

45. Структурно-функциональная схема условного рефлекса и механизм образования временной связи (по и.П. Павлову). ???????
Всякий УР может быть выработан на основе БУР. Поэтому обязательным компонентом УР является дуга безусловного рефлекса. Как Вам уже известно: любой БУР, даже если он осуществляется без вмешательства коры головного мозга, имеет свое корковое представительство (КПБУР). Процесс выработки УР заключается в том, что относительно индифферентный раздражитель (например, звук) при определенных условиях приобретает сигнальное значение. Т.е. сигнализирует о том, что вскоре появится безусловный раздражитель, например, пища. В нашем примере звук является условным раздражителем. Его воспринимает определенная афферентная система. При этом каждая афферентная система, также как и БУР имеет свое корковое представительство – корковое представительство афферентной системы условного раздражителя (КПАСУР). Это образование также генетически закреплено. При соблюдении определенных условий между этими корковыми представительствами возникает временная связь (ВС). Возбуждение доходит до КПАСУР, возникшее там возбуждение передается по временной связи до КПБУР и оттуда на эффектор (на нашей схеме это слюнная железа).
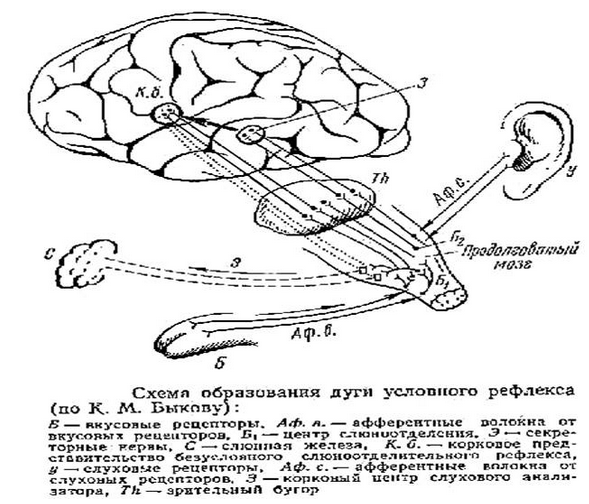
Таким образом, условный рефлекс – это приспособительная деятельность, осуществляемая высшими отделами ЦНС путем образования временных свзей между сигнальным раздражением и сигнализируемой реакцией. На базе УР формируется поведение.
Правила выработки условного рефлекса по Павлову: 1)Для выработки условного рефлекса необходимо наличие 2-ух раздражителей: условного (свет) и безусловного (мясо). Например: слюноотделение у собаки на свет при подаче мяса. 2)Нужно сначала давать условный раздражитель, только потом безусловное подкрепление. 3)Условный раздражительнее должен быть чрезвычайным. 4)Условный раздражитель не должен быть биологически более значим, чем безусловное подкрепление. 5)При выработки безусловного рефлекса не должно быть посторонних доминантов (переполненный мочевой пузырь). 6)При выработке условного рефлекса должна быть выражена мотивация. При выработке слюноотделительного рефлекса животное должно быть голодным, у сытого – этот рефлекс не вырабатывается.
46. Правила выработки условных рефлексов. 1. Неоднократное сочетание условного раздражителя и подкрепления 2. Условный раздражитель должен быть достаточно сильным, чтобы вызвать ориентировочную реакцию 3. По биологической силе безусловный раздражитель должен быть сильнее 4. Условный раздражитель должен опережать по времени 5. Не должно быть посторонних раздражителей 6. Активное состояние ЦНС без патологий
47. Торможение условнорефлекторной деятельности. Виды торможения и их характеристика. Торможение – активный процесс подавления возбуждения Делится на БЕЗУСЛОВНОЕ, УСЛОВНОЕ и ЗАПРЕДЕЛЬНОЕ Условное торможение (внутреннее, приобретенное) – если условный раздражитель перестает подкрепляться безусловным. Это торможение требует для выработки времени, к нему относят: 1. Угасательное (перестает подкрепляться безусловным: забываний неповторяющегося материала) 2. Дифференцировочное (вырабатывается на раздражители, близкие по характеру к условному) (Звук метронома с частотой 120 Гц – слюна, частота – 60 Гц – слюна, но позже дифференцировка звуков, слюна не выделяется) 3. Условный тормоз (разновидность дифференцировки, возникает, если условный раздражитель подкрепляется безусловным, а комбинация из условного и индифферентного (дополнительный) не подкрепляется, например, свет + звонок – реакция на свет, но позже нет реакции, звонок – тормоз), муха в супе 4. Запаздывающее (возникает при отставании подкрепления на 1-3 минуты относительно начала действия условного сигнала, постепенно появление условной реакции сдвигается к моменту подкрепления) пример: фальстарт Безусловное торможение – торможение, которое проявляется в ослаблении и прекращении протекающего в данный момент условного рефлекса при действии какого-либо постороннего раздражителя: 1. Постоянный тормоз – дополнительный раздражитель, который при повторении не теряет своего тормозящего свойства 2. Гаснущий тормоз –посторонний сигнал, который с повторением его действия теряет свое тормозящее свойство: торможение у собаки на команду сидеть Запредельное торможение (охранительное) – возникает при действии чрезвычайно сильного условного раздражителя
48. Учение И.П. Павлова о высшей нервной деятельности. Типы высшей нервной деятельности, критерии их оценки (сила, уравновешенность, подвижность). Понятие о первой и второй сигнальных системах. Изучение высшей нервной деятельности, проведенное Павловым, с неоспоримой убедительностью доказало идею Сеченова, что деятельность головного мозга в самых сложных своих проявлениях осуществляется по рефлекторному принципу. Основной акцент в исследованиях Павлова сделан, однако, не на изучении закономерностей поведения, а на изучении закономерностей механизма рефлекторной деятельности, лежащей в основе поведения. Павлов выделил четыре типа ВНД: Безудержный. (Холерик). Сильный, уравновешенный. Человек сильно раздражается и медленно успокаивается Инертный. (Флегматик). Сильный, уравновешенный, инертный. Процессы возбуждения и торможения сильные, с плохой подвижностью. Человек с трудом переключается с одного вида деятельности на другой Живой. (Сангвиник). Сильный, уравновешенный, подвижный. Сильные процессы торможения и возбуждения с хорошей подвижностью. Человек легко адаптируется к заданным условиям окружающей среды Слабый. (Меланхолик). Слабый. Возбуждение и торможение слабые. Адаптивная реакция организма на условия внешней среды очень плохая Показатели типологических различий возбудительного и тормозного процессов: 1. Сила – число сочетаний условного раздражителя и подкрепления для выработки УР 2. Уравновешенность – степень слабости торможения относительно возбуждения 3. Подвижность – способность реагировать на положительный условный раздражитель как на отрицательный и наоборот Взаимоотношения организма со средой осуществляются на основе сигналов, поступающих в нервную систему в результате непосредственного воздействия предметов и явлений внешнего мира на рецепторы. Этот тип сигнализации И. П. Павлов назвал первой сигнальной системой. В животном мире первая сигнальная система является единственным каналом информации организма о состоянии среды. Различные предметы внешнего мира, их физические и химические свойства (звук, цвет, форма, химический состав и др.) приобретают значение условных сигналов, оповещают организм о наступающих вслед за ними явлениях, вызывая тем самым приспособительные реакции. Например, дремлющее травоядное животное при звуке шагов или запахе хищника спасается бегством, так как эти раздражители сигнализируют об опасности. При воспитании человека развивается вторая сигнальная система, характерная только для человека. Это переводит высшую нервную деятельность человека на более высокую ступень. Она приобретает новые качества, обусловливающие расширение возможностей общения с внешним миром и многогранность его проявлений. И. П. Павлов назвал вторую сигнальную систему «чрезвычайной прибавкой» к механизмам высшей нервной деятельности человека. Вторая сигнальная система — это речь, слово, видимое, слышимое, произносимое мысленно. Это высшая система сигнализации окружающего мира. Она состоит в словесном обозначении всех его сигналов и в речевом общении. Вторая сигнальная система развилась у человека под влиянием социальной среды в процессе труда. Большую роль в этом сыграли кинестезические раздражения мозга, возникающие в результате трудовых процессов. Слово для человека служит таким же физиологическим раздражителем, как предметы и явления окружающего мира.
49. Высшие психические функции. Память: классификация, физиологические механизмы кратковременной и долговременной памяти Высшие психические функции (ВПФ) – это специфические психические функции человека. К ним относят: память, внимание, мышление, восприятие, воображение и речь. Память – позновательная функция, позволяющая накапливать, хранить и воспроизводить воспринимаемую информацию Виды памяти по продолжительности: 1. Мгновенная 2. Долгосрочная 3. Краткосрочная Виды памяти по форме проявления: 1. Образная (запоминание образов) 2. Эмоциональная (воспроизведение эмоционального состояния) 3. Словесно-логическая (запоминание информации в словесной форме) Психологическая классификация: 1. Процедурная (память на действия) – память тела 2. Декларативная – память на названия (память души) По Анализаторам: зрительная, вкусовая, мышечная, обонятельная, тактильная, эмоциональная Теория кратковременной памяти – образование кругов реверберации в определенных структурах мозга (150 мс – 15-20 минут) Теория долговременной памяти – биохимические изменения в пределах одного нейрона, увеличение синапса в размерах, посттетаническая потенциация (облегчение синаптической передачи за счет сенситизации НМДА-рецепторов, деполяризация олигодендроцитов) Консолидация – переход из кратковременной памяти в долговременную. Гиппокамп и височная доля КБП отвечают за память

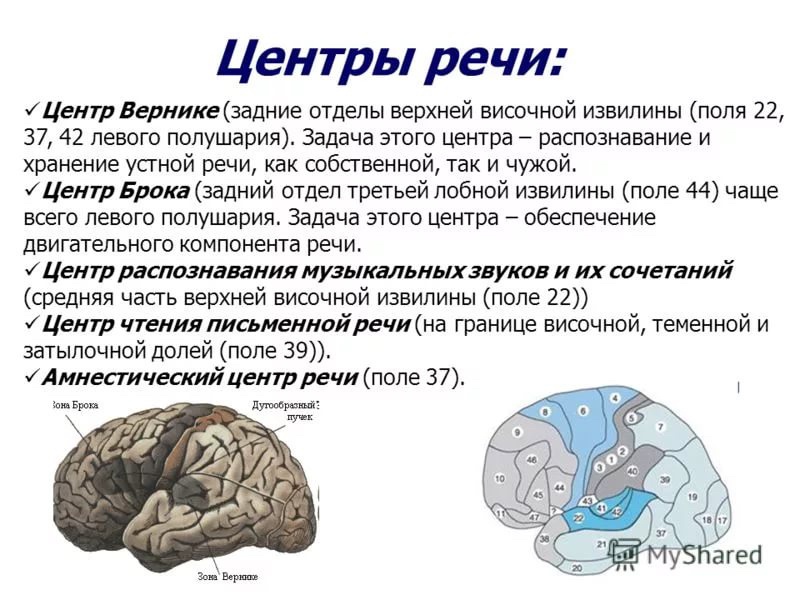
50. Речь как когнитивная функция. Система слухоречевых зон коры. Физиологическую основу речи составляет вторая сигнальная система, условными раздражителями которой являются слова в их звуковой (устная речь) или зрительной форме (письменная речь). Звуки и начертания слов, будучи вначале для отдельного человека нейтральными раздражителями, становятся условными речевыми раздражителями в процессе повторного сочетания их с первосигнальными раздражителями, вызывающими восприятия и ощущения предметов и их свойств. В результате - приобретают смысловое значение, становятся сигналами непосредственных раздражителей, с которыми сочетались. Образовавшиеся при этом временные нервные связи в дальнейшем укрепляются путем постоянных речевых подкреплений, делаются прочными и приобретают двусторонний характер: вид предмета немедленно вызывает реакцию его называния, и, наоборот, слышимое или видимое слово сейчас же вызывает представление обозначаемого этим словом предмета. Системы, обеспечивающие речь, могут быть разделены на две группы: периферические и центральные. К центральным относятся определенные структуры головного мозга, а к периферическим — голосовой аппарат и органы слуха. Все речевые анализаторы закладываться в обоих полушариях, но развиваются только с одной стороны (у правшей – слева, у левшей – справа). Эта зона состоит из 3-х отделов.
51. Цикл «бодрствование-сон» Биоритмы - циклические изменения функций органов, систем и организма в целом. Главная характеристика циклической активности - периодичность, т.е. время, за которое совершается один полный цикл. В соответствии с временем периодов циклической активности все биологические ритмы делятся на три группы: 1)циркадианные (ритмы с суточной периодичностью. К циркадианным ритмам относятся циклы сон-бодрствование, циклические суточные колебания температуры тела, содержания электролитов в жидкостях организма, уровня гормонов в крови и т.д). Эндогенный период цикла "сон - бодрствование" - около 25 ч, но в норме он подчиняется смене дня и ночи. Однако цикл нарушается в условиях изоляции человека от внешней среды либо при быстрой смене часового пояса. Важно различать реакцию функциональных систем на изменение окружающей среды или поведения (например, повышение АД и ЧСС при переходе в вертикальное положение) и те функциональные сдвиги, которыми управляет внутренний водитель циркадианного ритма (например, суточные колебания уровня кортизола в плазме , не зависящие от внешних воздействий). 2)инфрадианные (циклы с большей, чем сутки, длительностью - недельные, месячные, годовые ритмы. Ими являются менструальный цикл, сезонные изменения функций организма, жизненный цикл); 3)ультрадианные (ритмы с периодичность меньше суток - часовые, минутные, секундные ритмы. К этой группе биоритмов относятся потребление пищи, периодическая деятельность органов пищеварения, дыхание, сердечный ритм, ритмы активности различных клеток-пейсмекеров и т.д.).
52. Структура сна: фазы, стадии. Характеристика стадий сна: изменения на электроэнцефалограмме, вегетативные, соматические и психические проявления. Открытие Е. Азеринским и Н. Клейтманом явления «быстрого», или «парадоксального», сна, во время которо го были обнаружены быстрые движения глазных яблок (БДГ) при закрытых веках и общей полной мышечной релаксации послужило основанием для современных исследований физиологии сна. Сон представляет собой совокупность двух чередующихся фаз: 1) «медленного», или «ортодоксального», сна и 2) «быстрого», или «парадоксального», с быстрым движением глаз сна (Rapid Eyes Movements – REM-сон или БДГ-сон). Название этих фаз сна обусловлено характерными особенностями ЭЭГ: во время «мед ленного» сна регистрируются преимущественно медленные волны, а во время «быстрого» сна — быстрый бета-ритм, характерный для бодрствования человека, что дало основание называть эту фазу сна «парадоксальным» сном. На основании электроэнцефалографической картины фазу «медленного» сна в свою очередь подразделяют на несколько ста дий. Выделяют следующие основные стадии сна: стадия I — дремота, процесс погружения в сон. Для этой ста дии характерна полиморфная ЭЭГ, исчезновение альфа-ритма. В течение ночного сна эта стадия обычно непродолжительна (1 — 7 мин). Иногда можно наблюдать медленные движения глазных яблок (МДГ), при этом быстрые их движения (БДГ) полностью отсутствуют; стадия II – стадия легкого сна характеризуется появлением на ЭЭГ так называемых сонных веретен (12—18 в секунду) и К-комплексов (вертекс-потенциал с последующим «сонным веретеном»). Эта стадия является наиболее продолжительной из всех; она мо жет занимать около 50 % времени всего ночного сна. Движения глаз не наблюдаются; стадия III - стадия умеренно глубокого сна. Характеризуется присутствием К-комплексов и наличием и «сонных веретен». Появляются дельта-волны. стадия IV – стадия глубокого сна - преобладают высокоамплитудные (75мкВ) дельта-волны частотой 0,5-1,2 Гц. На стадию приходится 20-30% общей продолжительности сна. Стадия БДГ - «быстрого», или «парадоксального», сна характеризуется наличием десинхронизированной смешанной ак тивности на ЭЭГ: быстрые низкоамплитудные ритмы (по этим проявлениям напоминает стадию I и активное бодрствование — бета-ритм), которые могут чередоваться с низкоамплитудными медленными и с короткими вспышками альфа-ритма, пилообраз ными разрядами, БДГ при закрытых веках. Занимает 15-25% общей продолжительности сна. К концу ночи в последних циклах продолжительность быстрого сна увеличивается. Получается, что у спящего при полной обездвиженности мозг чрезвычайно активен. Из-за этого сочетания признаков РЕМ-сон получил свое второе название — «парадоксальный сон».
Ночной сон обычно состоит из 4-5 циклов, каждый из кото рых начинается с первых стадий «медленного» сна и завершается «быстрым» сном. Длительность цикла у здорового взрослого чело века относительно стабильна и составляет около 90 мин. В первых двух циклах преобладает «медленный» сон, в последних — «быст рый», а «дельта»-сон резко сокращен и даже может отсутствовать. Продолжительность «медленного» сна составляет 75-85%, а «парадоксального» — 15-25 % от общей продолжительности ночного сна. Мышечный тонус во время сна. На протяжении всех стадий «медленного» сна тонус скелетной мускулатуры прогрессивно па дает, в «быстром» сне мышечный тонус отсутствует. У спящего почти полностью пропадает тонус мыщц — так называемая атония, остаются работать только мышцы слуховых косточек, глазодвигательные мышцы и диафрагма. Вегетативные сдвиги во время сна. Во время «медленного» сна замедляется работа сердца, уменьшается частота дыхания, воз можно возникновение дыхания типа Чейна — Стокса, по мере углубле ния «медленного» сна может быть частичная обструкция верхних дыхательных путей и появление храпа. Секреторная и моторная функции пищеварительного тракта по мере углубления «медлен ного» сна уменьшаются. Температура тела перед засыпанием снижается и по мере углубления «медленного» сна это снижение про грессирует. Считают, что снижение температуры тела может являться одной из причин наступления сна. Пробуждение сопро вождается повышением температуры тела.
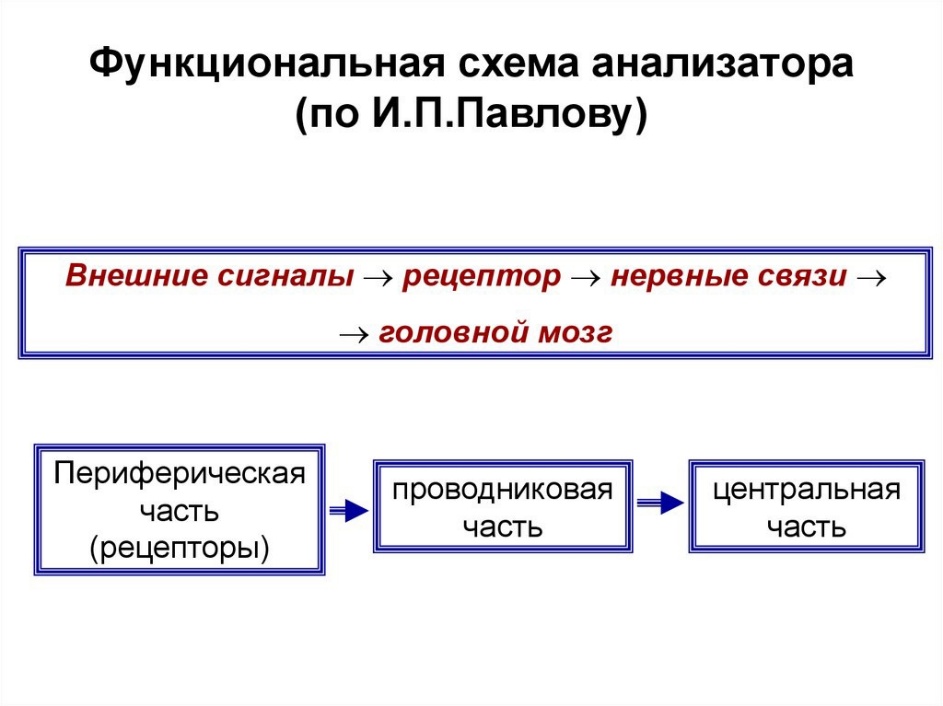
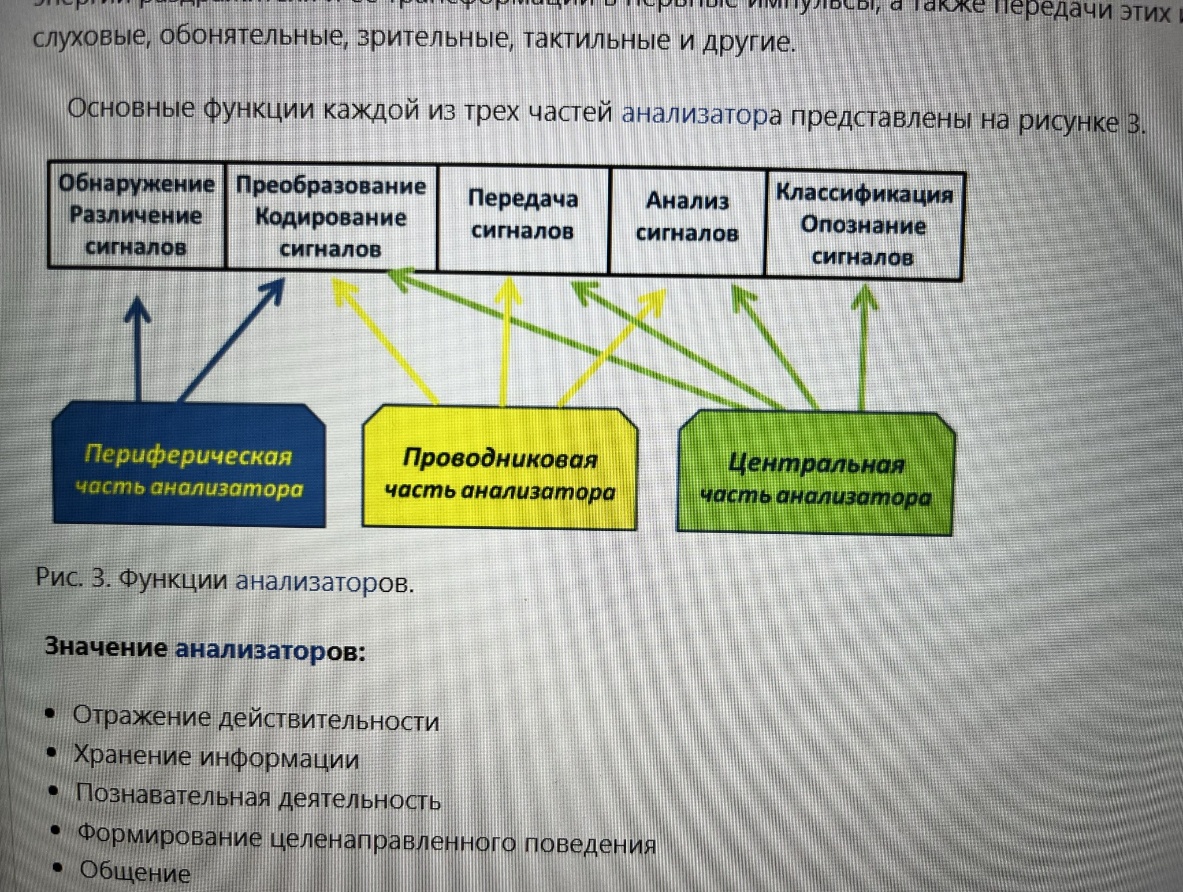
53. Анализатор: определение. Структурно-функциональная организация анализатора. Анализатор — функциональная единица, отвечающая за восприятие и анализ сенсорной информации одного вида (термин ввел И. П. Павлов). Анализатор представляет собой совокупность нейронов, участвующих в восприятии раздражений, проведении возбуждения и в анализе раздражения. Анализатор – нервный аппарат, осуществляющий получение и анализ информации о внешней и внутренней среде организма и формирующий специфические (для данного анализатора) ощущения и восприятие в целом. Орган чувств – периферическое образование, воспринимающее изменение окружающей среды Сенсорная система – совокупность центров и периферических образований НС, обеспечивающих восприятие, анализ информации из внешней и внутренней среды (соматовисцеральная, зрительная, звуковая, вестибулярная, вкусовая, обонятельная) Отделы: 1. Периферический – рецептор – восприятие сигналов внешней и внутренней среды 2. Проводниковый – чувствительный нервные волокна, проводящие сигналы от рецепторов к ЦНС 3. Центральный (корковый) – структуры ЦНС, включая кору больших полушарий головного мозга, которые анализируют эти сигналы Общие принципы строения сенсорных систем: 1. Многослойность - быстрая реакция на простые сигналы, анализ уже на первых уровнях сенсорной системы. Наличие нескольких слоев нервных клеток: a. Первый слой связан с рецепторами b. Последний – с нейронами моторных областей коры БМ 2. Многоканальность – детальность анализа сигналов, большая надежность. Наличие множества клеток в каждом слое, связанных с множеством клеток следующего слоя 3. «Сенсорные воронки»: a. Сужающаяся – схождение сигналов от большего числа клеток к меньшему – уменьшение избытка информации b. Расширяющаяся – наоборот – обеспечение дробного и сложного анализа разных признаков сигнала 4. Дифференциация сенсорной системы: a. По вертикали – образование отделов, каждый из которых состоит из нескольких нейронных слоев (каждый отдел выполняет определенную функцию) b. По горизонтали – различные свойства рецепторов, нейронов и связей между ними в пределах каждого из слоев (разграничение типов информации) Общие свойства сенсорных систем: 1. Высокая чувствительность к адекватным раздражителям 2. Наличие дифференциальной чувствительности 3. Адаптация 4. Тренируемость 5. Сохранение ощущения после прекращения действия раздражителя – последствие, последовательные образы 6. Постоянные взаимодействия анализаторов в условиях нормального функционирования
54. Свойства анализаторов. Основной психофизический закон На основе наблюдений Вебера Густав Фехнер в 1860 году сформулировал «основной психофизический закон» восприятия
Общие свойства сенсорных систем: 1. Высокая чувствительность к адекватным раздражителям 2. Наличие дифференциальной чувствительности 3. Адаптация 4. Тренируемость 5. Сохранение ощущения после прекращения действия раздражителя – последствие, последовательные образы 6. Постоянные взаимодействия анализаторов в условиях нормального функционирования Многослойность - быстрая реакция на простые сигналы, анализ уже на первых уровнях сенсорной системы. Наличие нескольких слоев нервных клеток: a. Первый слой связан с рецепторами b. Последний – с нейронами моторных областей коры БМ 2. Многоканальность – детальность анализа сигналов, большая надежность. Наличие множества клеток в каждом слое, связанных с множеством клеток следующего слоя 3. «Сенсорные воронки»: a. Сужающаяся – схождение сигналов от большего числа клеток к меньшему – уменьшение избытка информации b. Расширяющаяся – наоборот – обеспечение дробного и сложного анализа разных признаков сигнала 4. Дифференциация сенсорной системы: a. По вертикали – образование отделов, каждый из которых состоит из нескольких нейронных слоев (каждый отдел выполняет определенную функцию) b. По горизонтали – различные свойства рецепторов, нейронов и связей между ними в пределах каждого из слоев (разграничение типов информации)


55. Рецепторы: определение, классификация, свойства Рецепторы-Специализированные образования, участвующие в трансформации энергии раздражение в нервные импульсы. Классификация рецепторов: 1. От вида адекватных для них раздражителей (механо-, фото-, термо-, хемо-, баро-, осмо-) 2. По качеству ощущений (зрительные, обонятельные итд) 3. По дальности расположения воспринимаемого стимула (Дистантные – слух, зрение, Контактные – осязание, вкус, обоняние) 4. По локализации (экстеро-, интероцепторы) 5. По происхождению (первичные: обонятельные, болевые, термо-, большинство механо- кожи, проприо- и большинство интероцепторов внутренних органов – сами генерируют потенциал действия, вторичные: вкусовые, слуховые, вестибулярные, хемочувствительные клетки синокаротидного клубочка – реагируют на раздражитель возникновением РП и выделяют медиатор, действующий на сенсорные нейроны) ФОТОРЕЦЕПТОРЫ – общее происхождение с нервными клетками (первичный рецептор), но отсутствие способности генерировать ПД (вторичный рецептор) Свойства рецепторов: 1. Высокая возбудимость 2. Адаптация 3. Спонтанная активность (возбуждение без раздражителя) 4. Специфичность 5. Избирательная чувствительность 6. Преобразовательность (химия – электричество) 7. Функциональная лабильность (люди с севера имеют больше холодовых рец, чем тепловых)
56. Механизмы трансдукции первично и вторично чувствующих рецепторах. Рецепторы-Специализированные образования, участвующие в трансформации энергии раздражение в нервные импульсы. Когда стимул взаимодействует с рецептором, его энергия запускает молекулярно-биологические процессы, которые объединяются под общим названием – трансдукция. В результате этих процессов образуется энергия, под влиянием которой изменяется мембранная проницаемость мембраны для ионов. Происходит изменение потенциала покоя клетки и образуется рецепторный потенциал (РП). Важно отметить, что на рецепторной мембране нет потенциалзависимых ионных каналов, т.е. она не возбудима и ПД не возникает. РП не подчиняется закону «все или ничего», носит градуальный характер ответа, зависит в логарифмическом порядке от величины раздражителя, может быть деполяризационным или гиперполяризационным, распространяется с декрементом. Дальнейшее образование нервного импульса происходит по-разному в первичночувтвующем и вторичночувствующем рецепторе Рецепторный акт можно представить в следующем виде: • I этап – специфическое взаимодействие раздражителя с рецептирующим субстратом рецептора; • II этап – образование РП в месте взаимодействия стимула с рецептором (следствие изменения проницаемости плазмолеммы рецептора для ионов); • III этап – генерация ПД: — в первичночувствующих рецепторах ПД возникает вследствие электротонического распространения РП (пороговой величины) на возбудимые участки плазмолеммы рецепторной клетки; — во вторичночувствующих рецепторах РП распространяется по плазмолемме специальной клетки от места взаимодействия рецептора со стимулом к синаптическому контакту с аксоном; происходит выделение медиатора и синаптическая передача; на постсинаптической мембране возникает постсинаптический (генераторный, ГП) потенциал, который достигнув возбудимого участка мембраны, приводит к генерации ПД; — ПД по нервным волокнам устремляются без декремента в центральную нервную систему. Таким образом, сенсорный рецептор осуществляет преобразование энергии действующего раздражителя (механического, химического, светового, теплового и др.) в специфическую энергию нервной системы, т.е. в упорядоченную последовательность нервных импульсов. Нервные импульсы – это универсальные сигналы, заключающие в себе многогранную информацию об окружающей среде. Только в таком виде эту информацию может воспринять центральная нервная система, чтобы объединить отдельные свойства внешнего мира в его целостный образ и таким образом объективно отобразить действительность.
57. Структурно-функциональная организация зрительного анализатора. Зрительная сенсорная система, как и любая другая, состоит из трех отделов: 1. Периферический отдел – глазное яблоко, в частности — сетчатка глаза (воспринимает световое раздражение) 2. Проводниковый отдел — аксоны ганглиозных клеток — зрительный нерв — зрительный перекрест — зрительный тракт — промежуточный мозг (коленчатые тела) — средний мозг (четверохолмие ) — таламус 3. Центральный отдел — затылочная доля: область шпорной борозды и прилегающих извилин. Зрительный путь обеспечивает проведение световых импульсов от сетчатки до подкорковых и корковых центров, участвует в обеспечении ответной реакции на зрительные раздражения. От палочек и колбочек , которые находится в девятом слой сетчатки, импульсы поступают периферическом отростком и биполярных клеток ( 1 нейрон) Центральные отростки биполярных клеток заканчиваются на мультиполярных нервных клетках сетчатки (2 нейрон) Аксоны ганглиозных клеток формируют нерв, который идёт к перекресту Там перекрещиваются только 2/3 волокон, расположенных медиально, несущих инф с латеральных сторон зрительного поля, неперекрещенные направляются в зрительный тракт своей стороны. Зрительный перекроет продолжается в тракт, который проводит импульсы к подкорковым центрам (3 нейроны) а) От верхних холмиков импульсы поступают для обеспечения зрачков в рефлексы (изменение ширины и зрачка при разном освещение), часть Оксанов верхнего холмика направляется в интеграционный центр среднего мозга. б) От нейронов задних ядер таламуса импульсы поступают на медиальная ядра таламуса, которые являются подкорковых чувствительным центром экстрапирамидной системы, который обеспечивает изменение тонуса мускулатуры в ответ на зрительные раздражение в) От латеральных коленчатых тел зрительные информация проходит через заднюю ножку внутренней капсулы и затем в виде radiatio optica к шпорной борозде - проекционный центр зрения (4нейрон)
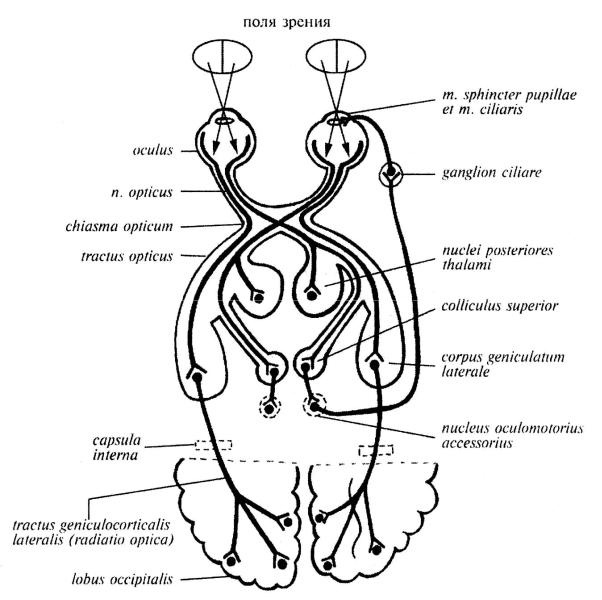
58.Острота зрения Острота зрения - Способность глаза воспринимать две точки, расположенные на минимальном расстоянии друг от друга, как отдельные. Зависит от освещенности, контрастности, формы объекта и других факторов. В странах СНГ определяется долями единицы: 1,0 — нормальное зрение, 0,9; 0,8, и т. д. до 0,1 — определяется количеством строк начиная с верхней по таблице Сивцева или Головина с расстояния 5 метров. Исследования проводят для каждого глаза отдельно: сначала определяют остроту зрения одного, затем другого глаза. V = d / D, где V — острота зрения; d — расстояние, с которого проводится исследование; D — расстояние, на котором нормальный глаз видит данный ряд.
59. Строение и функции оптическая системы глаза. Рефракция. Нарушения рефракции. ???? Глаз человека представляет собой сложную оптическую систему, которая состоит из роговицы, влаги передней камеры, хрусталика и стекловидного тела Преломляющая сила глаза зависит от величины радиусов кривизны передней поверхности роговицы, передней и задней поверхностей хрусталика, расстояний между ними и показателей преломления роговицы, хрусталика, водянистой влаги и стекловидного тела. Оптическую силу задней поверхности роговицы не учитывают, поскольку показатели преломления ткани роговицы и влаги передней камеры одинаковы (как известно, преломление лучей возможно лишь на границе сред с различными коэффициентами преломления). Для оценки преломляющей способности любой оптической системы используют условную единицу - диоптрию (сокращенно - дптр). За 1 дптр принята сила линзы с главным фокусным расстоянием в 1 м. Диоптрия - величина, обратная фокусному расстоянию: где D - диоптрия; F - фокусное расстояние. Следовательно, линза с фокусным расстоянием 0,5 м обладает преломляющей силой 2,0 дптр, 2 м - 0,5 дптр и т. д. Преломляющую силу выпуклых (собирающих) линз обозначают знаком «плюс», вогнутых (рассеивающих) - знаком «минус», а сами линзы называют соответственно положительными и отрицательными. Существует простой прием, с помощью которого можно отличить положительную линзу от отрицательной. Для этого линзу нужно расположить на расстоянии нескольких сантиметров от глаза и передвигать ее, например, в горизонтальном направлении. При рассматривании какого-либо предмета через положительную линзу его изображение будет смещаться в сторону, противоположную движению линзы, а через отрицательную, наоборот, - в ту же сторону. Как и другим оптическим системам, глазу свойственны различные аберрации (от лат. aberratio - отклонение) - дефекты оптической системы глаза, приводящие к снижению качества изображения объекта на сетчатке. Вследствие сферической аберрации лучи, исходящие из точечного источника света, собираются не в точке, а в некоторой зоне на оптической оси глаза. В результате этого на сетчатке образуется круг светорассеяния. Глубина этой зоны для «нормального» человеческого глаза колеблется от 0,5 до 1,0 дптр. Рефракция – преломление света в оптической системе. ВИДЫ РЕФРАКЦИИ ГЛАЗА Для хорошего зрения необходимо прежде всего четкое изображение рассматриваемого предмета на сетчатке. В здоровом глазу человека это зависит от соответствия параметров двух анатомических элементов глаза: преломляющей силы оптической системы и длины оптической оси глаза. Каждый из этих параметров имеет выраженные инди видуальные колебания. В связи с этим в понятии «рефракция глаза» принято выде лять физическую рефракцию, характеризующую преломляющую силу опти ческой системы глаза, и клиническую рефракцию. Физическая рефракция глаза взрослого человека варьирует в широких преде лах — от 52 до 71 дптр, составляя в среднем 60 дптр. Она формируется в период роста глаза и в дальнейшем не меняется. В практической деятельности офтальмолог определяет чаще клиническую реф ракцию. Клиническую рефракцию характеризует положение главного фокуса по отношению к сетчатке. Если главный фокус совпадает с сетчаткой, такая рефракция называется соразмерной — эмметропией (Е)'. Если главный фокус не совпадает с сетчаткой, то клиническая рефракция несо размерная— аметропия. Преломляющая сила оптического аппарата может быть слишком сильной для данного размера глаза, и тогда параллельные лучи собирают ся перед сетчаткой. Такой вид несоразмерной рефракции называется близоруко стью — миопией (М)2. Если преломляющая сила по отношению к размеру глаза будет слабой, то главный фокус расположится за сетчаткой. Этот вид несоразмерной рефракции называется дальнозоркостью — гиперметропией (Н)3.
60.Механизм аккомодации Механизм аккомодации: Аккомодация – Способность глаза приспосабливаться к смене расстояния, на котором расположен предмет. Она происходит автоматически при перемене внимание близко расположены объекты на удалённый. Чем эластичнее хрусталик, тем быстрее происходит процесс. Механизм: сокращение ресничной мышцы, расслабление цилиарной связки, на которую подвешен хрусталик – он становится выпуклым, преломляющая сила усиливается, на сетчатке фокусируется изображение близкорасположенных предметов. При расслаблении ресничной мышцы все наоборот. Сужение зрачка – парасимпатическая иннервация
61. Характеристика фоторецепторов Сетчатка представляет собой внутреннюю оболочку глаза, имеющую сложную многослойную структуру. Здесь расположены два вида различных по своему функциональному значению фоторецепторов — палочки и колбочки и несколько видов нервных клеток с их многочисленными отростками. Под влиянием световых лучей в фоторецепторах происходят фотохимические реакции, состоящие в изменении светочувствительных зрительных пигментов. Это вызывает возбуждение фоторецепторов и затем синаптическое возбуждение связанных с палочками и колбочками нервных клеток. Последние образуют собственно нервный аппарат глаза, который передает зрительную информацию в центры головного мозга и участвует в ее анализе и переработке. Сетчатка является как бы частью мозга, вынесенной на периферию. Пигментный слой сетчатки. Наружный слой сетчатки образован пигментным эпителием, содержащим пигмент фусцин. Этот пигмент поглощает свет, препятствуя его отражению и рассеиванию, что способствует четкости зрительного восприятия. Пигментные клетки, отростки которых окружают светочувствительные членики палочек и колбочек, принимают участие в обмене веществ в фоторецепторах и в синтезе зрительного пигмента. Фоторецепторы. К слою пигментного эпителия изнутри примыкает слой фоторецепторов, которые своими светочувствительными члениками обращены в сторону, противоположную свету. Каждый фоторецептор — палочка или колбочка — состоит из чувствительного к действию света наружного сегмента, содержащего зрительный пигмент, и внутреннего сегмента, содержащего ядро и митохондрии, обеспечивающие энергетические процессы в фоторецепторной клетке. У человека в глазу имеется около 6-7 млн. колбочек и 110-125 млн. палочек. Палочки и колбочки распределены в сетчатке неравномерно. Центральная ямка сетчатки (fovea centralis) содержит только колбочки (до 140 000 колбочек на 1 мм2). По направлению к периферии сетчатки число колбочек уменьшается, а количество палочек возрастает. Периферия сетчатки содержит почти исключительно палочки. Колбочки функционируют в условиях яркой освещенности и воспринимают цвета; палочки являются рецепторами, воспринимающими световые лучи в условиях сумеречного зрения. Раздражение различных участков сетчатки показывает, что различные цвета воспринимаются лучше всего при действии световых раздражителей на центральную ямку, где расположены почти исключительно колбочки. По мере удаления от центра сетчатки восприятие цвета становится все хуже. Периферия сетчатки, где находятся исключительно палочки, не воспринимает цвета. Световая чувствительность колбочкового аппарата сетчатки во много раз меньше таковой элементов, связанных с палочками. Поэтому в сумерках, в условиях малой освещенности, центральное колбочковое зрение резко понижено и преобладает периферическое палочковое зрение. Т.к. палочки не воспринимают цвета - в сумерках человек цвета не различает. Нарушение функций палочек, возникающее, например, при недостатке в пище витамина А, вызывает расстройство сумеречного зрения - куриную слепоту: человек слепнет в сумерках, но днем зрение остается нормальным. При поражении колбочек возникает светобоязнь: человек видит только при слабом свете и слепнет при ярком освещении. В этом случае может развиться и полная слепота на цвета — ахромазия. Слепое пятно. Место входа зрительного нерва в глазное яблоко не содержит фоторецепторов и нечувствительно к свету. Нейроны сетчатки. Кнутри от слоя фоторецепторных клеток в сетчатке расположен слой биполярных нейронов, к которым изнутри примыкает слой ганглиозных нервных клеток. Аксоны ганглиозных клеток образуют волокна зрительного нерва. Таким образом, возбуждение, возникающее в фоторецепторе при действии света, передается на волокна зрительного нерва через нервные клетки — биполярные и ганглиозные.
В синапсах между биполярными и ганглиозными клетками выявлена холинэстераза; это служит указанием на то, что передача импульса с одной клетки на другую совершается с помощью медиатора ацетилхолина. На 130 млн. фоторецепторных клеток приходится всего около 1 млн. 250 тыс. волокон зрительного нерва, являющихся отростками ганглиозных клеток. Импульсы от многих фоторецепторов конвергируют (сходятся) к одной ганглиозной клетке. Один биполярный нейрон связан со многими палочками и несколькими колбочками, а одна ганглиозная клетка в свою очередь связана со многими биполярными клетками. Каждая ганглиозная клетка суммирует возбуждение, возникающее в большом числе фоторецепторов. Лишь в центре сетчатки, в районе центральной ямки, каждая колбочка соединена с одной так называемой карликовой биполярной клеткой, с которой соединена также всего одна ганглиозная клетка. Фоторецепторы, соединенные с одной ганглиозной клеткой, образуют рецептивное поле ганглиозной клетки. Рецептивные поля различных ганглиозных клеток частично перекрывают друг друга. Взаимодействие соседних нейронов сетчатки обеспечивается горизонтальными и амакриновыми клетками, отростки которых соединяют по горизонтали биполярные и ганглиозные клетки. Амакриновые клетки осуществляют процесс горизонтального (бокового) торможения между соседними элементами. Кроме афферентных волокон, в сетчатке имеются и центробежные (эфферентные) нервные волокна, приносящие к ней импульсы из ЦНС. Эти импульсы действуют на синапсы между биполярными и ганглиозными клетками сетчатки и тем самым регулируют проведение возбуждения между ними. Второй тип центробежных нервных волокон представляет собой сосудодвигательные волокна, изменяющие кровоснабжение сетчатки.
62.Механизм трансдукции светового сигнала в сетчатке. Рецепторные поля сетчатки. Процесс преобразования (трансдукции) зрительных сигналов связан с гиперполяризацией палочек и колбочек. Передача зрительных сигналов отличается от обычного способа преобразования сигналов, при котором сенсорные рецепторы деполяризуются. Когда молекула родопсина палочек поглощает свет, сигнал усиливается посредством специального преобразующего механизма. Наличием усиливающего механизма, а также большого количества фотопигмента в наружном сегменте палочек объясняется чрезвычайно высокая их чувствительность: после полной темновой адаптации палочка реагирует на один фотон (квант света). В темноте натриевые каналы мембраны палочек открыты. Поэтому в эти клетки непрерывно входят ионы Na+ - темновой ток. Он обеспечивает постоянную деполяризацию мембраны палочек (потенциал покоя примерно - 40 мВ). Деполяризация вызывает тоническое (постоянное) высвобождение нейротрансмиттера (глутамата) из синапсов, образованных палочками на биполярных и горизонтальных клетках. Внутриклеточная концентрация Na+ поддерживается на стационарном уровне благодаря ионному насосу - Na+/К+-АТФазе.
Поглощение света сопровождается активацией G-белка трансдуцина, который активирует фосфодиэстеразу циклического гуанозинмонофосфата (цГМФ). Этот фермент, который находится в содержащих родопсин дисках, гидролизует цГМФ до 5'-ГМФ, снижая концентрацию его в цитоплазме палочек. При нормальной концентрации цГМФ поддерживается открытое состояние натриевых каналов. Его низкая концентрация заставляет натриевые каналы закрываться, и мембрана гиперполяризуется. Усиление сигнала обусловлено способностью одной молекулы родопсина активировать сотни молекул трансдуцина; каждая молекула фосфодиэстеразы гидролизует в секунду тысячи молекул цГМФ. Аналогичные события происходят в колбочках, но гиперполяризация мембраны в них развивается гораздо быстрее, чем в палочках за счет более коротких внутриклеточных расстояний. Рецептивное поле сетчатки - - это зона внутри которой зрительный стимул вызывает соответствующий ответ ганглионарных клеток. В сетчатке человека имеются два типа ганглиозных клеток, отличающихся реакцией на точечные световые стимулы, воздействовавшие на центр или периферию их рецептивного поля (рис. 17.9). Примерно половина ганглиозных клеток возбуждается действием света на центр рецептивного поля и тормозится при действии светового стимула на периферию рецептивного поля. Такие клетки принято называть оn-нейронами. Другая половина ганглиозных клеток возбуждается действием светового раздражителя на периферию рецептивного поля и тормозится в ответ на световую стимуляцию центра рецептивного поля — они получили название off-нейронов.
63. Поля зрения. Поле зрения - пространство, которое видит глаз человека при фиксации взора в одной точке и неподвижной голове. В зависимости от того, участвуют в зрении один или оба глаза, различают монокулярное и бинокулярное поле зрения. Каждый глаз человека имеет в норме поле ахроматического зрения: 55° вверх, 60° вниз, 90° наружу (суммарное поле зрения двумя глазами – 180°) и 60° – внутрь. Ахроматическое поле больше хроматического, т.к. обусловлено деятельностью палочек, число которых больше и которые расположены преимущественно на периферии сетчатки. Для различных цветов поле зрения также неодинаково: наименьший размер поля зрения – у зелёного цвета, наибольший – у синего. В нормальном поле зрения имеются физиологические скотомы (слепые участки): слепое пятно Мариотта, что соответствует проекции диска зрительного нерва, не содержащего фоторецепторы.
64. Свойства зрительного анализатора: инерционность, адаптация. Адаптация – приспособление органа чувства к раздражителям (внешним условиям). При адаптации чувствительность меняется в сторону повышения и в сторону понижения. Например, при переходе от яркого света в темноту мы сначала не видим предметов, но постепенно начинаем различать их очертания (адаптация к темноте). При слабой освещенности развивается темновая адаптация, а при сильной — световая адаптация зрительного анализатора. По мере развития темновой адаптации (то есть адаптации к темноте) абсолютная световая чувствительность возрастает. Ее максимальное значение достигается примерно за 30–35 минут. Именно в таком состоянии у человека возникает ощущение света под действием примерно 10 фотонов (по современным оценкам, от 8 до 50). Световая адаптация зрительной сенсорной системы выражается в уменьшении световой чувствительности при усилении освещенности. Этот процесс развивается гораздо быстрее — за 15–60 секунд. Временны́е различия примерно в два порядка обусловлены разными скоростями фотолиза и ресинтеза родопсина. При изменении уровня освещенности сетчатки включаются безусловно-рефлекторные механизмы, обеспечивающие адаптационные процессы. Так, при переходе человека из светлого помещения в темное прежде всего происходит переход от фотопического к скотопическому зрению — колбочки не реагируют на освещенность менее 0,01 лк, и в таких условиях начинают функционировать палочки, доселе заторможенные (главным образом, по механизму латерального торможения). Это сопровождается увеличением рецептивных полей мультиполярных (ганглиозных) клеток сетчаток, увеличением содержания зрительного пигмента в палочках, рефлекторным расширением зрачка (мидриазом). Длительность темновой адаптации зависит от яркости предварительной засветки глаз (от уровня преадаптации). Чем выше уровень преадаптации, тем дольше тянется переход от фотопического к скотопическому зрению.
65. Теории цветовосприятия. Аномалии цветового зрения. Теории цветового зрения 1) Трехкомпонентная (теория Юнга-Гельмгольца): для восприятия всех цветов, присущих лучам видимой части спектра, достаточно рецепторов трех типов. В сетчатке содержатся колбочки трех видов (S-, M-, L-колбочки), каждый из которых обла дает максимальной чувствительностью к свету с определенной длиной волны. Подобная избирательность связана с тем, что в этих колбочках содержатся фото пигменты трех видов, каждый из которых обладает свойством поглощать свет определенной волны (short, medium, long). Трехкомпонентная теория цветовосприятия объясняет также и такое явление, как комплементарные последовательные образы. При пристальном рассматривании синего квадрата происхо дит избирательная адаптация синих колбочек (их пигмент «истощается»). Когда после этого на центральную ямку проецируется изображение хроматически нейт ральной белой или серой поверхности, активными оказываются только неистощен ные пигменты зеленых и красных колбочек, которые и вызывают дополнительный последовательный образ (возникнет ощущение желтого цвета). Аналогичным образом при стальное всматривание в желтую поверхность вызывает адаптацию колбочек, «от ветственных» за ощущение желтого цвета, а именно красных и зеленых, при этом активными неадаптировавшимися остаются синие колбочки, которые и вызыва ют соответствующий, т. е. синий, комплементарный последовательный образ. На конец, на основе трехкомпонентной теории цветовосприятия можно объяснить и то, почему при одинаковой стимуляции всех фотопигментов мы видим белый цвет. 2) Цветоппонентная (теория Геринга): существует три независимых механизма, в основе каждого из них лежит пара оппонентных процессов (или нейрофизиологи ческих систем) цветоразличения: сине-желтый, зелено-красный и черно-белый. Каждый процесс способен вызвать сенсорные ощущения двух типов, являющиеся антагонистами друг друга. Иными словами, механизм цветовосприятия таков, что каждая пара способна вызвать только одно из двух возможных ощущений, т. е. че ловек видит красный или зеленый цвет, желтый или синий, а не зеленый и красный и не желтый и синий. Теория оппонентных процессов, так же как и трехкомпонентная теория цве тоощущения, находит свое подтверждение в наблюдениях над комплементарными последовательными образами. Если долго смотреть на синюю поверхность, а затем перевести взгляд бесцветную поверх ность, то возникнет комплементарный последовательный образ (т.е. будет ощущение желтого цвета). При одинаковом стимулировании обоих компонентов рецепторной пары они аннулируют действие друг друга (этот процесс аналогичен тому, который происходит при смешении комплементарных цветов, например синего и желтого) и возникает ощущение серого или белого цвета. Аномалии цветовосприятия 1) Монохроматизм: крайне редко встречается; как правило, люди с монохроматизмом вообще не видят никаких цветов, а нередко имеют и другие дефекты зрения. Не исключено, что причиной монохроматизма являются аномалии качественного и количественного состава колбочек в сетчатке. Несмот ря на то, что монохроматизм тоже считается наследственным дефектом, среди монохроматов женщин примерно столько же, сколько и мужчин. 2) Дихроматизм: людям с дихроматизмом для воспроизведения всех цветовых тонов нужны всего два цвета, а не три, как требуется людям с нормальным цветовым зре нием.
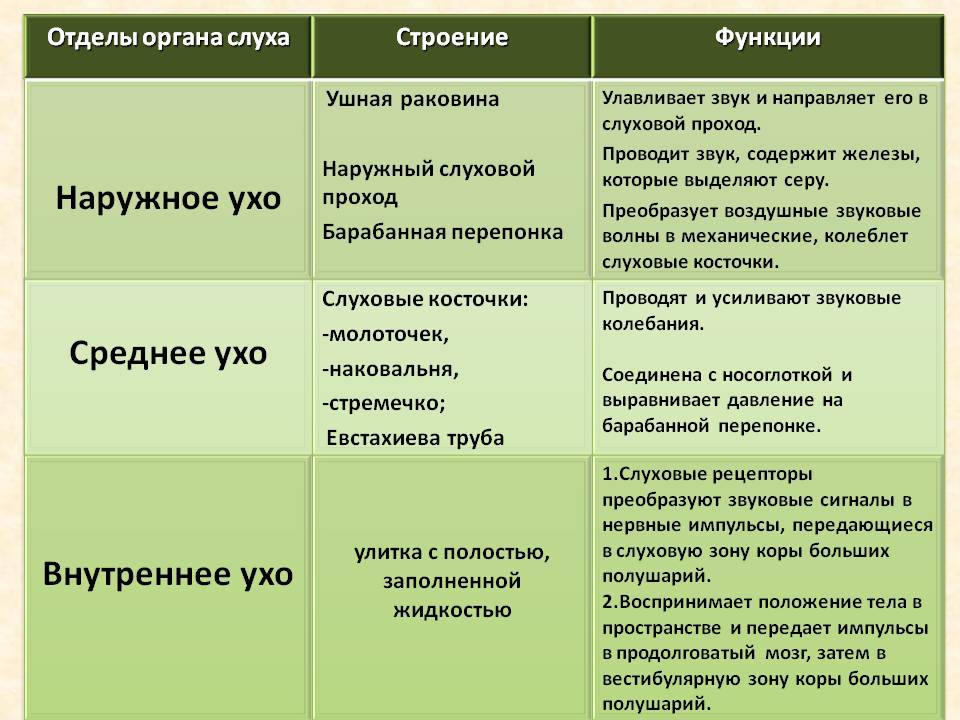
66.Структурно-функциональная схема слухового анализатора. Периферический отдел - механорецепторы-волосковые клетки-улитка-внутреннее ухо-орган слуха-ухо Проводниковый отдел- 8 пара ЧН Центральный отдел-кора височной доли Слуховой путь обеспечивает проведение импульсов до подкорковых и корковых центров, участвуя в обеспечении ответных реакций на звуковые раздражения - от рецепторов (волосковые клетки Кортиева органа) импульсы поступают к клеткам ganglion cochleare (1 нейроны) преддверно- улиткового нерва (ганглий расположен в костном стержне улитки); - центральные отростки биполярных нейронов ganglion cochleare образуют radix cochlearis n. vestibulocochlearis, который вместе с преддверным корешком направляется в мосто-мозжечковый угол; - далее импульсы поступают к nucleus cochlearis anterior et nucleus cochlearis posterior преддверно-улиткового нерва (2 нейроны); аксоны nucleus cochlearis posterior выходят на дорсальную юверхность моста, образуя мозговые (слуховые) полоски четвертого келудочка, striae medullares (acustici) ventriculi quarti, которые огружаются в s. medianus, проходят в составе трапециевидного тела к nucleus posterior corporis trapezoidei противоположной стороны; аксоны nucleus cochlearis anterior направляются к nucleus anterior corporis trapezoidei противоположной стороны, составляя основу трапециевидного тела; - аксоны ядер трапециевидного тела (3 нейроны) формируют lemniscus lateralis; по ее ходу в пределах моста имеются nuclei lemnisci laterales, в которых прерывается часть волокон улитковых ядер, проходящих транзитом через ядра трапециевидного тела; - волокна слуховой петли направляются к подкорковым центрам: нижние холмики среднего мозга, медиальные коленчатые тела, срединные ядра таламуса (4 нейроны) Рецепция звуковых раздражений - барабанная перепонка преобразует звуковые колебания в механические; - механические колебания барабанной перепонки передаются на слуховые косточки; - благодаря наличию двух суставов (art. incudomallearis et art. incudostapedia) срабатывает правило рычага и колебания усиливаются примерно в 80-90 раз; - определенную роль в услении звуковых колебаний и их смягчении играют m. tensortympani et m. stapedius; - колебательные движения стремени в овальном окне приводят к изменению давления перилимфы в преддверии лабиринта; - увеличение давления перилимфы из преддверия лабиринта передается в scala vestibule; - через геликотрему изменение давления распространяется в scala tympani; - колебания перилимфы барабанной лестницы устраняются вторичной барабанной перепонкой (в зависимости от величины давления она изгибается либо в преддверии, либо в барабанную лестницу); - изменения давления в перилимфе приводят к колебательным движениям базилярной и вестибулярной мембран и соответствующим преобразованиям давления в эндолимфе в улитковом протоке; - колебания эндолимфы приводят к смещению покровной мембраны кортикального органа и соответствующему раздражению его волосковых клеток; - высокие звуки вызывают колебания волосковых клеток, расположенных у основания улитки; - низкие звуки воспринимаются волосковыми клетками, расположенными на вершине улитки; - возникшие нервные импульсы переносятся к улитковым ядрам моста по улитковой части вестибулярно-улиткового нерва.
67. Функции вспомогательного аппарата слухового анализатора. Наружное ухо – звукоулавливающий аппарат: раковина и наружный слуховой проход. Наружный слуховой проход подводит колебания среды к другим структурам и поддерживает стабильные условия: температуру и влажность. Считается, что ушная сера поддерживает необходимый диаметр слухового прохода. Барабанная перепонка, это тонкая мембрана, обладающая высокой болевой чувствительностью. Нити, которые ее образуют, направлены в разные стороны, что позволяет улавливать различные частоты. Барабанная перепонка отделяет наружный слуховой проход от среднего уха, в котором так же находится воздух и сохраняется атмосферное давление благодаря евстахиевой трубы. В полости среднего уха расположена цепочка подвижно сочлененных косточек. Ручка молоточка прочно связана с барабанной перепонкой, а основание стремечка помещено в отверстие каменистой части височной кости – овальное окно. Овальное окно в 22 раза меньше по площади, чем барабанная перепонка, поэтому давление в овальном окне выше, чем на барабанной перепонке. Внутреннее ухо – улитка – спирально закрученный костный канал. Имеет 2,5 витка. Внутри костной капсулы две мембраны: базилярная (или основная) и рейснерова. Мембраны разделяют улитку на три лестницы: вестибулярную, барабанную и среднюю. Барабанная и вестибулярная лестницы заполнены перилимфой, а средняя – эндолимфой. Вестибулярная лестница примыкает к овальному окну, а барабанная - заканчивается круглым окном. Перилимфа барабанной лестницы и вестибулярной сообщаются в области геликотремы. А эндолимфа центральной лестницы с ними не сообщается. Содержание калия в эндолимфе повышено, поэтому она заряжена положительно по отношению к перилимфе с повышенным содержанием натрия. Разность потенциалов между эндолимфой и перилимфой называется эндокохлеарным потенциалом (ЭКП). ЭКП = 60-100 мВ. Эндолимфа несет положительный заряд относительно перилимфы. Звуки вызывают колебание ЭКП, амплитуда которых приблизительно 1% (0,6 – 1мВ). Это микрофонный потенциал улитки. Состав эндолимфы обеспечивает сосудистая полоска, расположенная на наружной стенке средней лестницы. Эта структура играет главную роль в удовлетворении энергетических потребностей улитки, поддерживает концентрацию калия в эндолимфе. При недостаточном обеспечении кислородом сосудистой полоски может возникнуть тугоухость, т.к. в этом случае снижается ЭКП. На базилярной мембране расположен рецептивный орган – кортиев орган. В кортиевом органе расположены рецепторные клетки. Это наружные и внутренние волосковые клетки, около 25000 штук. Это типичные вторичночувствующие рецепторы. Наружные волосковые клетки расположены в 3-5 рядов, их около 20000 штук, а внутренние в один ряд – 3500 штук. Волосковые клетки расположены между двух мембран: основной (базилярной) и покровной (текториальной) мембранами. При прохождении звукового стимула происходит колебание эндолимфы, что приводит к сдвигу мембран относительно друг друга и волоски клеток изгибаются
68. Механизм трансдукции сигнала в слуховых рецепторах. Кодирование частоты и интенсивности звука. Теория «бегущей волны» Д. Бекеши. Теория бегущей волны - в ответ на звуковой раздражитель внутри улитки возникает быстрая волна, распространяющаяся от основания до верхушки вдоль основной мембраны. Расстояние, которое проходит эта волна по мембране, определяется частотой колебания стремени. Волна от высоких звуков проходит меньшее расстояние и вызывает максимальную деформацию базилярной мембраны, и максимальное раздражение волосковых клеток, преимущественно в области основного завитка улитки. Волна от низких звуков способна проходить на большие расстояния и вызывать деформацию мембраны по всей ее длине. Ощущение высоты звука определяется участком максимальной амплитуды колебаний базилярной мембраны. Чем выше звук, чем больше частота колебаний, воспринимаемых ухом, тем меньше длина колеблющегося столба жидкости в каналах улитки и тем ближе к основанию улитки и овальному окну максимальная амплитуда колебаний. При низкочастотных звуках максимальная амплитуда колебаний приходится на вершину улитки. Механизм трансдукции (рецепции) звука: 1)волоски рецепторной волосковой клетки отгибаются в сторону, когда упираются в покровную мембрану, поднимаясь к ней вместе с базальной мембраной; 2)из-за этого растягивается клеточная мембрана волоска, в ней открываются ионные каналы для натрия (Na+). Механочувствительные ионные каналы (стретч-каналы), открываемые напрямую растяжением клеточной мембраны; 3)ионы Nа+ через открывшиеся для них каналы устремляются внутрь клетки; 4)они приносят с собой положительные электрические заряды (+), вызывают уменьшение электроотрицательности внутри клетки - процесс деполяризации. Электроотрицательность рецепторных волосковых клеток уменьшается, поляризация мембраны снижается, рецепторные клетки переходят в возбуждённое состояние; 5)в ответ на деполяризацию открываются другие каналы - потенциал-управляемые ионные каналы для Ca2+. В рецепторных клетках в отличие от обычных нейронов появляются «новые действующие лица» - кальциевые каналы, чувствительные к деполяризации. При деполяризационном возбуждении эти каналы открываются и впускают в рецепторную клетку ионы кальция; 6)через открытые деполяризацией потенциал-зависимые ионные каналы Ca2+ поступает в клетку. Кальций связывается со специальным белком и побуждает пузырьки с медиатором двигаться к мембране и выбрасывать медиатор наружу. Без кальция ничего бы не вышло: медиатор не выделился бы; 7)из рецепторной клетки под действием вошедшего в неё кальция начинает выделяться нейромедиатор. Нейромедиатор — вещество, передающее возбуждение на связанный с рецепторной волосковой клеткой биполярный нейрон. Как нейромедиатор передаст возбуждение? Он заставит биполярный нейрон породить нервный импульс.
69. Основы физиологической акустики. Сопоставление физических и субъективных (психофизических) характеристик звука. ????? Физические характеристики: 1. Гармонический спектр a. Основная гармоника b. Высшая гармоника 2. Интенсивность (плотность потока энергии) Психофизические характеристики: 1. Акустический спектр a. Основной тон (высота) b. Овертон (тембр) 2. Громкость


70. Строение и функции вестибулярного анализатора. Рецептивный отдел: волосковые клетки вестибулярного органа, расположенными в пирамиде височной кости Проводниковый отдел: 1)биполярные клети в вестибулярных ганглиях 2)вестибулярные ядра продолговатого мозга 3)ядра таламуса Корковый отдел: задняя постцентральная извилина коры Воспринимающей частью вестибулярной системы является вестибулярный аппарат, составляющий вместе с улиткой внутреннее ухо – лабиринт. Лабиринт разделяют на три отдела: передний – улитку, средний – преддверие и задний – полукружные каналы. В полукружных каналах на кристах расположены рецепторы, воспринимающие угловые ускорения, а в преддверии на макулах – рецепторы, воспринимающие линейные ускорения и вибрацию. Макула утрикулюса (маточка) лежит приблизительно в горизонтальной плоскости, а макула саккулюса (мешочек) расположена вертикально. Рецепторы вестибулярного аппарата являются вторичночувствующими механорецепторами. Они образованы волосковыми клетками. Каждая клетка содержит 50 – 60 коротких волосков - стереоцилии, и один длинный – киноцилия. Волоски сенсорных клеток полукружных каналов покрыты колпачком – купулой, которая полностью перекрывает просвет расширенной ампулярной части канала. Купула представляет собой желотинообразную мембрану. Вместе с кожным, зрительным и двигательным анализаторами вестибулярный анализатор обеспечивает поддержание равновесия при разнообразных движениях и участвует в ориентировочных реакциях организма в пространстве (анализатор равновесия и гравитации): - от рецепторов статокинетического анализатора (ампулярные гребешки и отолитовы аппараты внутреннего уха) импульсы поступают кganglion vestibulare преддверно-улиткового нерва (1 нейроны); - далее в составе radix vestibularis преддверно-улиткового нерва они входят в мосто-мозжечковый угол и внедряются в вещество моста (рис. 35), направляясь к вестибулярным ядрам (2 нейроны); - аксоны клеток ядер Дейтерса и Швальбе переходят на противоположную сторону, формируя преддверно-таламический тракт, tr. vestibulothalamicus; - в составе бульбарно-таламического тракта аксоны tr. vestibulothalamicus проходят до таламуса, заканчиваясь на его центральных ядрах (3 нейроны) ⁃ аксоны центральных ядер таламуса направляются через заднюю ножку капсулы в корковую часть вестибулярного анализатора - кора и нижняя височная извилина
71.Вестибуломоторные, вестибулосенсорные и вестибуловегетативные реакции. Вестибуломоторные реакции (головокружение): 1. Нистагм глаз 2. Изменения позы за счет перераспределения мышечного тонуса 3. Нарушения координации движения 4. Нарушения походки Вестибуловегетативные реакции (тошнота): 1. Изменение ширины зрачка 2. Изменение цвета кожи лица (сосудистая реакция) 3. Тахикардия 4. Изменение тонуса, моторики ЖКТ Вестибулосенсорные реакции (подкорковые ядра) 1. Ощущения головы и тела в пространстве Вестибуло-моторные рефлексы: нистагм глаз, изменение позы за счет перераспределения мышечного тонуса, нарушение координации движений (при пальценосовой пробе, пробе с почерком), нарушение походки. Вестибулосенсорные реакции относятся к особому виду ощущений, имеющему специфическое качество. С помощью этого ощущения человек при нормальном состоянии вестибулярного анализатора безошибочно определяет состояние своего пространственного положения. От действия вестибулярного стимула не существует «защиты», поскольку его адекватные раздражители (сила земного притяжения и инерции) являются физическим свойством, действующим на вездесущий феномен массы. Открытость вестибулярного аппарата своему адекватному стимулу — чрезвычайно важная биологическая необходимость, обеспечивающая организм информацией о положении тела в пространстве и его движении для принятия оперативных (безусловнорефлекторных) действий, сохраняющих или возвращающих телу биологически адекватное положение в пространстве. Вестибуловегетативные реакции в своем физиологическом выражении являются эволюционно обусловленными механизмами адаптации организма к его возрастающим энергетическим затратам, необходимым при действии знакопеременных ускорений. Эта адаптация определяется повышением трофической функции симпатической нервной системы. При пассивном восприятии этих ускорений (укачивание) у особо чувствительных лиц развивается болезнь движения.
72.Физиология вкусового анализатора. Вкусовой анализатор обеспечивает восприятие и анализ химических веществ, поступающих в полость рта и формирование вкусовых ощущений. Главное назначение этого анализатора заключается в оценке вкусовых свойств пищи и ее пригодности к употреблению. Рецепторный орган вкуса – вкусовые почки (вкусовые рюмки, вкусовые луковицы), открытые в 1868 году К. Ловеном и Г. Швальбе. Вкусовые почки сосредоточены в основном в языке, но также в слизистой мягкого неба, глотки, надгортанника и верхней части пищевода, где расположены одиночные вкусовые клетки. Вкусовые почки, расположенные в языке, связаны с тремя типами сосочков (грибовидные, листовидные и желобовидные). Вкусовые рецепторы разделяют в зависимости от вида ощущения, возникающего при их раздражении. Среди многообразия смешанных вкусов, которые различает человек, выделяют четыре элементарные: сладкий, кислый, соленый, горький. Вкусовые почки иннервированы: языкоглоточным нервом (задняя треть языка), ветвью лицевого нерва – барабанная струна (передние две трети языка) и верхнегортанным нервом – частью блуждающего нерва (области глотки, надгортанника и гортани). По данным афферентным нервам возбуждение поступает в ядро одиночного пути продолговатого мозга, оттуда к таламусу и далее пути направляются во вкусовую зону коры головного мозга, которая расположена в постцентральной извилине рядом с зоной тактильной чувствительности языка (рис. 3). Определенная часть вкусовых путей заканчивается в гиппокампе, височной области коры и покрышечной части лобной доли (pars opercularis — operculum Rolandi). Анализатор вкуса - вкусовой анализатор выполняет следующие функции в деятельности пищеварительной системы: a) представляет информацию о свойствах и качестве пищи; б) располагаясь в начальном отделе пищеварительной системы, рефлекторно воздействует на железы (слюнные железы, железы желудочно-кишечного тракта, печень, поджелудочную железу) и, тем самым, оказывает на них регулирующее влияние; b) предоставляя информацию о вкусовых раздражениях обеспечивает адекватную ответную реакцию на вкусовые раздражения (тонус мышц, эмоции, настроение и т.д.) - вкусовые рецепторы находятся в полости рта и представлены вкусовыми клетками, которые входят в состав вкусовых почек - луковиц - вкусовые луковицы располагаются большей частью на языке в области грибовидных, желобоватых и листовидных сосочков; меньшее их количество находится в эпителии слизистой оболочки полости рта, губ, мягкого неба, небныхдужек, глотки, надгортанника; - совокупность вкусовых почек в полости рта составляет орган вкуса; - вкусовая почка в центре имеет ямку, в которую попадают растворенные в слюне вещества; - в ямку обращены вкусовые (рецепторные) клетки; - вкусовые луковицы функционально специализированы: a) сладкое воспринимают вкусовые луковицы кончика языка; б) кислое - луковицы боковой поверхности языка; b) горькое - луковицы корня языка; г) соленое - луковицы всей поверхности языка; - вкусовые клетки трансформируеют химическое раздражение в нервный импульс; - от вкусовых клеток импульсы поступают к чувствительным ганглиям черепных нервов (1 нейроны): a) к ganglion geniculi лицевого нерва - от передних 2/3 языка; б) к ganglion inferius языкоглоточного нерва - от задней 1/3 языка; b) к ganglion inferius блуждающего нерва - от вкусовых рецепторов глотки, надгортанника, неба - от указанных чувствительных ганглиев импульсы следуют к nucleus tractus solitarii (2 нейроны); - большая часть аксонов клеток ядра одиночного пути переходит на противоположную сторону и в составе tr. nucleothalamicus следует к базальным ядрам таламуса (3 нейроны); - аксоны третьих нейронов в составе tr. thalamocorticalis направляются в крючок и парагиппокампальную извилину - проекционный центр вкуса; Часть аксонов базальных ядер таламуса направляются в медиальные ядра таламуса – чувствительный интеграционный центр экстрапирамидной систему, который в ответ на вкусовые раздражение обеспечивает изменение тонуса мышц
73. Физиология обонятельного анализатора. Обонятельная сенсорная система обеспечивает восприятие и анализ информации о пахучих химических веществах (одорантах), соприкасающихся со слизистой оболочкой носовой полости и формирует обонятельные ощущения. Деятельность обонятельного анализатора направлена на оценку качества внешней среды (например, воздуха, пищи), участие в регуляции системы пищеварения, коммуникационных функций, эмоциональных состояний. Хеннинг (1924) предложил графическое изображение всех обонятельных ощущений в виде призмы, в углах которой располагаются шесть первичных запахов (цветочный, гнилостный, фруктовый, прогорклый-запах горнлого, пряный и смолистый). Другие запахи должны размещаться на ребрах, гранях или внутри призмы, в зависимости от того, с какими классами у них обнаруживается сходство. Рецепторный орган обоняния – обонятельная выстилка, расположенная в верхней и средней раковинах носовой полости и перегородке носа. Обонятельную выстилку образуют три типа клеток: рецепторные и вспомогательные: опорные и базальные. рецепторами обоняния являются разветвления периферических отростков биполярных клеток (первые нейроны) слизистой оболочки полости носа (рис. 36) в regio olfactoria (область верхнего носового хода, верхней носовой раковины и верхней части перегородки носа); - периферические отростки биполярных клеток (обонятельные луковицы полости носа) заканчиваются булавовидными рецепторами, на каждом из которых находятся 10-15 обонятельных волосков, погруженных в слой слизи; - слизь вырабатывают обонятельные железы полости носа, окружающие обонятельные луковицы; - обонятельная область слизистой оболочки носа и обонятельные железы в совокупности составляют орган обоняния; пахучие вещества, проникающие с потоком воздуха в полость носа, растворяются в слизи, обонятельные волоски взаимодействуют с молекулами пахучих веществ и трансформируют энергию химического раздражения в нервные импульсы; - центральные отростки рецепторных клеток образуют пn. olfactorii, которые через lamina cribrosa проникают в полость черепа к митральным клеткам bulbusolfactorius(2 нейроны); -аксоны митральных клеток проходят в составе tr.olfactorius и в близи обонятельного треугольника распадаются на три пучка: а)волокна медиального пучка направляются через переднюю спайку мозга в обонятельный тракт противоположной стороны к митральным клеткам контралатеральной обонятельной луковицы ; б) волокна промежуточного пучка направляются к нейронам обонятельного треугольника, переднего продырявленного вещества и нейронам ядер прозрачной перегородки своей и, частично, противоположной сторон; b) волокна латерального пучка направляются к крючку и парагиппокампальной извилине - в проекционный центр обоняния; - важной особенностью обонятельного пути является то, что нервные импульсы первоначально поступают в кору полушарий большого мозга (gyrus parahippocаmpalis et uncus), и только затем - в подкорковые центры обоняния; Подкорковые центры - сосочковые тела и передние ядра таламуса
74. Физиология интероцептивного анализатора. Интероцептивный (висцеральный) анализатор обеспечивает восприятие и анализ информации о функционировании внутренних органов, тканей и составе жидких сред организма. Интероцептивный анализатор в норме не вызывает отчетливых ощущений, за редким исключением, например, при наполнении полых органов (желудок, прямая кишка, мочевой пузырь). При определенных состояниях висцеральной сферы возникают неосознаваемые ощущения, названные И.М. Сеченовым: «темные» (смутные) чувства. Понятие интероцептивной сенсорной системы объединяет тканевые рецепторы и рецепторы слизистых оболочек (В. Н. Черниговский, 1974), к которым принадлежат механо-, хемо-, термо- и осморецепторные аппараты. Интероцепторы могут быть представлены свободными нервными окончаниями (например, дендриты нейронов спинальных ганглиев), инкапсулированными нервными окончаниями (тельца Фатера-Пачини, колбы Краузе), специализированными клетками (рецепторы каротидного и аортального клубочков). Анализаторы: 1. Химизма внутренней среды 2. Осмотической среды 3. Давления в сосудах 4. Температуры внутренней среды 5. Давления во внутренних полых органах Периферический отдел: Механо, термо, хемо, осмо, баро на органах, сосудах – все первичные Хеморецепторы – в аортальном и каротидном клубочках, слизистые пищеварительной системы, органов дыхания Механорецепторы – сосуды, сердце, почки итд Осморецепторы – аортальный, каротидный синус, печень и др Терморецепторы – сл оболочки пищевара, органов дыхания, мочевого пузыря, серозные оболочки, каротидный синус, гипоталамус Проводниковый отдел: Афферентные волокна вегетативных нервов: 1. Нейрон – спинальный ганглий или черепной ганглий 2. Нейрон – спинной или продолговатый мозг 3. Нейрон – задне - медиальные ядра таламуса Корковый отдел: 4 Нейрон – соматосенсорные области коры (постцентральная извилина) – обеспечивает формирование натуральных вегетативных УР, например, пища в одно и то же время
75. Физиология кожного анализатора. Кожный анализатор представляет совокупность афферентных систем, обеспечивающих несколько видов кожной чувствительности: температурную, тактильную, болевую. Тактильная чувствительность. Ей принадлежит ведущая роль в формировании полного представления о внешнем мире (желание потрогать для достоверности). Рецепторы расположены на всей поверхности кожи. Эта модальность чувствительности объединяет четыре качества ощущения: давления, прикосновения, вибрации и щекотания. Тактильную чувствительность обеспечивают рецепторы различного вида (рис. 19) и строения: тельце Мейснера, диски Меркеля, тельце Пачини, рецептор волосяного фоликула, тактильный диск, окончание Руффини.

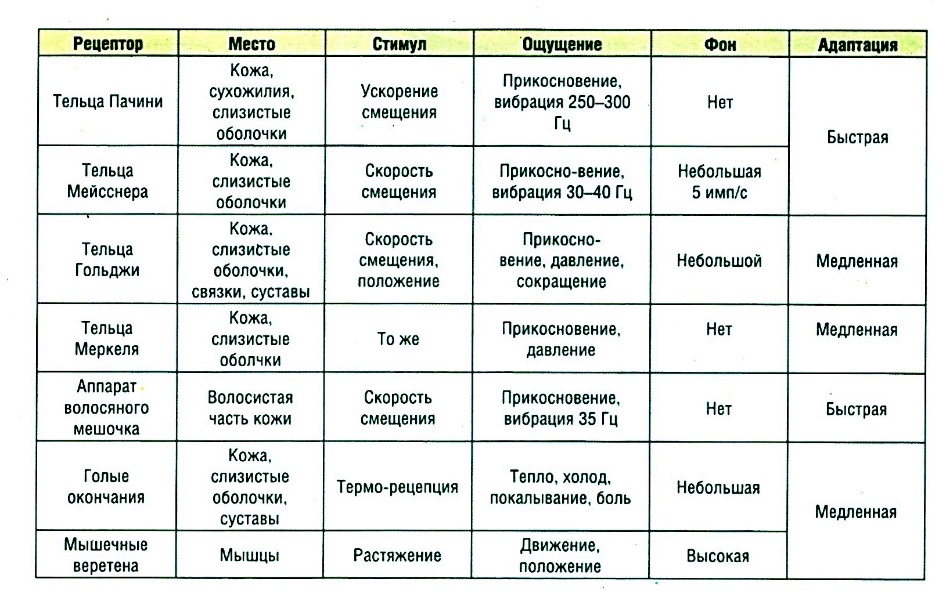
Проводящие пути кожного анализатора. Афферентные волокна, связанные с различными видами кожной чувствительности, отличаются друг от друга толщиной миелиновой оболочки, следовательно, и скоростью проведения импульсов. По волокнам безмякотным и обладающим тонкой миелиновой оболочкой поступают импульсы, связанные с температурной, болевой, а отчасти и тактильной чувствительностью. Волокна второго нейрона проводящих путей перекрещиваются, и направляются к ядрам бугров промежуточного мозга, где находятся тела третьего нейрона, проводящего импульсы к коре больших полушарий. Через боковые ответвления (коллатерали) нервных волокон и через промежуточные (вставочные) нейроны импульсы, идущие от кожных рецепторов, могут переходить в спинном мозге на эфферентные пути. Часть волокон направляется к бугоркам четверохолмия, при участии которых осуществляются такие рефлекторные реакции на раздражение кожи, как поворот головы, движение глаз, настораживание. Корковый отдел кожного анализатора. У человека ядро коркового отдела кожного анализатора расположено в постцентральной области коры больших полушарий. Основная масса афферентных волокон направляется к полю 5, которое находится в глубине центральной борозды, а также к соседним полям 1 и 2. Уже в спинном мозге пучки волокон, несущих импульсы от разных участков кожи, занимают строго определенное положение, и, дойдя до латерального ядра бугров промежуточного мозга, заканчиваются около различных клеточных групп, каждая из которых направляет свои аксоны к определенным участкам коры, преимущественно противоположного полушария. В результате ядро коркового отдела кожного анализатора можно рассматривать как своеобразную проекцию кожи. Исследование мозга людей, у которых при жизни отсутствовала чувствительность отдельных участков кожи, а также опыты с электрическим раздражением коры во время мозговых операций позволили установить корковую локализацию кожной чувствительности у человека (рис. 22). В верхнем участке постцентральной извилины проецируется кожа нижних конечностей, а в среднем и нижнем участках - кожа верхних конечностей и головы. Величина корковой проекции различных областей кожи пропорциональна их богатству рецепторами, что совпадает с функциональной значимостью отдельных участков кожного анализатора. Так, проекция кожи туловища занимает относительно малое пространство.
76. Физиология проприоцептивного (двигательного) анализатора. Двигательный анализатор (син.: кинестетический анализатор, проприоцептивный анализатор, мышечный анализатор) - физиологическая система, передающая и обрабатывающая информацию от рецепторов скелетно-мышечного аппарата (мышечных, сухожильных, суставных и костных) и участвующая в организации и осуществлении координированных движений. Двигательный анализатор, как и любой другой анализатор, состоит из трех основных отделов: воспринимающего, проводникового и коркового, тесно связанных между собой сложной системой восходящих и нисходящих взаимодействий. Воспринимающая часть представлена рецепторами со вспомогательными аппаратами. Скелетные мышцы содержат несколько видов механорецепторов. Лучше других изучены рецепторы растяжения, к которым относятся мышечные веретена и сухожильные органы Гольджи (рис. 2). Они необходимы для восприятия позы и движений тела, играют важную роль в регуляции двигательной активности. Кроме механорецепторов, в мышцах находятся болевые рецепторы, которые реагируют на сильное давление, оказанное на мышцы, и на изменение содержания в них метаболитов, особенно при ишемии (нарушении кровоснабжения) Вспомогательный аппарат рецепторных образований представлен интрафузальными мышечными волокнами мышечных веретен (рис. 3), внеклеточным матриксом, прикрепляющим элементы аппарата Гольджи к сухожилиям, а также капсулами телец Фатер — Пачини и Руффини. Все эти структуры улучшают восприятие действующих раздражителей, вызывающих укорочение, напряжение и растяжение мышцы, а также растяжение сухожилий и суставных сумок. Проводниковая часть двигательного анализатора начинается афферентными волокнами от различных рецепторов: от мышечных веретен — афференты групп Ia и II (рис. 4), от аппарата Гольджи — афференты группы Ib, от ноцицепторов — афференты группы III или IV, от телец Фатера — Пачини и Руффини — афференты группы II. Центральную (корковую) часть двигательного анализатора образуют нейроны, расположенные позади сильвиевой борозды и перед ней. Задняя центральная извилина образует сенсомоторную область (рис. 6), а передняя центральная извилина — моторную область.
77. Гормональная (эндокринная) регуляция: звенья гормональной системы. Функциональная система эндокринной регуляции функций включает несколько звеньев. 1. Звено управления представлено структурами центральной нервной системы и вегетативной нервной системой, гипоталамо-гипофизарным комплексом, церебро-питуитарным и церебро-гландулярным путями управления. 2. Звено синтеза гормонов определяется цитологическими и метаболическими характеристиками эндокринных клеток. 3. Звено секреции гормонов определяется процессами накопления гормонов или прогормонов, химического связывания и расщепления прогормонов, их внутриклеточным транспортом. 4. Звено транспорта гормонов обеспечивается кровью в виде свободных и связанных с белками форм. Биологическая активность обычно определяется содержанием свободных форм гормонов. Связи гормонов с белками являются физико-химическим процессом и не зависит от расхода энергии. Гормоны могут образовывать несколько транспортных связей с белками при увеличении их уровня в крови. 5. Звено депонирования локализовано в самих эндокринных клетках, например, катехоламины накапливаются в хромаффинных клетках мозгового вещества надпочечников. Однако не все эндокринные клетки способны депонировать готовые гормоны. Например, клетки коры надпочечников не могут депонировать гормоны и объём их секреции прямо зависит от объёма синтеза. Гормоны могут накапливаться в органах-мишенях, например, катехоламины в миокарде. 6. Звено метаболизма осуществляется в органах под влиянием имеющихся ферментов. Метаболические превращения гормонов протекают в печени, почках, органах-мишенях, самих эндокринных тканях. При этом образуются либо продукты деградации гормонов, не имеющие биологической активности, либо метаболиты с более высокой или более низкой активностью. 7. Звено экскреции обеспечивает поддержание концентрации гормонов на требуемом уровне. Выведение гормонов осуществляется преимущественно почками путем клубочковой фильтрации и канальцевой секреции, а также печенью с желчью, желудком, кишечником, слюнными железами, в составе пота и кожного сала. Вместе со звеньями депонирования и метаболизма выведение гормона обеспечивает его удаление из крови. 8. Звено эффектора реализует биологический эффект гормонов на клетки, ткани, органы и физиологические системы благодаря наличию на клеточных мембранах и цитоплазме специфических гормональных рецепторов. Количество рецепторов и их сродство к гормону определяет интенсивность ответа эффектора. Ткани и органы, клетки которых имеют рецепторы к гормону называются "мишенями". 9. Звено обратной связи управления. Различают обратные связи от эффекта и от концентрации гормона в крови. Примерами первого типа обратных связей являются: регуляция уровня сахара в крови продукцией инсулина и глюкагона, регуляция уровня кальция в крови продукцией паратгормона и кальцитонина. Пример второго типа обратной связи: если щитовидная железа секретирует избыточное количество тироксина, эта повышенная концентрация гормона подавляет продукцию тиролиберина или тиротропина или синтез самого тироксина, в результате чего секреция и уровень тироксина в крови снижается.
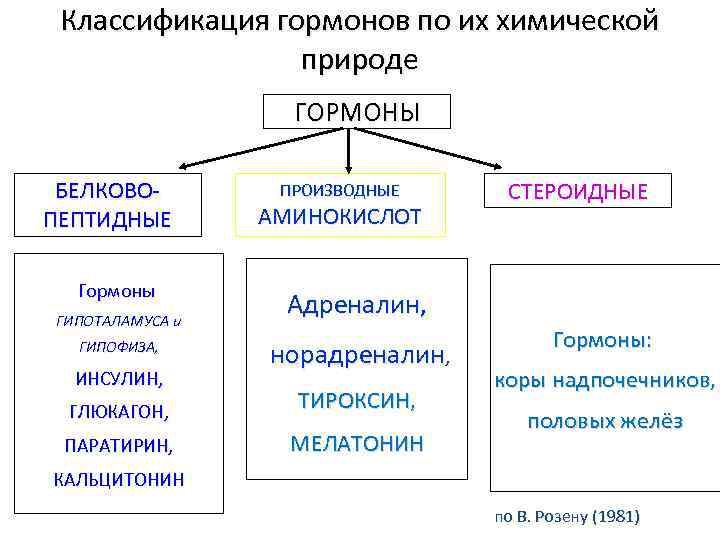
78. Классификация гормонов по химической природе, источнику образования По химической природе: 1. Белки и полипептиды: простые одноцепочечные пептиды с различным количеством аминокислотных остатков инсулин, глюкагон, кальцитонин, паратгормон, АДГ, окситоцин, тканевые гормоны – гастрин, секретин, ангиотензин, холицистокинин; тропные гормоны таламуса, релизинг-гормоны гипоталамуса, гипофиз: фолликулостимулирующий и тиреотропный2. Стероидные, производные холестерина (Тестостерон, эстрадиол, эстрон, прогестерон, кортизол, альдостерон) 3. Тиреоидные (тироксин, трийодтиранин) 4. Катехоламины (адреналин, норадреналин, дофамин) 5. Эйказаноиды - производные арахидоновой кислоты (простагландины, тромбоксаны, лейкотриены – местная регуляция)

79. Механизмы действия и эффекты гормонов. Принципы регуляции выработки гормонов Эффекты гормонов: 1. Пусковой (запускает деятельность эффектора АДГ на нефрон) 2. Пермиссивный – способность одного гормона обеспечивать реализацию эффекта другого гормона (Инсулин необходим для СТГ, пролактин – для эстрогена и прогестерона) 3. Модулирующий – изменение интенсивность деятельности органа (адреналин на сердце) Механизм: 1911, 1913 – П. Эрлих «замок-ключ» - для каждого гормона в клетках-мишенях есть специальный рецептор. Два основных варианта действия гормона на клетку: 1. Если клеточная мембрана непроницаема для гормона, то рецептор находится в мембране (для белково-пептидных, катехоламинов – нерастворимы в липидах). Образуются вторичные посредники, которые реализуют действие через ядерный аппарат: a. Аденилатциклаза – цАМФ b. Гуанилатциклаза – цГМФ c. Фосфолипаза С – ДАГ и Инозитолтрифосфат d. Ионизированный Кальций – кальмодулин 2. Если проницаемость для гормона нормальная (стероидные, растворимые в липидах) – взаимодействуют с внутриклеточными рецепторами, они связываются с последовательностями ДНК или РНК. Взаимодействие с Джи-белком – метаболические процессы.
80. Тиреоидные гормоны: физиологическая характеристика (групповая принадлежность по химическому строению, механизм действия, «структуры-мишени», физиологические эффекты, регуляция секреции). Изменения в организме при гиперсекреции и гипосекреции. Щитовидная железа (ЩЖ) – эндокринная железа, депонирующая йод и вырабатывающая йодосодержащие гормоны (йодтиронины), участвующие в регуляции обмена веществ и росте отдельных клеток и организма в целом. По хим природе: производные аминокислоты тирозина Механизм действия ТТГ на щитовидную железу. Гормон-рецепторный комплекс (ТТГ+Р) на мембране тироцита приводит к активации Na+/Н+-обменника, в результате происходит рН-зависимая активация ферментов синтеза белков коллоида и тироглобулина. Сопряжённо с переносом Na+ происходит перенос тирозина и Сl - в обмен на НСО-3. ТТГ активирует NIS (натрий-йодидный симпортер, 2Na+1/J+), обеспечивающий перенос одного иона йода в направлении более высокого электрохимического потенциала, а источником энергии служит Na+/K+-АТФаза в мембране тироцита. Таким образом, внутриклеточная концентрация Na+ восстанавливается, а клетка получает субстраты для синтеза гормонов. Йодид перемещается по цитоплазме от базального полюса к апикальному, где йодид/хлорный (J-/Cl-) обменник – белок пендрин переносит его в полость фолликула. Йодирование тирозина происходит до моно- (Т1) и дийодтирозана (Т2). Т1 и Т2 акцептируется на поверхности тироглобулина. Так как в клетку переносятся осмо- и гидрофильные ионы, в клетку поступает вода. Это стимулирует апокриновую секрецию коллоида в полость фолликула. Молекула тироглобулина свёртывается в гломерулу и молекулы Т1 и Т2 взаимодействуют друг с другом с образованием трийодтиронина (Т3) и тетрайодтиронина (Т4, тироксина). Механизмом пиноцитоза молекула тироглобулина возвращается в клетку. Пиноцитоз сопровождается закислением, это приводит к активации эстеразных свойств тироглобулина и отщеплению Т3, Т4 от носителя. Везикулярный экзоцитоз Т3, Т4 в кровь регулируется ТТГ.
Клетки-мишени: гепатоциты печени, адипоциты жировой ткани, клетки мышечной ткани, в том числе миокарда Основной механизм регуляции – гуморальный (рис.18). В гипофизотропной зоне гипоталамуса вырабатывается ТРГ – либерин (трипептид), который через воротные сосуды попадает в аденогипофиз и стимулирует выработку ТТГ (гликопротеин), который действует все этапы секреторного цикла и увеличивает секрецию Т3, Т4 в кровь. После гипофизэктомии железа захватывает йод очень мало, синтез тироглобулина, Т3, Т4 значительно снижен. Два продукта трийодтиронин и тетрайодтиронин поступают в кровь. Транспорт Т3 и Т4 кровью осуществляется белками-переносчиками: 60% α-глобулином (тироксин связывающий глобулин, ТСГ), преальбумином около 30% и 10% альбумином (тироксин связывающий альбумин, ТСА). Определяет гормональный статус концентрация гормона, не связанного с белками – это 0,1% свободного гормона. Концентрация свободного гормона находится в подвижном равновесии со связанным гормоном. Период полувыведения Т4 из плазмы составляет около 7 дней, Т3 от 8 до 10 часов. Эффекты: 1. Повышение потребления кислорода организмом (калоригенный эффект) 2. Рассопрягающее действие – разобщение окисления и фосфорилирования в процессах клеточного дыхания 3. Усиление энерготрат организма при активации всех биохимических процессов – увеличение размеров митохондрий, повышение их энергизованности, масса тела уменьшается при избытке тиреоидных гормонов 4. Усиление действия адреналина на гликогеноиз и глюконеогенез 5. Ускорение всасывания глюкозы в кишечнике, усиление ее захвата жировой и мышечной тканями 6. Усиление липолиза Гипотиреоз – заболевание, обусловленное снижением функции щитовидной железы и недостаточностью выработки ею гормонов. Проявляется замедлением всех процессов, происходящих в организме: слабостью, сонливостью, увеличением веса, медлительностью мышления и речи, зябкостью, гипотонией, у женщин - нарушениями менструального цикла. В тяжелых формах развивается микседема у взрослых и кретинизм (слабоумие) у детей. Кретинизм - дефицит Миксидема - дефицит (отёк подкожной клетчатки) Базедова болезнь

81. Физиологическое значение кальция в организме. Регуляция кальциевого обмена. Кальций организме выполняет важную регуляторную и структурную функции. Кальций участвует в таких процессах, как проведение нервного импульса, регулирует уровень возбудимости (инактивируя натриевые каналы, определяет уровень возбудимости), является потенциалобразующим ионом в кардиомиоцитах, необходим для синаптической передачи, обеспечивает сокращение мышечных волокон, является одним из факторов свёртывания крови, выполняет функцию вторичного посредника, является ко-фактором для многих ферментов, регулирует проницаемость клеточных мембран, принимает участие в оплодотворении, является основой костного скелета. Кальция в нашем организме довольно много – около 1200г Кальциемия регулируется тремя основными гормонами: тирокальцитонином (ТКТ), паратирином (паратиреоидным, ПТГ) и кальцитриолом – активным метаболитом витамина D3. Главный кальцийрегулирующий гормон – паратгормон (ПТГ) вырабатывается паращитовидными железами. В норме их четыре, реже больше или меньше. Они расположены на поверхности щитовидной железы. Иногда одна из желёз расположена атипично – в средостении, на дуге аорты. Основным регулятором секреции паратгормона является незначительное снижение содержания ионизированного кальция в плазме крови (рис.12). Рецептор Са2+ (Са2+-сенсор) относится к вмонтированным в плазмолемму главных клеток гликопротеинам, связанным с G-белком. Такие сенсоры присутствуют в клеточных мембранах паращитовидных желёз, К-клеток щитовидной железы, почек, кишечника, в некоторых структурах головного мозга. В почке паратгормон регулирует продукцию кальцитриола. Образовавшийся кальцитриол ингибирует синтез паратгормона по механизму отрицательной обратной связи. Органы-мишени ПТГ – кости, почки и желудочно-кишечный тракт (преимущественно тонкая кишка).
82. Катехоламины: физиологическая характеристика (групповая принадлежность по химическому строению, механизм действия, «структуры-мишени», физиологические эффекты, регуляция секреции) По хим строению: производные дифенола-катехола, мозговое вещество надпочечников Органы мишени - печень, мышцы Катехоламины – ФАВ, химический посредник, медиатор в межклеточном взаимодействии – срочное приспособление к действию сверхпорогового раздражителя при стрессовых ситуациях – мобилизуют глюкозу и ЖК для использования их в качестве источников энергии, подготавливают сердце и мышцы к выполнению работы (ПРОТИВОПОЛОЖНЫ ГЛЮКОКОРТИКОИДАМ) (А, Н, Д) Эффекты адреналина: 1. ЧСС + 2. Сужение сосудов 3. Теплообразование 4. Перистальтика ЖКТ – 5. Мускулатура бронхов – 6. Ренин + 7. Моча – 8. Возбудимость нервной ткани + Метаболические эффекты адреналина: 1. Повышение содержания глюкозы и ЖК в крови, активация гликогенолиза 2. Стимуляция липолиза (активация липазы) 3. Активация кетогенеза в печени 4. Усиление использования ЖК и ацетоуксусной кислоты в качестве энергии в миокарде и корковом слое почки 5. Усиление окислительных процессов в тканях 6. Значительное потребление организмом кислорода 7. Повышение Т тела (усиливаются калоригенные процессы, скорость метаболизма повышается на 20-30%) Метаболические эффекты: Эффекты норадренолина – мозговое вещество надпочечников, медиатор постганглионарных симпатических окончаний, медиатор ВНС, ЧСС (-) Дофамин – медиатор ЦНС, мозговое вещество надпочечников – тормозный медиатор в сонных гломусах, симпатических ганглиях, обеспечивает когнитивные процессы Тирозин – дофа – дофамин – норадреналин – адреналин Стресс – гипоталамус – АКТГ-релизинг-фактор – АКТГ – мозговое вещество надпочечников Регуляция секреции гормонов осуществляется благодаря наличию гипоталамо-симпатоадреналовой оси. Иннервируются длинными преганглионарными миелинизированными холинергическими волокнами преимущественно чревного нерва. Этот нерв регулирует секрецию и выделение адреналина надпочечниками. Перерезка нерва вызвало резкое уменьшение секреции, а его раздражение вело к усиленному отделению адреналина. В Регуляции деятельности мозгового вещества надпочечников высшая регуляторная роль принадлежит коре головного мозга и это влияние коры через подкорковые центры и симпатический отдел нервной системы передается хромофинным клеткам мозгового вещества. Мозговой вещество надпочечников и симпатический отдел ВНС - Примеры взаимодействия нервной и эндокринной систем. Их объединили в симпатоадреналовой систему. Они обеспечивают адаптацию организма к острым и хроническим стрессом, являются основными элементами реакции борьбы и бегства.
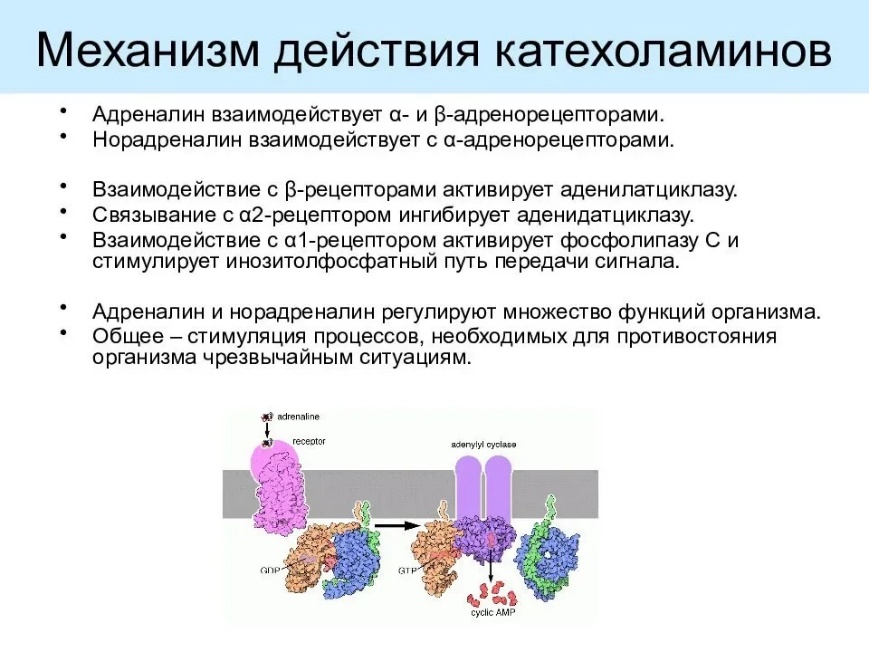
83.Глюкокортикоиды: физиологическая характеристика (групповая принадлежность по химическому строению, механизм действия, «структуры-мишени», физиологические эффекты, регуляция секреции). Понятие об общем адаптационном синдроме. Глюкокортикоиды – гормоны коркового слоя надпочечников (пучковая зона коры) 95% КОРТИЗОЛ Структуры-мишени: соединительная ткань, • мышцы, • жировая ткань, • лимфоидная ткань, • печень, • почки. По хим строению: кора надпочечников, стероидные гормоны Пучковая и сетчатая зоны коркового вещества надпочечников является гипофиз-зависимой частью железы. Секреция гормонов связана между собой классическими петлями прямых и обратных связей (рис.14). Кортиколиберин (АКТГ-либерин) гипофизотропной зоны гипоталамуса поступает в гипофиз и вызывает выделение АКТГ (адренокортикотропного гормона). АКТГ доставляется кровью к коре надпочечника и стимулирует синтез и секрецию глюкокортикоидов. Глюкокортикоиды по механизму отрицательной обратной связи подавляют его образование. Секреция кортизола эпизодична (изменяется в соответствие с циркадными биоритмами) Эффекты: 1. Иммунный - подавляет синтез антител, тормозит взаимодействие антител с антигенами, ослабляет фагоцитоз 2. В белковом обмене – катаболизм, повышение образования мочевины 3. Жировой обмен – липолиз, кетогенез 4. Противовоспалительный – стабилизация мембран лизосом, уменьшение секреции лейкоцитами интерлейкина-1, уменьшение проницаемости капилляров, уменьшение экссудации и отечности тканей 5. Противоаллергический – антигистаминное действие и ингибирование иммуноглобулина Е 6. ССС – уменьшение проницаемости стенок капилляров, нарушение возбудимости и проводимости миокарда (повышение чувствительности адренорецепторов к катехоламинам) 7. Гематологический – уменьшение содержания лимфоцитов, моноцитов, эозинофилов вследствие их перехода из крови в другие ткани 8. Адаптивный – накопление глюкозы для мозга, поддержание гомеостазиса 9. Подавление ИС – уменьшение клеточного и гуморального иммунитета 10. Неврологический –повышение сенсорной чувствительности (обоняние, вкус, слух) и возбудимость НС Гипоталамус – кортикотропный релизинг–фактор – передняя доля гипофиза – АКТГ – кора надпочечников – гидрокортизон, кортизол


1.Стадия тревоги (алярм-реакция) характеризуются мобилизацией липидов из клеток для синтеза глюкокортикоидов, выброс адреналина 2. Стадия резистентности (адаптации) сопровождается гипертрофией пучковой зоны, накоплением липидов в клетках, усиленным выделением глюкокортикоидов, гиперемия кровеносных сосудов, повышение АД, выброс гистамина 3. Стадия истощения развивается при тяжелых формах патологии и чаще в их терминальных фазах, характеризуется гипоплазией и гипофункцией пучковой зоны.
84.Минералокортикоиды: физиологическая характеристика (групповая принадлежность по химическому строению, механизм действия, «структуры-мишени», физиологические эффекты, регуляция секреции). Синтезируются в клубочковой зоне коры надпочечников, основной гормон – альдостерон – провоспалительный гормон (стимуляция воспаления), стероидные В то же время мишенями аналогичного действия минералокортикоидов являются слизистые оболочки желудочно-кишечного тракта, потовые, слюнные и молочные железы. Альдостерон является частью ренин-ангиотензин-альдостероновой системы (РААС). Основной регулятор секреции этого гормона ангиотензин II (АТ-II). АКГТ в регуляции синтеза альдостерона оказывает пермиссивное действие, также обеспечивает накопление холестерола. Механизм действия альдостерона связан с его геномными и негеномными эффектами. Основные клетки- и органы-мишени – это почки, слюнные и потовые железы, энтероциты толстой кишки, гладкомышечные волокна артериол, артерий и в меньшей степени вен. Суммарно все эффекты МК направлены на сохранение натрия, выведение калия, поддержание рН крови, регуляцию артериального давления и поддержания объёма циркулирующей крови (ОЦК). Функция – регуляция водно-солевого обмена за счет реабсорбции ионов Натрия в дистальных канальцах почки (Калий-Натриевая АТФ-аза), адаптация и акклиматизация к жаре

85. Ренин-ангиотензин-альдестероновая система. Ренин-ангиотензин-альдостероновая система (РААС) выполняет важное гуморальное влияние на сердечно-сосудистую систему и участвует в регуляции артериального давления. Центральным звеном РААС является ангиотензин II (АТ II) (схема 1), который обладает мощным прямым вазоконстрикторным действием преимущественно на артерии и опосредованным действием на ЦНС, высвобождением катехоламинов из надпочечников и вызывает увеличение ОПСС, стимулирует секрецию альдостерона и приводит к задержке жидкости и повышению (ОЦК), стимулирует выброс катехоламинов (норадренолина) и других нейрогормонов из симпатических окончаний. Влияние АТ II на уровень АД осуществляется за счет действия на тонус сосудов, а также посредством структурной перестройки и ремоделирования сердца и сосудов. В частности, ATII является также фактором роста (или модулятором роста) для кардиомиоцитов и гладкомышечных клеток сосудов. Ангиотензин I малозначим в системе РААС, так как быстро превращается в АТП, кроме того, его активность в 100 раз меньше активности АТП. Ангиотензин III действует подобно АТП, но его прессорная активность в 4 раза слабее АТП. Ангиотензин 1-7 образуется вследствие превращения ангиотензина I. По функциям он значительно отличается от АТП: он не вызывает прессорного действия, а наоборот, приводит к снижению АД благодаря секреции АДГ, стимуляции синтеза простагландинов, натрийуреза. РААС оказывает регулирующее влияние на почечную функцию. АТП вызывает мощный спазм приносящей артериолы и снижение давления в капиллярах клубочка, уменьшение фильтрации в нефроне. В результате снижения фильтрации уменьшается реабсорбция натрия в проксимальном отделе нефрона, что приводит к увеличению концентрации натрия в дистальных канальцах и активации Na-чувствительных рецепторов плотного пятна в нефроне. По механизму обратной связи это сопровождается торможением выделения ренина и увеличением скорости клубочковой фильтрации. Функционирование РААС связано с альдостероном и посредством механизма обратной связи. Альдостерон - важнейший регулятор объема внеклеточной жидкости и гомеостаза калия. Прямого действия на секрецию ренина и АТП альдостерон не оказывает, но возможно косвенное влияние через задержку натрия в организме. В регуляции секреции альдостерона участвуют АТП и электролиты, причем АТП - стимулирует, а натрий и калий - уменьшают его образование. Ключевым ферментом РААС является ангиотензин-превращающий фермент (АПФ), он обеспечивает превращение ΑTI в ATII. Основное количество АПФ присутствует в системном кровотоке, обеспечивая образование циркулирующего АТII и краткосрочные геодинамические эффекты. Превращение АТ в ATII в тканях может осуществляться не только с помощью АПФ, но и другими ферментами (химазы, эндопероксиды, катепсин G и др.); считают, что им принадлежит ведущая роль в функционировании тканевых РАС и развитии длительных эффектов моделирования функции и структуры органов-мишеней.