

4. Теории происхождения многоклеточности.
Классические колониальные теории возникновения многоклеточности основаны на агрегации одноклеточных в метазоон. Они опираются на онтогенез Metazoa и строение колонии Volvox. Наблюдается поразительное сходство между развитием и строением высших Вольвоксовых и бластулы низших Metazoa (губки, гидроиды, иглокожие). Дробление бластулы можно соотносить с палинтомией у вольвокса. Но Вольвоксовые относятся к фитомонадам и непосредственной родней Metazoa не могут быть, поэтому их сходство указывает только на закономерность процессов возникновения многоклеточности на флагеллятном уровне организации.
Первую колониальную гипотезу – гипотезу «гастреи» – разработал Э. Геккель (1874). Он считал, что протозойным предком Metazoa была «бластея» – шаровидная колония жгутиковых, похожая на стадию бластулы в онтогенезе многоклеточных. От нее путем инвагинации (впячивания) мог возникнуть первый двуслойный многоклеточный организм – «гастрея» (рис. 11, I). Ее наружный слой (эктодерма) выполнял двигательную функцию, а внутренний (энтодерма) – пищеварительную. Непосредственно от гастреи произошли кишечнополостные. О. Бючли в 1884 г., развивая теорию «гастреи», предложил ее новый вариант – гипотезу «плакулы » (рис. 11, II). Он считал гипотетическим колониальным предком пластинчатую колонию типа современных Gonium. Путем разделения такой пластинки на два слоя возникла «плакула», а из нее, путем чашевидного изгибания – гастрея. Бючли основывался на строении трихоплакса (Trichoplax), которое сходно с плоскими колониями жгутиконосцев. Он предполагал, что подобные трихоплаксу животные могли быть промежуточными между «плакулой» и «гастреей». Из нижнего слоя клеток примитивных плакулообразных организмов возникла энтодерма гастреи, а из верхнего – эктодерма.
И .И.
Мечников в 1882 г.
опубликовал гипотезу «фагоцителлы ».
Он открыл фагоцитоза этот способ
переваривания пищи более примитивным,
чем полостное пищеварение. Поэтому
первые многоклеточные были примитивнее
«гастреи » по организации и не имели
пищеварительной полости и полостного
пищеварения. Эта теория находит
подтверждение в ходе онтогенеза
большинства губок и кишечнополостных.
У них образование двуслойной фазы
развития происходит не путем инвагинации,
а путем иммиграции части клеток наружного
слоя в полость зародыша (бластоцель).
Такая личинка с паренхимными клетками
внутри была названа паренхимулой, и
Мечников рассматривал ее как живую
модель гипотетической фагоцителлы
(рис. 11, III). Она могла возникнуть из
шаровидных колоний жгутиконосцев путем
иммиграции клеток внутрь колонии. При
этом наружные клетки со жгутиками
выполняли функцию движения (кинобласт),
а внутренние утрачивали жгутики,
становились амебоидными и выполняли
функцию фагоцитоза (фагоцитобласт).
.И.
Мечников в 1882 г.
опубликовал гипотезу «фагоцителлы ».
Он открыл фагоцитоза этот способ
переваривания пищи более примитивным,
чем полостное пищеварение. Поэтому
первые многоклеточные были примитивнее
«гастреи » по организации и не имели
пищеварительной полости и полостного
пищеварения. Эта теория находит
подтверждение в ходе онтогенеза
большинства губок и кишечнополостных.
У них образование двуслойной фазы
развития происходит не путем инвагинации,
а путем иммиграции части клеток наружного
слоя в полость зародыша (бластоцель).
Такая личинка с паренхимными клетками
внутри была названа паренхимулой, и
Мечников рассматривал ее как живую
модель гипотетической фагоцителлы
(рис. 11, III). Она могла возникнуть из
шаровидных колоний жгутиконосцев путем
иммиграции клеток внутрь колонии. При
этом наружные клетки со жгутиками
выполняли функцию движения (кинобласт),
а внутренние утрачивали жгутики,
становились амебоидными и выполняли
функцию фагоцитоза (фагоцитобласт).
Гипотеза фагоцителлы как наиболее обоснованная завоевала широкое признание и развивалась другими учеными. Так, А.А. Захваткин (1949) предположил, что предковые колониальные простейшие имели не голофитный, а голозойный способ питания и гаметическую редукцию хромосом в онтогенезе. С другой поправкой Захваткина трудно согласиться. Он считал, что фагоцителла отражает строение не взрослого предка многоклеточных, а расселительной личинки синзооспоры. А взрослая фаза представляла сидячую форму колониального типа, похожую на губок. Но трудно допустить, что такие прикрепленные формы могли дать начало всем Metazoa.
А
1
2
3
4
5
6
7
8
9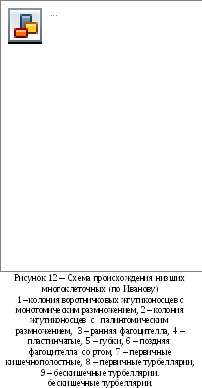 ереходе
к ползанию по субстрату возникли
Пластинчатые (рис. 12, 4), а при переходе
к прикрепленному образу жизни Губки, и
при этом произошло извращение зародышевых
листков (рис. 12, 5). При плавании фагоцителлы
пища концентрировалась на заднем полюсе,
что привело к формированию на нем
ротового отверстия, и появилась поздняя
фагоцителла (рис. 12, 6). При опускании на
дно и переходе к ползающему образу жизни
она дала начало примитивным ресничным
червям с ротовым отверстием на заднем
конце тела, которое в ходе эволюции
постепенно смещалось вперед(рис.
12, 8, 9). У плавающих форм
происходило расхождение клеток паренхимы
к периферии, что привело к появлению
гастральной полости и возникновению
типа Кишечно-полостные (рис. 12, 7).
Часть кишечнополостных осталась
планктонными формами (медузы), а часть
перешла к прикрепленному образу жизни
и дала начало гидрозоям и коралловым
полипам. Сравнительный анализ колониальных
гипотез приведен в таблице (табл. 1).
ереходе
к ползанию по субстрату возникли
Пластинчатые (рис. 12, 4), а при переходе
к прикрепленному образу жизни Губки, и
при этом произошло извращение зародышевых
листков (рис. 12, 5). При плавании фагоцителлы
пища концентрировалась на заднем полюсе,
что привело к формированию на нем
ротового отверстия, и появилась поздняя
фагоцителла (рис. 12, 6). При опускании на
дно и переходе к ползающему образу жизни
она дала начало примитивным ресничным
червям с ротовым отверстием на заднем
конце тела, которое в ходе эволюции
постепенно смещалось вперед(рис.
12, 8, 9). У плавающих форм
происходило расхождение клеток паренхимы
к периферии, что привело к появлению
гастральной полости и возникновению
типа Кишечно-полостные (рис. 12, 7).
Часть кишечнополостных осталась
планктонными формами (медузы), а часть
перешла к прикрепленному образу жизни
и дала начало гидрозоям и коралловым
полипам. Сравнительный анализ колониальных
гипотез приведен в таблице (табл. 1).
Таблица 1. Колониальные гипотезы происхождения Metazoa.
|
Гипотеза |
Колониальный предок из Protozoa |
Процесс преобра- зования |
Предок мно- гоклеточных Metazoa |
Живая модель предка |
|
«Гастреи» (Геккель, 1874) |
Бластея (одно- слойная шаро- видная) |
Инваги- нация |
Гастрея (дву- слойная ша- ровидная со ртом) |
Гаструла и личинка кишечнополостных |
|
«Плакулы» (Бючли, 1884) |
Плоская одно- слойная коло- ния |
Расслоение колонии и изгибание |
Гастрея (дву- слойная со ртом) |
Трихоплакс, гаструла |
|
«Фагоцителлы» (Мечников, 1882) |
Бластея (одно- слойная шаро- видная) |
Иммигра- ция |
Фагоцителла (из двух сло- ев без рта) |
Личинка гу- бок – па- ренхимула |
|
«Фагоцител- лы» (Иванов, 1967) |
Колония во-ротничковых жгутиковых (без полости) |
Иммигра- ция |
Фагоцителла (из двух сло- ев без рта) |
Паренхиму- ла, трихо- плакс |
Кроме колониальных, существуют также и полиэнергидные гипотезы происхождения многоклеточных, которые исходят из того, что предками Metazoa были полиэнергидные простейшие. Образование многоклеточности происходило путем неполного обособления составляющих энергид и превращения их в тканевые клетки. Впервые эта идея была предложена Иерингом (1877), а позже активно защищалась Стейнбоком (1954), Иованом Хаджи (1944, 1963) и некоторыми другими учеными. По их мнению, предками Metazoa были многоядерные инфузории или другие полиэнергидные простейшие, схожие, например, с опалинами. Инфузории имеют сложное строение: клеточный рот, глотку, сократительные вакуоли с приводящими каналами, трихоцисты, мионемы и т.д. Эти дифференцированные органеллы Хаджи считал гомологичными органам многоклеточных, выполняющих сходные функции. Так, кожные покровы Metazoa произошли из эктоплазмы, кишечник – из эндоплазмы, мышцы – из мионем, органы выделения – из пульсирующих вакуолей, гонады – из микронуклеуса. Акт спаривания выводится из конъюгации инфузорий, а процесс оплодотворения яйца – из слияния половых ядер конъюгантов. Процесс образования многоклеточности происходил путем целлюляризации, т.е. обособления цитоплазмы вокруг ядер с появлением границ между клетками. За примитивнейших Metazoa, непосредственно выводимых из инфузориеобразного прототипа, Steinbock и Хаджи принимают Turbellaria Acoela, которым приписывают синцитиальную структуру. Но утверждение о полном отсутствии клеточной структуры у Acoela ошибочно и онтогенез Acoela менее примитивен, чем онтогенез губок иди гидроидов, и служить прототипом для них бескишечные турбеллярии не могут. Таким образом, авторы полиэнергидных гипотез допускают определенную механистичность в объяснении происхождения от Protozoa довольно высокоорганизованных трехслойных животных – червей. В этих гипотезах не учитываются постулаты клеточной теории, теории зародышевых листков, биогенетический закон, гомологичность структур, особенности жизненных циклов предков и потомков.
Считают, что в настоящее время наиболее серьезно аргументирована гипотеза фагоцителлы И.И. Мечникова, доработанная А.В. Ивановым.