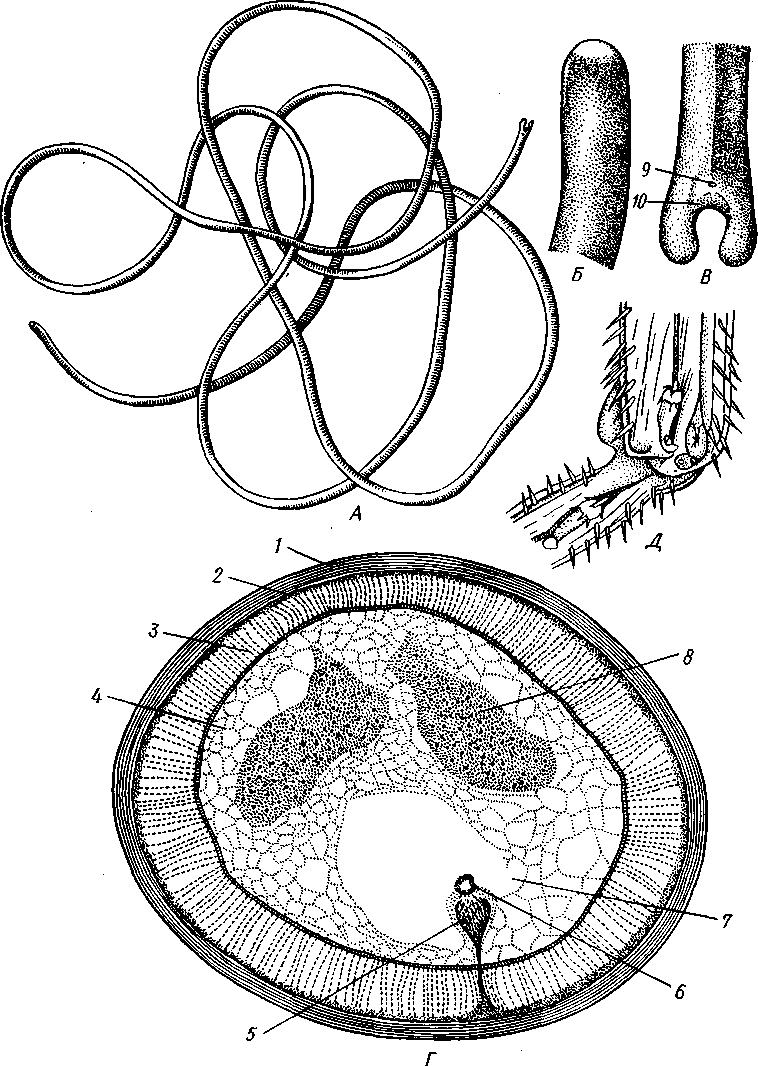
Рис. 199. Волосатик Gordius aquaticus. A—внешний вид самца в натуральную величину; Б — передний конец червя; В — задний конец самца; Г — поперечный разрез самца; Д — личинка Gordius aquaticus в ноге водной личинки поденки (Л—Г по Стрелкову,.
Д — по Мейснеру):
1 — кутикула, 2 — гиподерма. 3 — слой продольных мышечных клеток, 4 — паренхима, 5 — брюшной нервный ствол, 6 — кишечник, 7 — околокишечный синус, ft — семенной мешок, 9 — порошица, 10 --
полулунная складка кутикулы
Тело одето плотной кутикулой, выделяемой лежащим под ней однослойным эпителием, еще глубже залегает кожно-мускульный мешок, состоящий, как у нематод, лишь из продольных мышечных волокон.
Пространство между мышцами и проходящим по оси тела кишечником занято паренхимой из многогранных клеток и из прослоек соединительной ткани. Однако эти клетки занимают далеко не всю внутренность тела. Вокруг кишечника и над ним имеются большие, тянущиеся через все животное участки свободной полости тела (рис. 199, Г). Последняя представляет, как и у нематод, первичную полость тела.
Кишечник имеет вид тонкой трубки, состоящей из передней, средней и задней кишки. У многих представителей Gordiacea передняя кишка редуцируется, превращается в плотный клеточный тяж или вовсе исчезает.
Выделительная, дыхательная и кровеносная системы отсутствуют.
Нервная система состоит из залегающего в переднем конце тела нервного кольца и брюшного нервного ствола, который доходит до заднего конца тела. Нервная система лежит в самом эпителии или непосредственно под ним. Органы чувств развиты очень слабо.
Половая система. Волосатики раздельнополы. Половые железы парны и имеют вид длинных колбасовидных тел, которые начинаются слепо недалеко от переднего конца животного, а на заднем конце впадают при помощи половых протоков в заднюю кишку.
Яичники самок образуют множество следующих друг за другом боковых выпячиваний. Выводные протоки у самки состоят из яйцеводов и матки, а у самца — из двух семяпроводов. Задний отдел кишки, в который открываются половые пути, называется клоакой.
Развитие сопровождается метаморфозом с образованием личинки, резко разнящейся от взрослых волосатиков (рис. 198, Б и 199, Д).
Жизненный цикл. Волосатики паразитируют чаще всего в полости тела насекомых, особенно у некоторых прямокрылых, жужелиц и мертвоедов. Достигнув предельных размеров, волосатики выходят из хозяев наружу, пробуравливая их покровы, и в течение некоторого времени живут свободно в воде. Соответственно выход паразита из хозяина приурочен к таким'моментам, когда хозяин либо случайно попадает в воду, либо находится вблизи от воды. В воде волосатики достигают половой зрелости и копулируют, после чего самки откладывают яйца, скрепленные в длинные шнуры, на различные подводные предметы. Взрослые волосатики после этого умирают. Выходящие из яиц личинки могут некоторое время жить в воде или в сырой земле. В это время они отыскивают промежуточного хозяина (обычно водные личинки насекомых) и внедряются в него через кожу при помощи своего сверлящего хоботка. Наземные насекомые заражаются, поедая инвазированных водных личинок. В человеке и домашних животных волосатики никогда не паразитируют. Народное поверье о том, что проглоченные во время питья воды волосатики могут стать причиной заболеваний, не имеет никакого основания.
Филогения волосатиков слабо выяснена. Частью признаков: общий вид, кутикула, строение мускулатуры, некоторое сходство в форме половых желез — класс Nematomorpha напоминает нематод. Родство их с типом Nemathelminthes оправдывается также сходством личинок волосатиков и представителей класса Kinorhyncha.
234
КЛАСС V. КОЛОВРАТКИ (ROTATORIA)
Очень мелкие водные животные, большей частью не. превышающие своими размерами крупных инфузорий. Коловратки — преимущественно обитатели пресных вод, хотя имеются и довольно многочисленные морские представители кл. Rotatoria. Громадное большинство коловраток свободноподвижные, немногие ведут прикрепленный образ жизни. Известно свыше 1500 видов.
Строение и физиология. Размеры коловраток не превышают 1—2 мм, но есть среди них формы и гораздо мельче. Так, к коловраткам относится самый мелкий представитель многоклеточных Ascomorpha minima, достигающий едва 0,04 мм длины. Форма тела сильно варьирует. В редких случаях (Trochosphaera) тело шаровидно (см. рис. 201, Д), тогда как у громадного большинства оно продолговато и разделяется на три участка: передний головной с мерцательным аппаратом, туловищный, содержащий все внутренности, и задний, ножной (рис. 200), Нога может и отсутствовать.

Головной отдел мало обособлен от туловищного, а внутренне и вовсе не отделен от него. Передний конец головного отдела в некоторых случаях имеет вид диска, усаженного по краю венчиком крупных ресниц (рис. 200). Позади венчика на брюшной стороне головы лежит рот, позади рта — второй венчик ресниц, более мелких. Совокупность обоих венчиков образует характерный для Rotatoria ко- 2-ловращательный аппарат. Реснички его находятся в постоянном мерцании, чем и обусловливают движение животного в воде, его плавание. Одновременно с этим движением ресничек в воде вызывается водоворот, привлекающий мелкие пищевые частицы ко рту коловратки.
Форма коловращательного аппарата изменчива. Так, края предротового диска могут образовать различные выросты, а за краями следует и венчик ресниц; то диск образует четыре краевые лопасти (Floscula-ria; рис. 201, А), то он вытягивается в пучок длинных щупалец (Stephanoceros) и т. д.
Рис. 200. Анатомия коловратки, схематизиро-вид со спинной стороны; Б — вид сбоку (по Ремане):
/ — головные чувствительные щупальца, 2 — коловра-щательный аппарат, 3 — рот, 4 — глотка с жеватель-
202). У ЭТИХ КОЛОВраТОК ВМеСТО 7H±"»SS£S,a£e! жГлеСзТТ1жЖе!?удоЬк, ^-я^нТ
ДВУХ веНЧИКОВ МОЩНЫХ реСНИЦ ГойЛ^рТ^Г^оа^^ГГверТтГ^^-цем^:
ИЗ ОрЮШНОИ СТОрОНе Передне- ные железы, 15 — ножной ганглий, 76 — пальцы ноги,
ГП пппшгя трля имрртро r-nnnin /7 —клоака, /« — спинные чувствительные щупаль-
ГО ПОЛЮСа Тела ИМееТСЯ СПЛОШ- ЦЭр 19 _ надглоточный ганглий
Однако за исходную форму коловращательного аппарата всех Rotatoria в настоящее время принимают слабо диф- ван0' ференцированный аппарат семейства Notommatidae (рис.
235
21
36
Д

Рис. 201. Коловратки. А—сидячая коловратка Floscularia ringens, строящая домик из комочков своих экскрементов; Б — сидячая коловратка Stephanoceros fimbriatus (по Везенберг — Лунду); В — Вгас1г1опи$,самка; Г — Brachionus, самец (по Везен-берг — Лунду); Д — планктонная коловратка Trochosphaera solsticialis (по Русселг): I—лопасти коловращательного аппарата, 2 — боковое щупальце, 3 — головное щупальце, 4, 15 — глотка с жевательным аппаратом, 6 — желудок, 6 — задняя кишка. 7 — мочевой пузырь, 8 — студенистый домик, 9 — щетинки коловращательного аппарата, 10—чувствительные щетинки, // — реснички коловращательного аппарата, 12 — спинное щупальце, 13 — мозговой ганглий, 14 — ретрак-торы переднего конца тела, 16 — пищеварительные-железы, /7 —панцирь, 18 — канал мротонефри-дия, 19 — концевая клетка протонефридия с мерцательным пламенем, 20 — боковой нерв и чувствительное щупальце, 11 — яичник, 22— кишка, 23 — заднепроходное отверстие, 24 — ножные железы, 25 — их протоки, 26 — пальцы ноги, 27 — мочевой пузырь, 28 — яйцо, 29 — желудок, 30 — пищевод, 31 — глазок, 32 — семенник. 33 — экваториальный нерв, 34 — глазок, 35 — предротовой венчик ресниц, 36 — кишечник, 37 — клоака. 38 — протонефридий, 39 — рот
ная мерцательная площадка из мелких ресничек, окружающая рот. Такой мерцательный аппарат примитивных коловраток напоминает ресничный покров многих Gastrotricha.
При помощи особых мышц-ретракторов головной отдел может втягиваться внутрь туловищного и затем снова выворачиваться.

Рис. 202. Примитивный тип устройства мерцательного аппарата коловраток. А — вид с брюшной стороны; Б — вид сбоку (по Бошану): / — передний край головы, 2 — боковые выступы мерцательного поля, 3 — рот, 4 — брюшное мерцательное поле, 5 — глотка
Нога — мускулистый вырост тела, одетый членистой оболочкой и снабженный на конце двумя щупальцевидными подвижными отростками, пальцами. В основании пальцев имеются две небольшие цементные железы, выделяющие особое клейкое вещество. При помощи его многие коловратки могут временно, а другие и постоянно прикрепляться к отдельным подводным предметам. В ноге хорошо развиты кольцевые и продольные мышцы. Нога позволяет коловраткам ползать, вытягиваясь по направлению движения и временно прикрепляясь передним концом к субстрату.
Стенка тела образована синцитиальным слоем гиподермы. Распространенное ранее мнение о сильном развитии кутикулы у коловраток оказалось неверным. Последние исследования, проведенные с помощью электронного микроскопа, показали, что для Rotatoria характерно наличие в гиподерме плотного волокнистого слоя под наружной цитоплазматической мембраной. Именно этому слою обязаны коловратки сохранением формы тела и из него сформированы уплотненные покровы панцирных видов. Настоящая кутикула нетипична для коловраток и известна лишь у ограниченного числа форм.
Кожно-мускульный мешок у коловраток отсутствует. В теле имеются отдельные, большей частью поперечнополосатые мышечные волокна, служащие для втягивания головного отдела, для сокращения туловища и т. д. Непосредственно под наружным эпителием лежит первичная полость тела.
Пищеварительная система коловраток состоит из трех отделов. Рот лежит неподалеку от переднего конца, на брюшной стороне между венчиками ресниц коловращательного аппарата. Он ведет в ротовую полость, а та, расширяясь, — в мускулистую глотку с характерным для коловраток жевательным аппаратом, состоящим из заложенных в стенках глотки двух пар твердых зазубренных хитиновых пластинок. Это пара молоточков и пара наковален. Они трутся друг о друга, подобно паре микроскопических жерновов, и содействуют перетиранию пищи, которая состоит у коловраток главным образом из жгутиконосцев, одноклеточных водорослей и т. п.
У хищных коловраток вооружение глотки способно выдвигаться через ротовое отверстие и служит для ловли добычи.
Глотка переходит в узкий пищевод, а за ним следует мешковидный
237
энтодермальный желудок из крупных многоугольных клеток. В место соединения пищевода с желудком открывается пара желез, функция которых точно не выяснена. Желудок ведет в более узкую заднюю кишку, а последняя открывается в клоаку.
Срок пребывания пищи в кишечнике коловраток очень незначителен: от времени захвата до дефекации частицы проходит от 2 до 20 мин.
Нервная система коловраток сравнительно проста, так как состоит из одного надглоточного ганглия и из отходящих от него во все стороны и кпереди, т. е. к диску, и кзади, т. е. к туловищу, нервов. Значительнее других два нерва, идущих от мозга назад по бокам кишечника через все тело до ноги. Органами чувств коловраткам служат прежде всего щупальца. Их чаще всего три: два брюшных и одно непарное на спине. Щупальца имеют вид маленьких конических бугорков с пучком осязательных волосков на вершине.
Большинство коловраток имеет пару или всего один глаз. Глаза лежат на самом надглоточном ганглии или же непосредственно над ним. Они устроены крайне просто: состоят из маленького лучепреломляюще-го хрусталика, лежащего в бокале из красного пигмента. По-видимому, это глаза инвертированного типа. Когда глаз один, то он представляет результат слияния пары глазков, обращенных друг к другу выпуклыми сторонами пигментных бокалов, почему пигмент в таком глазу дает фигуру X.
Кровеносная и дыхательная системы у коловраток отсутствуют.
Выделительные органы протонефридиального типа. По сторонам от кишечника в полости тела залегают два слабо ветвящихся канальца, открывающихся сзади в клоаку. Концы веточек каналов замкнуты слепо и внутри них бьет мерцательное пламя. Число веточек варьирует от 4 до 50.
Половая система. Коловратки раздельнополы и отличаются половым диморфизмом. Самки встречаются гораздо чаще самцов, и все предыдущее описание относится именно к ним. Они снабжены цельным или двухлопастным яичником, который лежит в задней части туловища, под кишечником. Обыкновенно яичник разделен на две части: производящую яйцевые клетки, или собственно яичник, и продуцирующую клетки, идущие для питания яиц, т. е. желточник. Яичник впадает при помощи короткого яйцевода в клоаку.
Самцы (см. рис. 201, Г) значительно меньше самок и часто заслуживают названия карликовых. Кишечник самцов редуцирован, выделительная система развита слабо; тотчас после оплодотворения самцы умирают. Мужская половая система состоит из единственного семенника с отходящим от него семяпроводом, впадающим в клоаку, причем конец его пронизывает мускулистый, способный к выворачиванию совокупительный орган, или cirrus. Сильная редукция многих органов у самцов объясняется кратковременностью их жизни, которая заканчивается сразу после оплодотворения самки.
Жизненный цикл и развитие. Жизненный цикл коловраток представляет чередование настоящих половых поколений (включают самцов и самок) с партеногенетическими, т. е. имеет характер гетерогонии. Обыкновенно весной из яиц, пролежавших зиму, выходят партеногенетические самки, дающие начало опять-таки партеногенетическим самкам, и затем может следовать еще ряд таких же партеногенетических поколений (рис. 203). Далее наступает половой период, когда одно из поколений партеногенетических самок начинает откладывать яйца по объему в 2— 3 раза мельче яиц, дававших до сих пор самок. Меньшим количеством материала в яйце объясняются карликовые размеры самцов и быстрота
238

Количество жизненных циклов в течение одного года варьирует у различных коловраток: соответственно различают моноциклические, дициклические и полициклические виды. Впрочем, нередко один и тот же вид, но в разных странах или даже в разных водоемах одной и той же местности обнаруживает различное годовое число циклов. Это показыва-
Рис. 203. Годичный цикл Апигаеа cochlearis (no Кречмеру):
/ — зимнее яйцо, 2—6 — последовательные в течение лета поколения партеногенетических самок с летними яйцами; самки последнего поколения (в) откладывают мужские яйца, из которых развиваются самцы, 7 — самец. Самцы копулируют с самками последнего поколения, которые откладывают оплодотворенные зимние яйца (/)
ет, что цикличность коловра-
ток в значительной мере
зависит от внешних усло-
вий, меняющихся в разных
водоемах и местностях. От
внешних же факторов, по-
видимому, зависит и появле-
ние полового поколения са-
мок, производящих самцов. В результате значительного количества эк-
спериментальных работ выясняется, что появление самок полового поко-
ления обусловливается и ускоряется различными факторами: условиями
питания (пониженное питание, голодание ведет к продукции самцов),
химическим составом среды.
Цикличность коловраток усложняется наличием у многих видов сезонных изменений, не связанных с характером размножения. Так, например, у ряда следующих друг за другом в течение года поколений длинношипная зимняя форма Апигаеа cochlearis может постепенно превращаться в формы со слабо развитыми шипами и, наконец, совершенно лишается большого заднего шипа, переходя в поколение, которое называется A. cochlearis tecta и раньше считалось за самостоятельный вид. К зиме она вновь возвращается к исходной форме. Такие периодические изменения в строении отдельных поколений коловраток называются цикломорфозом (рис. 203).
Развитие яиц коловраток идет быстро; у партеногенетических яиц, не имеющих периода покоя, развитие длится 3 — 4 дня. Развитие прямое, без метаморфоза; в общем дробление очень рано становится двусторон-не-симметричным. Другая характерная черта развития — формирование органов из небольшого числа клеток, после чего размножение клеток в
239
них на всю жизнь прекращается. Результатом этого является постоянство клеточного состава Rotatoria, подобное отмеченному у представителей кл. Nematoda. Для многих коловраток точно доказано, что тело их состоит из небольшого, вполне определенного и постоянного числа клеток. Так, например, у Epiphanes senta покров состоит из 301 клетки, глотка — из 165-, средняя кишка — из 76, половой аппарат — из 19, мускулатура— из 122, нервная система—из 247, выделительная система — из 24, а все тело животного — из 959 определенным образом расположенных клеток. У самцов в связи с редукцией кишечника клеток еще меньше. Постоянство клеточного состава связано с тем, что после эмбрионального периода клетки коловраток перестают делиться. С последним обстоятельством, в свою очередь, связано отсутствие у коловраток способности к регенерации. Утраченные или удаленные экспериментально участки тела не восстанавливаются. Та же особенность наблюдается и у других животных, обладающих постоянством клеточного состава, например у нематод.
Экология. Большинство коловраток живет в пресных водах, причем они делятся на донные и планктонные формы. Донные коловратки большей частью ползают или плавают у дна, но среди них имеется и целый ряд сидящих видов, последние прикрепляются выделением цементных желез ноги к субстрату, причем нога изменяется в своего рода стебелек. Большинство сидячих коловраток выделяет вокруг себя защитную трубку из прозрачного студенистого вещества или склеенных вместе в правильные комочки испражнений коловратки, которыми трубки обложены как микроскопическими кирпичами. У типичных планктонных коловраток на теле вырастают различного рода придатки, увеличивающие поверхность животного. У некоторых это длинные неподвижные шипы панциря (Notholca], у других — подвижно сочлененные с телом гребные щетинки (Polyarthra) или это особые, снабженные мышцами и усаженные щетинками выросты самого тела (Pedalion). Пресноводные коловратки играют немаловажную роль как пища различных, более крупных пресноводных организмов. Морских коловраток значительно меньше.
Небольшое количество коловраток приспособилось к жизни на суше, главным образом во влажном мху, среди лишайников и т. п. Некоторые из них (Callldina) охотно поселяются в особых мешочках на слоевище печеночных мхов, в которых долго удерживается вода.
Наземные, а также некоторые водные коловратки способны выдерживать полное высыхание, не погибая. При этом они впадают в анабиотическое состояние (мнимая смерть), но после осторожного смачивания оживают. Оживание коловраток было впервые открыто Левенгуком в 1701 г. и с тех пор многократно исследовалось. При применении ряда предосторожностей смоченные водой коловратки оживают после многолетнего высушивания. Мало того, высушенные коловратки способны в течение 4 ч выдерживать понижение температуры до —270°С (в жидком гелии) и в течение 5 мин повышение температуры до +100°С.
Филогения коловраток. О происхождении коловраток существовали различные точки зрения. Долгое время их считали неотеническими личинками каких-то кольчатых червей на основании сходства Rotatoria с личинкой Polychaeta трохофорой (см. рис. 225).
В последние годы установилось мнение, что сходство коловраток с трохофорой следует считать результатом конвергенции, или совпадения, связанного с одинаковым образом жизни, путь же эволюции группы Rotatoria идет от прямокишечных турбеллярий. Защитники этого мнения считают наиболее примитивными не плавающих, а ползающих коловраток и исходным типом не хорошо дифференцированные венчики, а
240
сплошную брюшную площадку ресниц, окружающую рот. Эта площадка— остаток общего мерцательного покрова Rhabdocoela. О близости к турбелляриям свидетельствует примитивное строение протонефридиев коловраток и брюшное положение рта. Из первичнополостных червей к кл. Rotatoria ближе всего стоит кл. Gastrotricha. Их сближает отсутствие кожко-мускульного мешка, наличие протонефридиев, некоторое сходство в строении мерцательного аппарата.