

1. Нормальная физиология как наука: объект, предмет, цель, задачи, связь с медицинскими науками. Физиология – наука о механизмах функционирования и регуляции деятельности клеток, органов, систем и организма в целом и взаимодействии его с окружающей средой. Предмет изучения физиологии – функции живого организма, их связи между собой, регуляция и приспособление к внешней среде. (Лекция)Физиология (от греч. physis – природа и logos – уче ние) – наука о природе, о существе жизненных процессов. Это наука, изучающая процессы жизнедеятельности организма, составляющих его физиологических систем, отдельных органов, тканей, клеток и субклеточных структур, механизмы регуляции этих процессов, а так же действие факторов внешней среды на динамику жизненных процессов. ПРЕДМЕТ изучения нормальной физиологии – выяснение функций и механизмов (закономерностей деятельности органов и систем организма), обеспечивающих его целостность и приспособление (адаптацию) к изменяющимся условиям окружающей среды. Объект - живой организм
Цель физиологии: Установление факта, свидетельствующего о том, что происходит с той или иной функцией во время жизнедеятельности человека и животного, и выяснения, с помощью каких механизмов реализуется функция и с какой целью обеспечивается данная функция в определённой системе, органе, клетке.
ИЛИ глубокое познание механизмов и закономерностей осуществления процессов и функций, их регуляции. Основные задачи: -исследование механизмов функционирования клеток, тканей, органов, систем органов, организма в целом -изучение механизмов регуляции функций органов и систем органов -выявление реакций организма и его систем на изменение внешней и внутренней среды, а также исследование механизмов возникающих реакций -разработка методов изучения функций организма
Близко связана физиология и со всеми медицинскими дисциплинами: 1.разработка физиологии пищеварения И. П. Павловым дала возможность понять заболевания пищеварительного тракта и явилась основой диететики питания — мощного средства лечения; 2.изучение витаминов позволило успешно бороться с такими заболеваниями, как цинга и рахит;3. открытие гормона поджелудочной железы— инсулина и выяснение способов его получения способствовали сохранению жизни тысяч больных диабетом; 4.исследование групп крови явилось основой такого важного для медицинской практики мероприятия, как переливание крови.
2.
Структура физиологии. Методы физиологических
исследований.

 Методы
изучений:
1. Наблюдение – неудобный,
т.к. многие физ процессы очень быстры,
что приводит к ошибкам.
2. Графическая
регистрация физиологических процессов
– объективные записи изучаемого
процесса
3. Исследование биоэлектрических
явлений (Гальвани) (ЭКГ – Эйнтховен),
живые ткани – источники электрических
потенциалов, способных взаимодействовать
на нервы и мышцы другого организма
4.
Методы эл раздражения органов и тканей
– Дюба (сонный аппарат – индукционная
катушка для дозированного эл раздражения
живых тканей)
5. Острый эксперимент
– группа аналитических методов или
живосечение под наркозом, рассекают,
изучают деятельность органов (изолированное
сердце)
6. Хронический эксперимент
(Павлов) – изучение деятельности органа,
не нарушая целостность организма
(изучение путем внедрения фистул)
Методы
изучений:
1. Наблюдение – неудобный,
т.к. многие физ процессы очень быстры,
что приводит к ошибкам.
2. Графическая
регистрация физиологических процессов
– объективные записи изучаемого
процесса
3. Исследование биоэлектрических
явлений (Гальвани) (ЭКГ – Эйнтховен),
живые ткани – источники электрических
потенциалов, способных взаимодействовать
на нервы и мышцы другого организма
4.
Методы эл раздражения органов и тканей
– Дюба (сонный аппарат – индукционная
катушка для дозированного эл раздражения
живых тканей)
5. Острый эксперимент
– группа аналитических методов или
живосечение под наркозом, рассекают,
изучают деятельность органов (изолированное
сердце)
6. Хронический эксперимент
(Павлов) – изучение деятельности органа,
не нарушая целостность организма
(изучение путем внедрения фистул)
7. Метод экстирпации основан на удалении органа либо его части с последующим наблюдением и регистрацией последствий вмешательства с целью изучения функции и значения данного органа.
8.Метод трансплантации заключается в пересадке изучаемого органа на новое место в том же организме или переносе в другой организм.
3.Физиологическая школа Военно-медицинской академии. Вклад И.М, Сеченова, И.Р.Тарханова, И.П., Павлова, Л.А. Орбели в физиологию. Основателем самостоятельной кафедры физиологии в Академии является профессор А.П. Загорский. Он наладил систематическую демонстрацию экспериментов на лекциях, но не смог организовать на кафедре научную работу. Это удалось его преемникам: Н.М. Якубовичу и И.М. Сеченову. Первый из них был избран ординарным профессором в 1860 г., а И.М. Сеченов, избранный в том же году адъюнкт-профессором, в 1864 г. тоже стал ординарным профессором. С 1864 по 1868 г. на кафедре физиологии работали два ординарных профессора, чего не было ни на одной из академических кафедр. На этой кафедре И.М. Сеченов в 1860-е годы создал первую в России научную физиологическую школу, стал «отцом русской физиологии». С 10-летней работой в академии связаны все его выдающиеся научные достижения: открытие центрального торможения, обоснование рефлекторной природы психической деятельности человека, измерение среднего кровяного давления в артерии, вскрытие закономерностей растворения CO2 в крови и других жидкостях. Эти открытия послужили в дальнейшем базой авиационной и водолазной физиологии. Работы по газообмену и биоэнергетике легли в основу физиологии труда. Исследования в области нервно-мышечной физиологии и электрофизиологии сделали Сеченова родоначальником биофизики в России. Изученные им механизмы взаимодействия организма с внешней средой были реализованы в 1940-е годы при создании кибернетики. С 1872 по 1875 г. ординарным профессором физиологии работал И.Ф. Цион. Первый российский Нобелевский лауреат, академик АН СССР И.П. Павлов возглавлял кафедру с 1895 по 1925 г. Имел репутацию превосходного лектора. Поскольку каждая лекция сопровождалась демонстрациями экспериментов на животных, его лекционный курс неофициально называли «Физиологией в опытах». И.П. Павлов и его сотрудники проводили научные исследования в трех областях физиологии. При изучении кровообращения они уточнили важные вопросы его нервной регуляции. Физиологию пищеварения Павлов кардинально перестроил, за что был удостоен Нобелевской премии, а физиологию высшей нервной деятельности выстроил заново. До его работ такого раздела физиологии не существовало. И.П. Павлов настолько развил метод хронического эксперимента, что это позволило ему построить синтетическую физиологию. В 1905–1906 гг. кафедру переместили в новое здание, где она находится и поныне (ул. Комиссара Смирнова, 12а). Преемником И.П. Павлова стал его ученик, академик АН и АМН СССР Л.А. Орбели. При нем практические занятия стали не факультативными, а обязательными. Началось преподавание физиологии военного труда для врачей сухопутных войск, армии и флота, проходивших усовершенствование в академии. Три четверти кафедрального плана НИР занимала военно-медицинская тематика. Наряду с ней кафедральный коллектив продолжал разрабатывать фундаментальные проблемы физиологии. В 1939 г. во дворе кафедры построили специальное здание баротермолаборатории для решения актуальных задач авиационной и морской медицины. Кафедра была признана лучшей физиологической кафедрой во всем мире. Однако по воле И.В. Сталина в 1950 г. Л.А. Орбели был освобожден от занимаемой должности. Это привело к значительному ослаблению кафедры, хотя в течение четырех лет ее возглавляли академик АМН СССР А.В. Лебединский и профессор М.П. Бресткин, которые прежде были заместителями Л.А. Орбели
2001 г. заведование кафедрой было доверено член-корреспонденту РАМН, профессору В.О. Самойлову, а заведующим учебной частью стал профессор В.Н. Голубев. Оба ученики А.С. Мозжухина. Вместе с ними на кафедре трудятся доценты И.М. Алекперов, Е.В. Антоненкова, Ю.Н. Королев, старший преподаватель Е.Б. Филиппова, преподаватели В.В. Бекусова, Е.М. Лесова. Тарханов в 1879 году появляется его монография «О психомоторных центрах и развитии их у человека и животных». В том же году он переводит «Технический учебник гистологии» Ранвье и тогда же выходит его статья «О применении телефона в животном электричестве». Под его редакцией переводится «Общая мышечная и нервная физиология» И. Розенталя, а также «Опыты над естественным сном животных». В лаборатории И. Р. Тарханова были начаты опыты по изучению влияния различных ядов на биоэлектрические процессы в нервно-мышечной системе у лягушки. Им была открыта кожно-гальваническая реакция (феномен Тарханова), к-рая используется для изучения функционального состояния в. н. с., исследований в области физиологии труда, спорта. Одним из первых (1896) исследовал действие на ц. н. с. ионизирующего излучения. Выдающийся ученый Л.А. Орбели является автором более 130 фундаментальных и крупных научных работ. Он создал новое научное направление — эволюционную физиологию, впервые получил объективные данные о зрительной функции животного, зависящей от деятельности полушарий головного мозга. Орбели обосновал учение об адаптационно-трофической функции симпатической нервной системы и об адаптационнотрофической роли вегетативной нервной системы.
4. Основные свойства живых тканей. Раздражимость и возбудимость. Классификация раздражителей. Основные свойства живых тканей: 1. Специфическая организация – клетка – элементарная структурно-функциональная единица живой системы 2. Специфический метаболизм - это ряд химических и физических превращений в живых клетках, обеспечивающих их жизнедеятельность во взаимосвязи с внешней средой 3. Способность к самоорганизации и саморегуляции на всех уровнях биол системы 4. Подвижность (актин-миозиновая система, тубилин-динеиновая) 5. Рост и развитие 6. Наследственность и изменчивость 7. Реактивность (раздражимость) Раздражимость-Способность системы реагировать на факторы внешней среды переходом из состояния покоя в состояние активности, либо изменением уровня функциональной активности. (Все ткани) Возбудимость – Способность живых клеток воспринимать изменение внешней среды и отвечать на это изменение реакцией возбуждения (нервные мышечные железистые) Клетки, способные к генерации ПД – возбудимые, сам процесс – возбуждение. Раздражители: По силе: 1. Пороговые – приводят к возникновению ПД 2. Подпороговые – ниже порога возбуждения 3. Максимальные – минимальная сила раздражителя, вызывающая максимальный ответ 4. Субмаксимальные – сила меньше максимальной силы 5. Супермаксимальные – сила больше максимальной силы По природе: 1. Физические 2. Химические 3. Физико-химические По биологической значимости: 1. Адекватные – раздражитель, к восприятию которого данная ткань приспособилась в процессе эволюции (орган слуха – к упругим механическим колебаниям среды) 2. Неадекватные – раздражитель, который в естественных условиях ж/д организма не вызывает возбуждение живой ткани
5. Возбудимые ткани. Биоэлектрические явления в возбудимых тканях. Опыты Луиджи Гальвани и Алессандро Вольта. Возбудимыми тканями, являются такие ткани, клетки которых способны в ответ на раздражение генерировать специфическую реакцию - возбуждение. К ним относятся нервные, мышечные и некоторые секреторные клетки. Признаки возбуждения: 1. Специфическая реакция (Нервная ткань – нервный импульс, мышечная – сокращение) 2. Генерация ПД 3. Проводимость – бездекрементное распространение ПД по возбудимым мембранам 4. Уменьшение эл импеданса (сопротивления) вследствие повышения проницаемости возбудимых мембран для ионов 5. Усиление метаболизма (преимущественно катаболизма, повышение потребления О2 и субстратов метаболизма) 6. Повышение температуры возбужденной ткани Биоэлектрические явления — те же, что и животное электричество, процессы распределения и транспорта электрических зарядов в живом организме, обусловленные присутствием в живых клетках и тканях большого количества фиксированных (заряженные группы био-макромолекул) и подвижных (свободные ионы и электроны) электрических зарядов. Биоэлектрические явления обусловливают возникновение возбуждения и его проведение по нервным волокнам, являются причиной процессов сокращения мышечных волокон скелетных, гладких и сердечных мышц, выделительной функции железистых клеток. Биоэлектрические явления лежат в основе процессов всасывания в желудочно-кишечном тракте, в основе восприятия вкуса, запаха, в основе деятельности всех анализаторов. Нет физиологического процесса в живом организме, который в той или иной форме не был бы связан с биоэлектрическими явлениями. Гальвани стал одним из основателей учения об электричестве. Луиджи Гальвани также известен тем, что он первый обратил внимание на то, что электрические явления возникают при мышечном сокращении (этот эффект, а точнее, явление, был назван «животным электричеством») Его заинтересовала способность мертвого препарата проявлять себя как живой материал. Он менял положение металлического провода в теле лягушки, менял источники тока и множество других параметров. Проводя такой опыт, Луиджи Гальвани хотел использовать в качестве источника тока природное электричество, но погода стояла ясная и на небе не было ни облачка. Ученый чисто случайно прижал электроды, которые были воткнуты в спинной мозг лягушки, к железной решетке, на которой и лежала лягушка. Луиджи Гальвани был очень сильно удивлен, когда увидел, что появились такие же сокращения, как и во время опытов, которые проводились во время грозы. Еще больше Луиджи Гальвани был удивлен, когда выяснил, что мышцы сокращаются и в то время, когда внешний источник тока отсутствует. Оказалось, что мышцы начинают сокращаться и при простом наложении на них двух пластин разных металлов, соединенных проводником. Этими опытами физиолога Луиджи Гальвани заинтересовался другой известный ученый – физик Алессандро Вольта. Вольта высказал предположение, что электричество заключается в тех двух пластинах разных металлов, которые использовал Гальвани. И электричество возникает при соединении этих пластин проводником. Таким образом, физик Алессандро Вольта стал оппонентом в научном споре физиолога Луиджи Гальвани. Так начался величайший спор между двумя учеными. Алессандро Вольта настаивал на том, что источник электричества – это металлы, а другой настаивал на том, что источник тока – это животные. Оба ученых проводили эксперименты в подтверждение своей теории. Луиджи Гальвани, как ему показалось, нашел неопровержимые доказательства своей точки зрения, которая состоит из двух элементов: 1) доказал, что электричество возникает и без участия металлов; 2) сняв кожный покров с нерва лапки лягушки, Луиджи Гальвани поднес его к мышцам. Мышца начала сокращаться. Алессандро Вольта, однако, не успокоился и не отступился. Он тоже привел весьма и весьма убедительные доказательства в пользу своей точки зрения. Хотя и Гальвани, и Вольта считали, что в споре прав только один из них, по прошествии продолжительного периода времени стало ясно, что обе точки зрения имеют право на существование. Важнейшим его вкладом в развитие науки было изобретение им принципиально нового источника постоянного тока. В 1800 г. Алессандро Вольта создал так называемый вольтов столб. Это был первый химический источник электричества. Гальвани проиграл спор с Вольтом. Аргументы Вольта казались вполне убедительными. В 1797 г наступает окончательный крах: по политическим мотивам Гальвани выгнали из университета. Он лишился возможности работать и через год умер.
6.Потенциал покоя: механизмы генерации. ПП – разность электрических потенциалов между внутренней и наружной средой клетки в состоянии покоя (измерение – прокол осциллографа) Величина ПП – (-30-90 мВ) В волокнах скелетных мышц от -60 до -90 мВ В нервных клетках от -50 до -80 мВ В гладких мышцах от -30 до -70 мВ В сердечной мышце от -80 до -90 мВ Роль – основа для возникновения ПД ПП влияет на транспорт ионов и воды Причина существования ПП – 1)разная концентрация анионов и катионов внутри и вне клетки (Натрия больше вне клетки, Калия больше в клетке, Хлора больше вне клетки) 2)Неравномерное распределение ионов: - Неодинаковая проницаемость клеточных мембран -Работа ионных насосов Калий – основной ион в формировании ПП Внутри отрицательно, снаружи положительно При уменьшении конц Калия ПП уменьшается, при увеличении конц Калия ПП увеличивается. В покое – устанавливается динамическое равновесие между входящими и выходящими ионами Калия Вклад Хлора и Натрия в ПП невелик, так как проницаемость ниже, чем у Калия Мембранный потенциал покоя возникает вследствие дефицита положительных зарядов внутри клетки, возникающий за счет работы Натрий-калиевого насоса и последующей утечке из клетки положительно-заряженных ионов Калия
7.Электрофизиология возбуждения: потенциал действия. ПД – Быстрое колебание величины мембранного потенциала вследствие инактиваии и активации ионных каналов и диффузии ионов из клетки и в клетку. (Перезарядка мембраны) Роль ПД – передача нервных импульсов между нервными клетками, нервными центрами, органами. Параметры: 80 – 130 мВ ПД подчиняется закону «Все или ничего» Механизм: быстрое движение Натрия в клетку, Калия – из клетки Фазы ПД: 1. Деполяризация – уменьшение заряда клетки до нуля, разв при действии деполяризующего раздражителя. Натриевые каналы открываются, Натрий поступает в клетку – мембр потенциал уменьшается до нуля. 2. Инверсия (Овершут) – изменение знака заряда клетки на противоположный, включает восход и нисход части пика. Восходящая – обеспечивается входом Натрия в клетку, после исчезновения мембранного потенциала вход Натрия в клетку продолжается – число положит ионов больше отриц ионов, заряд внутри клетки Положительный, закрытие Натриевых каналов, Натрий не поступает. Нисходящая – открытие Калиевых каналов, возрастает выход Калия из клетки, уменьшается число положительно заряженных ионов – заряд клетки уменьшается – продолжается до полного исчезновения положительного заряда внутри клетки. 3. Реполяризация –восстановление заряда клетки – продолжается быстрый выход Калия из клетки. 4. Гиперполяризация – в результате еще сохранившейся повышенной проницаемости мембраны для калия. Калиевые каналы не полностью закрыты, Калий продолжает выходить. Раздражители: По силе: 1. Пороговые – приводят к возникновению ПД 2. Подпороговые – ниже порога возбуждения 3. Максимальные – минимальная сила раздражителя, вызывающая максимальный ответ 4. Субмаксимальные – сила меньше максимальной силы 5. Супермаксимальные – сила больше максимальной силы По природе: 1. Физические 2. Химические 3. Физико-химические По биологической значимости: 1. Адекватные – раздражитель, к восприятию которого данная ткань приспособилась в процессе эволюции (орган слуха – к упругим механическим колебаниям среды) 2. Неадекватные – раздражитель, который в естественных условиях ж/д организма не вызывает возбуждение живой ткани Максимальная сила раздражителя – минимальная сила раздражителя, вызывающая максимальный ответ
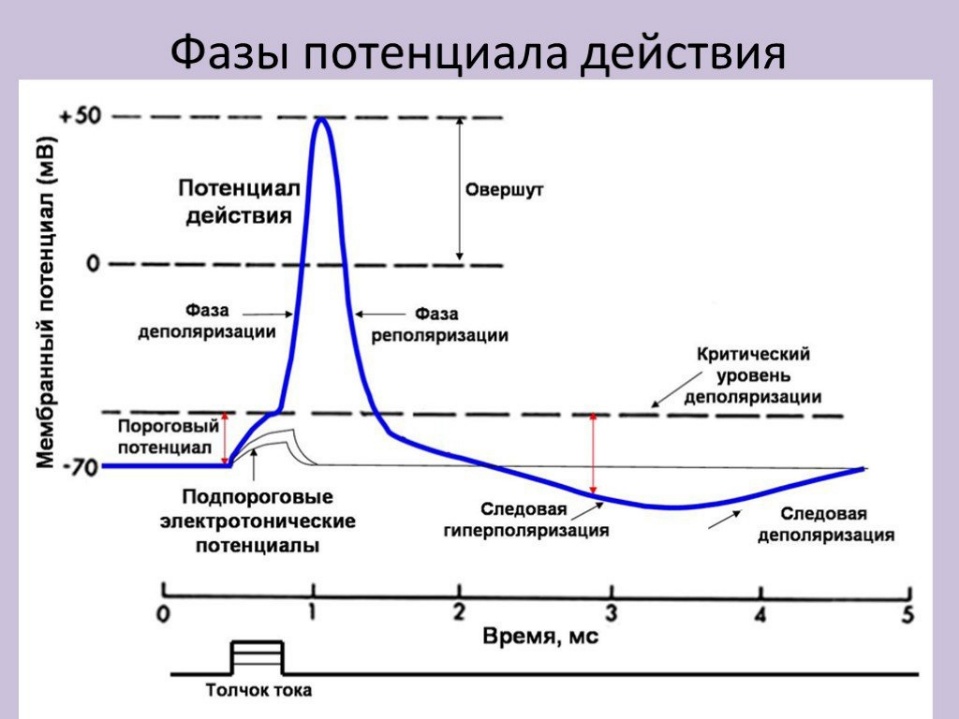
8.Мера (количественная характеристика) возбудимости.
Основной количественной мерой возбудимости является порог раздражения.
Порог раздражения – минимальная сила раздражения, которая вызывает распространяющееся возбуждение, характеризующееся потенциалом действия. Величина порога раздражения для каждой клетки индивидуальна и зависит от разности между уровнем заряда мембраны и уровнем критической деполяризации. Возбудимость и порог раздражения находятся обратно пропорциональной зависимости. Чем больше порог, тем меньше возбудимость и наоборот. Нервная ткань характеризуется высокой возбудимостью и низким порогом, а мышечная ткань – низкой возбудимостью и высоким порогом.
9.Рефрактерность: фазовые изменения возбудимости при генерации потенциала действия. Лабильность Рефрактерность – способность ткани терять или снижать возбудимость в процессе возбуждения.
1. АРФ – полная утрата возбудимости – соответствует времени развития деполяризации. В течение АРФ возбудимая мембране не может генерировать новый ПД, даже если на нее подействовать сколь угодно сильным раздражителем. Природа АРФ – во время деполяризации все ПИК открыты, и дополнительные стимулы не могут вызвать воротный процесс (им не на что действовать) 2. ОРФ – возбудимость от нуля возвращается к исходному уровню, когда сильное раздражение(сверхпороговое) может вызвать новое возбуждение – соответствует реполяризации. Все большее число ПИК возвращается в закрытое состояние и ПИК вновь готовы к следующему переходу в открытое состояние под действием отдельных импульсов 3. Экзальтация – период повышенной возбудимости – связан с изменением свойств сенсора напряжения во время возбуждения. Она соответствует следовой деполяризации. Очередной ПД можно вызывать более слабым раздражением, поскольку мембранный потенциал несколько ниже обычного и он оказывается ближе к критическому уровню деполяризации, что объясняют повышенной проницаемостью клеточной мембраны для ионов Nа+. КМП ПРИБЛИЖАЕТСЯ К ПП. Возбудимость меняется за счет изменения КМП. 4. Пониженная возбудимость- следовая гиперполяризация. Лабильность (Введенский) – Способность возбудимых тканей отвечать максимальным числом возбуждения в единицу времени на их электрическое раздражение Мера лабильности – макс число ПД, которое ткань может воспроизвести за 1 секунду (Нерв 200 – 400 Гц, ППМТ 100 – 200 Гц) Лабильность определяется продолжительностью рефрактерного периода (чем короче рефрактерный период, тем больше лабильность).

10. КРИВАЯ «СИЛА-ДЛИТЕЛЬНОСТЬ». РЕОБАЗА, ХРОНАКСИЯ. Реобаза – наименьшая сила эл тока, способная вызвать возбуждение ткани, независимо от длительности действия. Чем выше возбудимость, тем ниже пороговая сила. Важное условие для возбуждения – полезное время – мин время, в течение которого должен действовать на ткань раздражитель пороговой силы, чтобы вызвать возбуждение. После КМП ПД развивается вне зависимости от раздражителя. Хронаксия (исп в медицине вместо порогового времени) – наименьшее время, в течение которого должен действовать эл ток в 2 Реобазы, чтобы вызвать возбуждение (Хронаксия для мышц конечностей 0.1 – 0.7 мс) Кривая показывает, что с увеличением сверхпороговой силы раздражителя время его действия, необходимое для возбуждения, уменьшается и наоборот. 1. Правая часть графика – если использовать раздражитель меньшей амплитуды, чем реобаза, то возбуждения не возникнет 2. Левая сторона – если использовать раздражитель с длительностью меньше критического интервала – возбуждение не возникнет, даже если раздражитель бесконечно большой


11. ПЕРВИЧНЫЕ И ВТОРИЧНЫЕ ЭЛЕКТРОТОНИЧЕСКИЕ ЯВЛЕНИЯ. ИСПОЛЬЗОВАНИЕ В МЕДИЦИНЕ. Электротонические явления – изменение возбудимости под влиянием постоянного подпорогового электрического тока, развивающееся в нервной и мышечной тканях. Под катодом – деполяризация, под анодом – гиперполяризация Сильнее тем больше, чем пропускаемый ток сильнее Под катодом возникает катэлектротон (повышение возбудимости под катодом) Под анодом возникает анэлектротон (уменьшение возбудимости под анодом) Катэлектротон + анэлектротон – первичные электротонические явления Существуют только при недолгом воздействии постоянного подпорогового эл тока. При длительном действии тока – извращение возбудимости (мех – ослабление проницаемости потенциал-зависимых Натриевых каналов и усиление проницаемости Калиевых каналов) – возникают вторичные электротонические явления: Катодическая депрессия – уменшение возбудимости под катодом Анодическая экзальтация – повышение возбудимости под анодом Физиотерапевтическая процедура (гальванизация) – управление возбудимостью тканей при помощи подпорогового эл тока, который изменяет уровень ПП, но не сдвигает КМП (уменьшение боли при процедуре) ДОЛГО - БОЛЬ При кратковременном пропускании под порогового постоянного электрического тока изменяется возбудимость ткани под стимулирующими электродами. Под катодом возбудимость увеличивается (катэлектрон), под анодом возбудимость уменьшается (анаэлектрон). Это связано с тем, что микроэлектродные исследования показали, что под катодом происходит пассивная деполяризация клеточной мембраны, под анодом гиперполяризация. Значение критического потенциала не изменяется – изменяется за счёт мембранного потенциала покоя. При длительном воздействие подпорогового постоянного тока изменяется не только мембранный потенциал покоя, но и значение критического потенциала. Под катодом – смещение уровня критического потенциала вверх (Инактивация натриевых каналов). Под анодом – смещение уровня критического потенциала вниз. В результате под катодом возбудимость снижается - катодическая депрессия Вериго. Под анодом - повышается (анодическая экзальтация)
12. Виды мышечной ткани, их свойства и функции. Механизмы сокращения гладких мышечных волокон Виды мышечной ткани: 1)гладкая мышечная ткань. Состоит из одноядерных клеток — миоцитов веретеновидной формы длиной 20—500 мкм. Их цитоплазма в световом микроскопе выглядит однородно, без поперечной исчерченности. Обладает особыми свойствами: медленно сокращается и расслабляется, обладает автоматией, является непроизвольной (ее деятельность не управляется по воле человека). Входит в состав стенок внутренних органов: кровеносных и лимфатических сосудов, мочевыводящих путей, пищеварительного тракта (сокращение стенок желудка и кишечника); 2)поперечнополосатая скелетная мышечная ткань. Состоит из миоцитов, имеющих большую длину (до нескольких сантиметров) и диаметр 50—100 мкм; эти клетки многоядерные, содержат до 100 и более ядер; в световом микроскопе цитоплазма выглядит как чередование тёмных и светлых полосок. Свойства - высокая скорость сокращения, расслабления и произвольность (её деятельность управляется по воле человека). Входит в состав скелетных мышц, стенки глотки, верхней части пищевода, ей образован язык, глазодвигательные мышцы. Волокна длиной от 10 до 12 см; 3)поперечнополосатая сердечная мышечная ткань. Состоит из 1 или 2-х ядерных кардиомиоцитов, имеющих поперечную исчерченность цитоплазмы (по периферии цитолеммы). Кардиомиоциты разветвлены и образуют между собой соединения — вставочные диски, в которых объединяется их цитоплазма. Существует также другой межклеточный контакт - анастомозы (впячивание цитолеммы одной клетки в цитолемму другой). Этот вид мышечной ткани образует миокард сердца. Особое свойство - автоматия — способность ритмично сокращаться и расслабляться под действием возбуждения, возникающего в самих клетках (типичные кардиомиоциты). Эта ткань является непроизвольной (атипичные кардиомиоциты). Существует 3-й вид кардиомиоцитов - секреторные кардиомиоциты (в них нет фибрилл). Они синтезируют гормон тропонин, понижающий артериальное давление и расширяющий стенки кровеносных сосудов. Процесс сокращения гладкомышечных во локон совершается по тому же механизму скольжения нитей актина и миозина относи тельно друг друга, что и в скелетных мышцах. Однако у гладкомышечных клеток нет той стройной аранжировки сократительных бел ков, как у скелетных мышц. У этих клеток миофибриллы с саркомерами расположены нерегулярно, поэтому клетка не имеет по перечной исчерченности. Электромеханичес кое сопряжение в этих клетках идет иначе, чем в скелетных мышцах, так как в них слабо выражен саркоплазматический ретикулум. В связи с этим триггером для мышечного со кращения служит поступление ионов Са2+в клетку из межклеточной среды в процессе ге нерации ПД. Того количества кальция, кото рое входит в клетку при возбуждении, вполне достаточно для полноценного фазного сокра щения. Инициация сокращений гладких мышц с помощью ионов Са2+также имеет несколько другой механизм, чем в поперечнополосатых волокнах. Ионы Са2+воздействуют на белок кальмодулин, который активирует киназы легких цепей миозина. Это обеспечивает перенос фосфатной группы на миозин и сразу вызывает срабатывание, т.е. сокраще ние, поперечных мостиков. О существовании тропонин-тропомиозиновой системы сведе ний не имеется.
При снижении в миоплазме концентрации ионов Са2+фосфатаза дефосфорилирует миозин, и он перестает связы ваться с актином. Скорость сокращения гладких мышц невелика — на 1—2 порядка ниже, чем у скелетных мышц. Сила сокра щений некоторых гладких мышц позвоноч ных не уступает силе сокращений скелетных мышц. Подобно сердечной и скелетной мускулатуре, гладкие мышцы всегда расслабляются при падении внутриклеточной концентрации Са2+ ниже 10-7М. Однако их рас слабление происходит гораздо медленнее, посколь ку скорость поглощения ионов Са2+ саркоплазматическим ретикулумом или удаления их через кле точную мембрану здесь ниже. Удаление Са2+при водит к расщеплению фосфатазой функционально важной фосфатной группы миозина. Его дефосфорилированные головки теряют способность образо вывать поперечные мостики с актином. Пока неяс но, каким образом образующиеся в гладкомышечных клетках цАМФ и цГМФ вызыва ют понижение их тонуса. Возможно, цАМФ ингибирует активность киназы легких цепей миозина или усиливает поглощение Са2+саркоплазматическим ретикулумом. С другой стороны, вполне вероятна роль цГМФ как внутриклеточного посредника в рас слаблении гладких мышц сосудов, которое индуци руется расслабляющим фактором эндотелия.
13. Гетерогенность миоцитов скелетных мышц. Электромеханическое сопряжение в миоците скелетной мышцы. Волокна скелетных мышц неодинаковы по своим механическим и метаболическим особенностям. Типы волокон различаются на основе следующих характеристик: в зависимости от максимальной скорости укорочения – волокна быстрые и медленные; в зависимости от главного пути образования АТФ – волокна оксидативные и гликолитические. На основании двух характеристик (скорость укорочения и тип метаболизма) выделяют три типа волокон скелетных мышц. 1. Медленные фазические оксидативные волокна (тип I). Характеризуются низкой активностью миозиновой АТФазы, высокой окислительной способностью, низким содержанием гликогена и высоким содержанием миоглобина. Медленные волокна генерируют медленные, продолжительные сокращения и устойчивы к утомлению. 2. Быстрые фазические оксидативные волокна (тип IIа). Характеризуются высокой активностью миозиновой АТФазы, высокой окислительной способностью, высокое содержание миоглобина. 3. Быстрые фазические гликолитические волокна (тип IIб). Для волокон этого типа характерна высокая активность миозиновой АТФазы и высокая гликолитическая способность. Содержание миоглобина низкое. Быстрые волокна отвечают короткими, энергичными и более сильными сокращениями. У человека нет мышц, состоящих только из быстрых или только из медленных мышечных волокон, каждая мышца содержит разные типы мышечных волокон. Волокна различаются не только по своим биохимическим особенностям, но и по размерам: у гликолитических волокон диаметр больше, чем у оксидативных. Это влияет на величину развиваемого ими напряжения. Число толстых и тонких филаментов на единицу площади поперечного сечения примерно одинаково для всех типов скелетных мышечных волокон. Следовательно, чем больше диаметр волокна, тем большее число параллельно задействованных толстых и тонких филаментов участвует в генерировании силы, и тем больше максимальное напряжение мышечного волокна. Гликолитическое волокно, имеющее больший диаметр, развивает в среднем более значительное напряжение, по сравнению с напряжением оксидативного волокна. Рассмотренные три типа мышечных волокон характеризуются разной устойчивостью к утомлению. Быстрые гликолитические волокна утомляются через короткое время, а медленные оксидативные волокна выносливы, что позволяет им длительно поддерживать сократительную активность практически при постоянном уровне напряжения. Быстрые оксидативные волокна занимают промежуточное место по способности противостоять развитию утомления. Для мышечных волокон перечисленных групп характерно наличие одной, в крайнем случае нескольких концевых пластинок, образованных одним двигательным аксоном.
4. Тонические волокна. В отличие от предыдущих типов мышечных волокон в тонических волокнах двигательный аксон образует множество синаптических контактов с мембраной мышечного волокна. Развитие сокращения и расслабления происходит медленно, что обусловлено низкой активностью миозиновой АТФазы. Мышечные волокна этого типа эффективно работают в изометрическом режиме. Эти мышечные волокна не генерируют потенциал действия и не подчиняются закону «все или ничего». Одиночный импульс вызывает незначительное сокращение. Серия импуль сов вызовет суммацию постсинаптических потенциалов и плавно возрастающую деполяризацию мышечного волокна. У человека мышеч ные волокна этого типа входят в состав наружных мышц глаза. Последовательность процессов, в результате которых потенциал действия Сарколеммы приводит к запуску сокращения мышцы – цикл поперечных мостиков Совокупность явлений, обусловливающих связь между возбуждением (ПД) и сокращением мышечных волокон – электромеханическое сопряжение. К регулярно расположенным Т‑трубочкам с двух сторон подходят терминальные цистерны саркоплазматического ретикулума (САР). Т‑трубочка и расположенные с двух сторон от неё цистерны образуют триаду. Z‑мембраны и зоны перекрытия актиновых и миозиновых нитей расположены рядом с триадами. Такая структура необходима для эффективного электромеханического сопряжения. + Электромеханическое сопряжение включает следующие процессы: 1)образование потенциала концевой пластинки (ПКП); 2)электротоническое распространение ПКП в околосинаптическую область; 3)генерация потенциала действия мышечного волокна в околосинаптической области; 4)распространение потенциала действия по поверхно стной мембране мышечного волокна; 5)распространение потенциала действия по мембранам поперечных трубочек (Т‑системы); 6)реакция рецепторов дигидропиридина на изменение мембранного потенциала; 7)передача сигнала дигидропиридиновыми рецепторами рианодиновым рецепторам терминальных цистерн саркоплазматического ретикулума; 8)открытие каналов Ca2+ с рианодиновыми рецепторами и выход Ca2+ из саркоплазматического ретикулума (цистерн) в саркоплазму (цитозоль); 9)диффузия Ca2+ в межфибриллярное пространство; 10)связывание Ca2+ с тропонином, расположенным на тонком, актиновом, миофиламенте; 11)конформационные изменения тропонина при связывании Са2+ => «заталкивание» тропомиозина в канавки актиновых нитей.
14. ТИПЫ И РЕЖИМЫ МЫШЕЧНОГО СОКРАЩЕНИЯ. 3 Типа мышечного сокращения: Разная степень укорачивания мышц: 1. Изометрический 2. Изотонический 3. Ауксотонический (анизотонический) Изометрическое – длина мышцы почти не меняется, увеличение напряжения, вся сила затрачивается на развитие напряжения – статика, поддержание позы Изотоническое – укорочение волокна в условиях постоянной внешней нагрузки, напряжение сохраняется, происходит быстрое укорочение мышцы, сменяющееся расслаблением, описывается уравнением Хилла (бицепс) Ауксотоническое – в процессе сокращения мышцы и укорачиваются, и напрягаются 2 режима сокращения скелетных мышц: 1. Одиночное – при действии одиночного стимула с 3 стадиями – латентный период, напряжения и расслабление. Золотая пропорция – соотношение двух этих фаз (1.618) 2. Тетаническое – сокращение, при котором происходит суммация одиночных мышечных сокращений в результате ее раздражения частыми (тетаническими) стимулами. Сила сокращения в 2-4 раза больше, чем при одиночном. Амплитуда увеличивается благодаря наложению (суперпозиции) следующих друг за другом одиночных сокращений. По мере повышения частоты стимуляции сначала возникает зубчатый тетанус, если последующий стимул поступает к мышце на стадии ее расслабления. Если интервал между стимулами меньше длительности стадии укорочения мышцы, то возникает гладкий тетанус - сокращение Оптимум частоты – частота раздражающих импульсов каждый последующий импульс совпадает с фазой повышенной возбудимости-тетанус наибольшей амплитуды Пессимум частоты - Более высокая частота разрешение – каждым последующий импульс тока в фазу рефрактерности – амплитуда тетануса уменьшается


15. Организм как открытая саморегулирующаяся система. Стратегия регулирования функций. Внутренняя среда организма. Гомеостазис. Жёсткие и пластичные гомеостатические константы
Живой организм – открытая система, имеющая связь с окружающей средой посредством нервной, пищеварительной, дыхательной, выделительной систем и др.
В процессе обмена веществ с пищей, водой, при газообмене в организм поступают разнообразные химические соединения, которые в организме подвергаются изменениям, входят в структуру организма, но не остаются постоянно. Усвоенные вещества распадаются, выделяют энергию, продукты распада удаляются во внешнюю среду. Разрушенная молекула заменяется новой и т.д.
Организм – открытая, динамичная система. В условиях непрерывно меняющейся среды организм поддерживает устойчивое состояние в течение определенного времени.
Внутренняя среда организма (клод Бернар)– внеклеточные жидкости, интерстициальная (тканевая)-10л, кровь-3л, лимфа-2л, ликвор, внутриглазная, синовиальная. Жидкости характеризуются постоянством физико-химических показателей: осмоляльность, рН, температура, постоянный объем
Жесткая константа – показатели внутренней среды организма, отклонения от средней величины малое, ведут к необратимым наращениям метаболизма и смерти (концентрация Натрий, Калий, рН, парциальное напряжение кислорода)
Пластическая константа – отклонения от средней величины имеет большой диапазон, без существенных нарушений физ функций (объём внеклеточной жидкости, вязкость, клеточный состав крови)
Выход за нормы – заболевание
Рецепторы следят за изменением, устраняют сдвиги
У. Кэннон – гомеостазис – состояние относительного и динамического постоянства показателей внутренней среды организма, отклонение от которого устраняется регуляторным механизмом
Пример Жесткой константы – концентрация Натрия = 140+- 5 ммоль/л, рН крови = 7.36-7.4, осмотическое давление 25-30 мм.рт.ст
Пластичные – глюкоза 3.3-5.5 ммоль/л, Калий 3.4-5.6 ммоль/л, Частота пульса = 60-90 в покое
Количество гемоглобина у мужчин – 130-160 г/л, у женщин 120-140 г/л, СОЭ (М) 1-10 мм/час, (Ж) 2-15 мм/час
16. Механизмы регуляции функций в организме человека Внутриклеточные системы служат ключевыми звеньями всех форм физиологических процессов: местной, гуморальной, нервной 1. Местная регуляция – информационное взаимодействие между соседними клетками, сигналы химич и эл природы. Химич связь через 2ичные мессенджеры, образование комплексов с элементами цитозоля (прохождение через мембрану) – НО, СО, помимо этого есть щелевые контакты. Данный вид рег – объединение клеток в ткань. 2. Гуморальная регуляция – перенос физиологически активных веществ жидкостями, перемещающимися в организме – кровь, лимфа, ликвор. Объединяет органы и ткани всего организма. БАВ (действует на рецептор, проникает через мембрану) 3. Нервная регуляция – вся соматическая активность обеспечивается нервной регуляцией. Рефлекс – реакция организма на изменение вешней среды, протекающая при участии ЦНС. +местная Осуществляется: 1)креаторными связями; 2)метаболитами (молочная кислота, пировиноградная кислота, аденозин); 3)ионами; 4)БАВ.
17. Физиологическая система: общие принципы организации. Физиологическая система – наследственно закрепленная, регулируемая система органов и тканей, которая функционирует в организме во взаимодействии друг с другом. Стратегии регулирования биол системы – при любых условиях, при любых изменениях окр среды сохранять жизнеспособность. Система регулирования состоит из 3-х уровней: 1. Состоит из относительно автономных местных систем, которые поддерживают физиологические константы, задаваемые метаболическими потребностями организма – местная саморегуляции 2. Приспособительные реакции в связи с изменением внутренней среды (спинной или продолговатый мозг, или эндокринные железы) – задается величина физиологических параметров, которые поддерживаются системой 1го уровня 3. Образования головного мозга, обеспечивающих анализ и оценку состояния внутренней и внешней среды, настройку режимов систем 1го и 2го уровней Биша считал, что ж/д человека проявляется в вегетативных, соматических, психических процессах: 1. Суть вегетатики – обмен веществ и энергии между орг и окр средой. Регулируется дыханием, пищеварением, кровообращением итд. 2. Соматика – двигательная активность 3. Психические процессы – приспособление к обществу, влияние на окр ср
18. Понятие о функциональной системе Функциональная система — динамическая совокупность органов и тканей, относящихся к различным анатомо-физиологическим структурам и объединившихся для достижения определенной приспособительной деятельности (полезного приспособительного результата). В основе функциональной системы лежит принцип возвращения к норме той или иной величины. Каждая функциональная система возникает в том случае, если какая-либо величина отклоняется от нормы. Функциональная система — это временное образование, до достижения определенного результата. Цель работы функциональной системы — возвращение величины к норме. Организм человека — совокупность различных функциональных систем. Из всех функциональных систем в данный момент есть одна — доминирующая. Каждая функциональная система состоит из 4-х звеньев: 1. центральное звено — совокупность нервных центров, регулирующих ту или иную функцию; 2. исполнительное звено — органы и ткани, которые работают для достижения результата (сюда включаются поведенческие реакции); 3. обратная связь (афферентация) — после работы второго звена возникает вторичный поток импульсов от рецепторов в центральную нервную систему, идет информация об изменении той или иной величины; 4. полезный результат — для достижения которого и работает функциональная система. Каждая функциональная система обладает 2-мя свойствами: 1. динамичность — каждая функциональная система — это образование временное. Различные органы могут входить в состав одной функциональной системы, одни и те же органы могут входить в состав различных функциональных систем; 2. саморегуляция — функциональная система обеспечивает поддержание на постоянном уровне различных параметров без вмешательства из вне. Все функциональные системы работают по принципу опережения. При отклонении от нормы величины импульсы поступают в центральное звено, и там формируется эталон будущего результата. Затем начинает работать 2-е звено. Как только полученный результат будет соответствовать эталону, то функциональная система распадается.
Выделяют два типа функциональных систем. Функциональные системы первого типа обеспечивают само регуляцию функционирования систем организма, направленных на возможность его существования в данных условиях среды. Функциональные системы второго типа обеспечивают приспособительный эффект через изменение поведения. Именно этот тип функциональных систем лежит в основе различных поведенческих актов. Согласно П.К. Анохину, функциональная система второго типа состоит из следующих стадий: • афферентный синтез;
• стадия принятия решения;
• стадия акцепторов результата действия;
• эфферентный синтез (программа действия);
• само действие;
• оценка достигнутого результата. Афферентный синтез представляет собой объединение всей сенсорной информации, поступающей в мозг. Его содержание определяется мотивационным возбуждением, памятью. Любая информация, поступающая информация соотносится с доминирующим в настоящее время мотивационным возбуждением. Пусковая афферентация определяет то возбуждение, которое будет формироваться в сенсорной системе под влиянием внешнего биологически значимого раздражителя. Распределение раздражителей во времени и пространстве определяет обстановочную афферентацию (при изменении последовательности действий (обстановки) условный рефлекс может не проявляться).
Функциональная роль пусковых и обстановочных афферентаций обусловлена прошлым опытом человека, хранящегося в виде памяти. На основе взаимодействия мотивационного, обстановочного возбуждения и памяти формируется так называемая интеграция или готовность к определенному поведению. Чтобы она трансформировалась в определенное целенаправленное поведение требуется воздействие со стороны пусковых раздражителей (пусковая афферентация). Внешним проявлением афферентного синтеза, обусловленного влияниями лимбической системы и ретикулярной формации на кору, является активизация ориентировочно-исследовательского поведения. Завершение этой стадии сопровождается переходом в стадию принятия решения, которая определяет тип и направленность поведения, этот этап реализуется через формирование аппарата акцепторов результата действия, программирующий результаты будущих событий. Эфферентный синтез или стадия программа действия осуществляет интеграцию соматических и вегетативных возбуждений в целостный поведенческий акт. Эта стадия характеризуется тем, что действие уже сформировано как нервный процесс, но внешне оно еще не реализуется. На основании этой программы происходит конкретное действие, результаты которого благодаря наличию обратной афферентации сопоставляются с акцептором результатов действия. В случае достижения желаемого результата действие прекращается, в противном случае в программу поведения вносятся соответствующие корректировки.
19.Типы регуляции функций в организме человека. Принцип обратной связи в регуляции функций. Регуляция по отклонению — циклический механизм, при котором всякое отклонение от оптимального уровня регулируемого показателя мобилизует все аппараты функциональной системы к восстановлению его на прежнем уровне. Регуляция по отклонению предполагает наличие в составе системного комплекса канала отрицательной обратной связи, обеспечивающего разно направленное влияние: усиление стимулирующих механизмов управления в случае ослабления показателей процесса или ослабление стимулирующих механизмов в случае чрезмерного усиления показателей процесса. Например, при повышении АД включаются регуляторные механизмы, обеспечивающие снижение АД, а при низком АД включаются противоположные реакции. В отличие от отрицательной обратной связи, положительная обратная связь, встречающаяся в организме редко, оказывает только однонаправленное, усиливающее влияние на развитие процесса, находящегося под контролем управляющего комплекса. Поэтому положительная обратная связь делает систему неустой чивой, неспособной обеспечить стабильность регулируемого процесса в пределах физиологического оптимума. Например, если бы артериальное давление регулировалось по принципу положи тельной обратной связи, в случае снижения артериального давления действие регуляторных механизмов привело бы к еще большему его снижению, а в случае повышения - к еще большему его увеличению. Примером положительной обратной связи явля ется усиление начавшейся секреции пищеварительных соков в желудке после приема пищи, что осуществляется с помощью продуктов гидролиза, всосавшихся в кровь. Регуляция по опережению заключается в том, что регули рующие механизмы включаются до реального изменения парамет ра регулируемого процесса (показателя) на основе информации, поступающей в нервный центр функциональной системы и сигна лизирующей о возможном изменении регулируемого процесса в будущем. Например, терморецепторы (детекторы температуры), находящиеся внутри тела, обеспечивают контроль за температурой внутренних областей тела. Терморецепторы кожи, в основном, иг рают роль детекторов температуры окружающей среды. При зна чительных отклонениях температуры окружающей среды создают ся предпосылки возможного изменения температуры внутренней среды организма. Однако в норме этого не происходит, так как им-пульсация от терморецепторов кожи, непрерывно поступая в гипо-таламический терморегуляторный центр, позволяет ему произве сти изменения работы эффекторов системы до момента реального изменения температуры внутренней среды орга низма. Усиление вентиляции легких при физической нагрузке на чинается раньше увеличения потребления кислорода и накопления угольной кислоты в крови человека. Это осуществляется благода ря афферентной импульсации от проприорецепторов активно ра ботающих мышц. Следовательно, импульсация проприорецепторов выступает как фактор, организующий перестройку работы функ циональной системы, поддерживающей оптимальный для метабо лизма уровень Р02, Рсс,2 и рН внутренней среды с опережением. Регуляция по опережению может реализоваться с помощью механизма условного рефлекса.
20. Безусловные рефлексы: определение, классификация Безусловные рефлексы — наследственно передаваемые (врождённые) реакции организма, присущие всему виду. Классификация: 1. По биологическому значению (оборонительные, половые, пищевые) 2. По расположению рецепторов в составе афферентного звена (Экстероцептивные, интро, проприо) 3. По модальности адекватного раздражителя (Механо-, хемо-, баро-, фото-, термо-) 4. По местоположению нервных центров (спинальные, бульбарные, метэнцефалические, кортикальные, диэнцефалические) 5. По эфферентному звену рефл дуги (соматические, вегетативные) Безусловные рефлексы (Видовые) – относительно постоянные стереотипные, вырожденные, генетически закрепленные реакции организма на внутренние и внешние раздражители, осуществляемые при участии ЦНС
21. Схема рефлекторной дуги соматического рефлекса. Рефлекторное кольцо. Рефлекторное кольцо – совокупность структур НС, участвует в осуществлении рефлекса и обратной передачи информации о характере и силе рефлекторного действия в ЦНС

22. Принципы рефлекторной теории Сеченова - Павлова. Динамический стереотип. 1. Принцип детерминизма (причины) – обусловленность всех поступков и действий, концепция «свободы воли». 2. Принцип структурности – определенная структура нервной системы выполняет свою конкретную функцию 3. Принцип синтеза и анализа раздражителя – анализ центральными структурами поступающих сигналов и синтез целостного образа сигнального раздражителя. В периферии – элементарный анализ и синтез (без участия ЦНС). Динамический стереотип -это закрепившаяся система временных нервных связей (условных рефлексов), образовавшаяся в ответ на устойчиво повторяющуюся систему одних и тех же условных раздражителей и позволяющая выполнять определенные действия без значительного напряжения сознания. Стереотип раздражителей – комплекс стимулов в строго определенном порядке пространства-времени, определяет характерные нормы поведения Стереотип комплексных условных рефлексов – стандартный признак сигнальных стимулов, вследствие чего их различают по качеству и силе, 3 свойства: 1. При его формировании – стремление к автоматизму(ходьба) 2. Облегчение усвоения новых навыков при присутствии стандартного признака 3. Адекватная реакция на новую обстановку при схожей основе
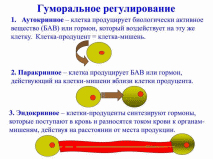
23. Гуморальная регуляция функций. Факторы гуморальной регуляции. Единство нервной и гуморальной регуляции Гуморальная регуляция это совокупность физических механизмов, изменение состояния отдельных клеток, тканей и систем посредством химических соединений, придающие информацию через внутреннюю среду Гуморальная регуляция – регуляция ж/д организма, осуществляемая при участии гормонов (Рег обменных процессов, гомеостазиса, роста, развития) Гормон – БАВ, вырабатываемый специализированными эндокринными клетками Эндокринные железы – не имеют выводных протоков – инкрет экзоцитозом в межклеточное пространство Гормон нервных клеток – нейрогормон Гипоталамус (либерины и статины) Тканевые гормоны (паракринные гормоны) – выделяются неспециализированными клетками Действие на соседние клетки – паракринная регуляция На рецептор, выделивший их же – аутокринная регуляция Действие гормона специфично (мишень либеринов и статинов – гипофиз) Либерины активируют, статины – тормозят Факторы гуморальной регуляции: 1. Неорганические метаболиты и ионы (Кальций, Натрий, Водород) 2. Гормоны желез внутренней секреции (Инсулин, тироксин, итд) 3. Местные или тканевые гормоны (гистамин, серотонин, гормоны ЖКТ) 4. БАВ, регулирующие дифференцировку, рост и развитие клеток Настоящие гормоны – выделяются специализированными эндокринными железами (инсулин, либерины, статины) Тканевые гормоны – неспециализированными тканями Метаболиты – вещества, выделяемые клетками в процессе обмена веществ (СО2, лактат, пируват) Гуморальная регуляция: местная (электролиты, метаболиты, креаторные факторы - межклеточные сигнальные макромолекулы, тканевые гормоны - бав Гормональная Креаторные факторы - кейлоны- Тканиспецифичные Ингибиторы пролиферации и дифференцировки клеток, способствующие подавлению деления клеток и синтеза днк Цитокины - Пептиды малой массы, регулирующие межклеточные и межсистемной взаимодействия: определяют выживаемость клеток, стимуляция или подавление роста, дифференциацию, активность и апоптоз
В организме высших животных и человека в результате длительной эволюции сложилась единая нейрогуморальная система регуляции функций. Деление этой системы на нервную и гуморальную - условное, оно необходимо для анализа сложнейших процессов, управляющих живым организмом. Ведущую роль в регуляторных реакциях играет нервная система и ее высший отдел - кора больших полушарий головного мозга. От многочисленных рецепторов, находящихся во всех органах и тканях сюда поступает, как в главный диспетчерский центр, информация о состоянии внешней среды и внутренней среды организма, обо всех изменениях в работе органов и систем, об изменениях в составе крови и тканевой жидкости. Однако, гуморальные агенты играют роль не только раздражителей, они могут включаться в рефлекторные дуги как самостоятельные звенья. Такое место, например, занимают гормоны. Допустим, в крови животного увеличился уровень глюкозы. Это вызывает возбуждение рецепторов сосудов (хеморецепторов), информация поступает в гипоталамус - отдел промежуточного мозга, и в кору больших полушарий. После оценки ситуации из гипоталамуса возбуждение передается в островковый аппарат поджелудочной железы, где вырабатывается гормон инсулин. Инсулин выделяется в кровь, действует на клеточные мембраны и снижает содержание глюкозы в крови до обычного уровня. Таким образом, гормон включился в эфферентную часть рефлекторной дуги. Однако единство нервной и гуморальной регуляции этим не ограничивается. Известно, что нервные клетки (нейроны) обладают двумя функциями: способностью генерировать биотоки и передавать их на другие клетки, и способностью вырабатывать биологически активные вещества. О том, что в нервных окончаниях выделяются химические вещества - медиаторы - было известно еще в 20-х годах XX века. Медиаторы являются химическими посредниками между эфферентными нервами и органами. Никогда нервные окончания не проникают внутрь другой клетки. Они заканчиваются на небольшом расстоянии от мембраны иннервируемой клетки. Место контакта нервного окончания с другой клеткой - нервной, мышечной или секреторной - называется синапсом. Синапс состоит из трех элементов: пресинаптической мембраны (часть нервного окончания), постсинаптической мембраны (часть мембраны другой клетки) и синаптической щели (пространство между пре- и постсинаптической мембранами). Передача возбуждения с нерва на орган (клетку) заключается в том, что под влиянием нервного импульса из пресинаптической мембраны выделяется медиатор, который является химическим раздражителем для постсинаптической мембраны. В результате в ней возникает возбуждение, распространяющееся по всей клетке. Самыми распространенными медиаторами являются ацетилхолин, норадреналин, адреналин и др. Помимо медиаторов, нейроны головного мозга вырабатывают гормоны - нейропептиды. Таковы, например, гормоны гипоталамуса. Их называют релизинг-факторами. Эти гормоны стимулируют или угнетают образование гормонов в передней доле гипофиза, которые, в свою очередь, регулируют деятельность других желез внутренней секреции. Некоторые гормоны гипоталамуса поступают в заднюю долю гипофиза уже в «готовом виде», а затем уже из гипофиза поступают в кровь. Таким образом, единство нервной и гуморальной регуляции осуществляется на уровне не только нервных окончаний, но и на более высоком уровне - гипоталамо-гипофизарной системы. Итак, регуляция всех жизненных процессов осуществляется единой нейрогуморальной системой, в которой ведущее значение играет центральная нервная система и ее высший отдел - кора больших полушарий. На более простых уровнях организации живой материи - молекулярном, внутриклеточном, тканевом - большую роль играет химическая регуляция. Чем сложнее биологическая система, тем большее значение приобретает нервная регуляция, которая объединяет органы и ткани в различные системы и осуществляет функционирование организма во внешней среде как единого целого.
24. Местные механизмы регуляции функций организма. Местная регуляция - регуляция в пределах одной ткани или органа. Осуществляется: 1)креаторными связями; 2)метаболитами (молочная кислота, пировиноградная кислота, аденозин); 3)ионами; 4)БАВ. Креаторные связи – обмен макромолекулами между клетками. Регулируют внутриклеточные обменные процессы, синтез белка, объединяют клетки в ткань, обеспечивают дифференцировку, рост и развитие клеток как единой ткани. Такими веществами являются: кейлоны – простые белки или гликопротеиды, подавляют деление клеток и синтез ДНК (отсутствие опухоли). Метаболиты обеспечивают регуляцию по принципу обратной связи на уровне функционального элемента ткани. Например, закисление среды кислыми метаболитами расширяет МЦР, обеспечивая больший приток крови, но ухудшаются сократительные способности мышц. Регуляторные эффекты метаболитов неспецифичны. БАВ или «тканевые гормоны» (гистамин; серотонин; кинины; простогландины): 1)изменяют проницаемость мембраны, величину мембранного потенциала, обмен веществ и энергии, чувствительность клеточных рецепторов, скорость образования вторичных посредников, просвет сосудов; 2)изменяют чувствительность клеток к нервным и гуморальным влияниям - модифицируют ответ на регуляторные воздействия.


25. Нейрон как структурно-функциональная единица нервной системы. Классификация нейронов. Свойства и функции нейронов. Структурной и функциональной единицей нервной системы является нервная клетка — нейрон. НЕЙРОНЫ — специализированные клетки, способные принимать, обрабатывать, кодировать, передавать и хранить информацию, организовывать реакции на раздражения, устанавливать контакты с другими нейронами, клетками органов. Функционально в нейроне выделяют следующие части: ВОСПРИНИМАЮЩУЮ — дендриты, мембрана сомы нейрона; ИНТЕГРАТИВНУЮ — сома с аксонным холмиком; ПЕРЕДАЮЩУЮ — аксонный холмик с аксоном. Сома обеспечивает также рост дендритов и аксона. Сома нейрона заключена в многослойную мембрану, обеспечивающую формирование и распространение электротонического потенциала к аксонному холмику. ВОСПРИНИМАЮЩАЯ ЧАСТЬ. ДЕНДРИТЫ — основное воспринимающее поле нейрона. Мембрана дендрита и синаптической части тела клетки способна реагировать на медиаторы, выделяемые аксонными окончаниями изменением электрического потенциала. МЕМБРАНА СОМЫ НЕЙРОНА имеет толщину 6 нм и состоит из двух слоев липидных молекул. Гидрофильные концы этих молекул обращены в сторону водной фазы: один слой молекул обращен внутрь, другой – наружу. Гидрофильные концы повернуты друг к другу – внутрь мембраны. ИНТЕГРАТИВНАЯ ЧАСТЬ. АКСОНОВЫЙ ХОЛМИК – место выхода аксона из нейрона. СОМА НЕЙРОНА (тело нейрона) выполняет наряду с информационной и трофическую функцию относительно своих отростков и синапсов. Сома обеспечивает рост дендритов и аксонов. Сома нейрона заключена в многослойную мембрану, которая обеспечивает формирование и распространение электротонического потенциала к аксонному холмику. ПЕРЕДАЮЩАЯ ЧАСТЬ. АКСОН представляет собой вырост цитоплазмы, приспособленный для проведения информации, собранной дендритами, переработанной в нейроне и переданной аксону через аксонный холмик — место выхода аксона из нейрона. Аксон данной клетки имеет постоянный диаметр. Аксон имеет разветвленные окончания. Нейроны классифицируются по нескольким признакам: по форме тела – звездчатые, веретенообразные, пирамидные и др по локализации – центральные (расположены в ЦНС) и периферические (расположены вне ЦНС, а в спинномозговых, черепно-мозговых и вегетативных ганглиях, сплетениях, внутри органов) по числу отростков – униполярные, биполярные и мультиполярные (рис. 3.3.2) по функциональному признаку – рецепторные, эфферентные, вставочные Физиологические свойства нейронов : 1. Раздражимость – способность клетки реагировать изменением обмена в-в на действие раздражителя. 2. Возбудимость – способность клетки реагировать на раздражение возбуждением. 3. Проводимость – способность клетки к распространению возбуждения вдоль своей мембраны. 4. Лабильность – способность клетки воспроизводить без искажения максимальную частоту возбуждения. 5. Рефрактерность – способность клетки в течение некоторого времени не возбуждаться Функции нейронов: 1. Восприятие информации – за счёт дендритов и мембраны тела клеток ( сомы ) 2. Кодирование информации – за счёт сомы и аксонного холмика 3. Интегративная функция – сома и аксонный холмик 4. Хранение информации 5. Передача информации за счёт аксона и аксонного холмика .
26.классификация нервных волокон Миелиновые волокна типа А – 4 подгруппы (альфа, бэта, гамма, дельта) Тип А-альфа – наиболее толстые (12-22 мкм), быстрая скорость проведения сигнала (70-120 м/с), длительность ПД 0,4-0,5 мс. Проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам и от рецепторов к нервным волокнам. (Двигательные волокна скелетных мышц) Тип А-бэта – диаметр волокна 8-12 мкм, скорость проведения импульса 40-70 м/с, длительность ПД 0,4-0,6 мс. Это афферентные волокна от тактильных рецепторов (давление, прикосновение). Тип А-гамма – диаметр волокна 4-8 мкм, скорость проведения импульса 15-40 м/с, длительность ПД 0,5-0,7 мс. Это афферентные волокна от тактильных рецепторов и барорецепторов, эфферентные волокна к мышечным веретенам. Тип А-дельта – диаметр волокна 1-4 мкм, скорость проведения импульса 5-15 м/с, длительность ПД 0,6-1 мс. Это афферентные волокна от терморецепторов, барорецепторов, болевых рецепторов. Тип В – миелиновые, преганглионарные волокна автономной НС. Скорость проведения 3-18 м/с, диаметр волокна 1-3,5 мкм, продолжительность ПД 1-2 мс. Безмиелиновые волокна тип С – очень малый диаметр (0,5-2 мкм), скорость проведения 0,5-3 м/с. Постганглионарные волокна симпатической НС, афферентные волокна от термо-, баро-, и болевых рецепторов. Длительность ПД 2 мс.
27. Механизмы и законы проведения возбуждения по нервным волокнам. Механизм проведения возбуждения по нервным волокнам: 1) По безмиелиновому a) Деполяризация мембраны нервного волокна b) Возникновение продольных токов (направленные в обе стороны от активного участка генерации ПД) c) Продольные токи вызывают открытие потенциал-зависимых натрий ионных каналов в участке мембраны, который имеет нормальную возбудимость d) Генерация ПД в этом участке e) Рефрактерный участок невозбудим (натриевые ионные каналы находятся в инактивированном состоянии) 2) По миелиновому a) Деполяризация мембраны нервного волокна в перехвате Ранвье b) Возникновение продольных токов (направленные в обе стороны от активного участка генерации ПД) c) Продольные токи вызывают открытие потенциал-зависимых натрий ионных каналов в следующих перехватах Ранвье d) Генерация ПД в этом перехвате (минуя миелинизированный участок нервного волокна – сальтоторно) e) Предыдущий перехват Ранвье невозбудим (натриевые ионные каналы находятся в инактивированном состоянии) Законы проведения возбуждения по нервным волокнам: 1. Закон анатомической и физиологической целостности нерва. Первая нарушается при повреждении, 2 – при блокаторах. 2. Закон 2х стороннего возбуждения – распространение в обе стороны от места раздражения 3. Закон одностороннего проведения – передача ПД от тела нейрона к окончаниям (центробежно) или наоборот (центростремительно) 4. Закон изолированного проведения. Возбуждение не передается с одного волокна на другое, входящего в состав нерва. Обеспечивает изолированное сокращение каждой Нейро моторной единицы. 5.Скорость проведения прямопропорциональна диаметра нерва
28. Синапс: определение, классификация. Механизмы (этапы) синаптической передачи. Синапсы – специализированная структура, обеспечивающая передачу нервного импульса из нервного волокна на эффекторную клетку (мыш волокно, нейрон, секреторная клетка) Синапс – место соединения нервного отростка одного нейрона с телом или отростком другой нервной клетки Виды синапсов: По расположению: Аксо-дендрические, аксосоматические, аксо-аксональные, аксо-шипиковые, дендрито-дендритные, сомато-соматические По способу передачи: Химические (медиаторы), электрические (ионы), смешанные По анатомо-гистологическому принципу: Нейросекреторные, нервно-мышечные, межнейронные. По нейрохимическому принципу: Адренергические, холинегрические итд По функциональному признаку: Возбуждающие, тормозные. Строение синапса: Пресинаптич мембр, синаптич щель, постсинаптич мембр Гипотеза о химической синаптической передаче с нерва на мышцу была предложена А.Ф. Самойловым в 1924 году. В хим синапсе щель – 20 нм. Механизм передачи ПД по нексусу: 1. Электрическая синаптическая передача возможна при очень тесном соприкосновении взаимодействующих клеток (не более 10-20 нм) 2. Переход возбуждения с клетки на клетку посредством локальных токов 3. Ретрансляция ПД на мембране второй клетки (щелевые контакты – нексусы) 4. Электрическое сопротивление нексусов регулируется посредством изменения содержания ионов кальция и водорода в цитозоле (увеличивается при повышении концентрации ионов кальция) 5. Скорость передачи значительно выше, чем в химическом 6. Происходит потеря амплитуды – декремент (нет механизмов ретрансляции) 7. Ненадежная межнейронная передача, не адресная 8. Не характерно одностороннее проведение сигнала Химические синапсы: 1. Возбуждающие (медиатор вызывает деполяризацию) 2. Тормозные (выз реполяризацию) Выделяют следующие этапы передачи ПД в химическом синапсе. 1. Под влиянием поступающих по пресинаптическому волокну нервных импульсов происходит деполяризация пресинаптической мембраны. 2. Это сопровождается открыванием потенциалзависимых кальциевых каналов и входом кальция в пресинаптическое окончание. Кальций связывается с белком кальмодулином и стимулируют процессы фосфорилирования белка синапсина, входящего в состав мембраны везикул; в результате происходит слияние мембраны синаптических пузырьков с синаптической мембраной, то есть экзоцитоз. 3. Выделившиеся кванты медиатора диффундируют через синаптическую щель к постсинаптической мембране, где связываются со специальными клеточными рецепторами, специфичными для каждого медиатора. 4. Образовавшейся на постсинаптической мембране комплекс «медиатор – рецептор» активирует хемочувствительные мембранные каналы, что повышает проницаемость мембраны для ионов и меняет ее потенциал покоя. Возбуждающие медиаторы вызывают появление на постсинаптической мембране возбуждающего постсинаптического потенциала (ВПСП). Его формирование обусловлено тем, что комплекс «медиатор – рецептор» активирует ионные каналы мембраны, прежде всего натриевые, а также кальциевые и вызывает деполяризацию мембраны за счет поступления натрия или кальция внутрь клетки. Одновременно происходит и уменьшение выхода из клетки ионов калия. Амплитуда одиночного ВПСП довольно мала, от 0,12 до 5 мВ и для уменьшения заряда мембраны до критического уровня необходима одновременная активация нескольких возбуждающих синапсов. ВПСП, образующиеся на постсинаптической мембране этих синапсов, способны суммироваться, приводя к росту амплитуды ВПСП. Процесс спонтанной деполяризации обычно возникает в аксонном холмике, где аксон еще не покрыт миелином и порог возбуждения низкий. Таким образом, ВПСП, возникающие в разных участках мембраны нейрона и на его дендритах, распространяются к аксонному холмику, где суммируются, деполяризуя мембрану до критического уровня и приводя к появлению потенциалов действия. Отметим, что в химическом синапсе электрический процесс – генерация ПД – полностью угасает и переходит в химический процесс – взаимодействие медиатора с рецептором, в результате которого ПД возникает на постсинаптической мембране.

29. Передача сигнала в возбуждающих и тормозных синапсах
При возбуждениинейрона возникаетпотенциал действия (ПД), который подходит к окончанию аксона и ионы кальция начинают поступать из внеклеточной жидкости внутрь нервного окончания. Кальций активирует перемещение синаптических пузырьков к пресинаптической мембране, где они разрушаются с выходом медиатора в синаптическую щель. В возбуждающих синапсах медиатор диффундирует в щели и связывается с рецепторами постсинаптической мембраны, что приводит к открытию каналов для ионов натрия, а следовательно, к ее деполяризации — возникновению возбуждающего постсинаптического потенциала (ВПСП). Между деполяризованной мембраной и соседними с ней участками возникают местные токи. Если они деполяризуют мембрану до критического уровня, то в ней возникает потенциал действия. Для восстановления возбудимости постсинаптической мембраны после очередного импульса необходима инактивация медиатора. В противном случае, при длительном действии медиатора происходит снижение чувствительности рецепторов к этому медиатору.
В тормозных синапсах медиатор (например, глицин) аналогичным образом взаимодействует с рецепторами постсинаптической мембраны, но открывает в ней калиевые и/или хлорные каналы, что вызывает переход ионов по концентрационному градиенту: калия из клетки, а хлора — внутрь клетки. Это приводит к гиперполяризации постсинаптической мембраны — возникновению тормозного постсинаптического потенциала (ТПСП). Тормозной эффект ТПСП основывается на двух механизмах. Во-первых, это электротоническое действие гиперполяризационного ТПСП на триггерную (аксонный холмик) зону нейрона: ТПСП порождает ток, который входит в холмик и повышает его мембранный потенциал.
Во-вторых, имеет значение действие хлорного шунта на ВПСП. Открытие хлорных каналов закорачивает ток ВПСП и уменьшает плотность тока, протекающего через триггерную зону нейрона. Хлорный механизм торможения существует в нейронах ЦНС наряду с механизмом активации калиевых каналов и повышением калиевой проницаемости.
30. Нейронные цепи и нейронные сети: определение. Взаимодействие нейронов в нервных цепях и нейронных сетях.
???? Нейронные цепи – это соответствующим образом, чаще всего последовательно, соединенные между собой нейроны, которые выполняют определенную задачу.
Нейронные сети – это объединение нейронов, которое содержит множество параллельно расположенных и связанных между собой последовательных цепей нейронов. Такие объединения выполняют сложные задачи, участвующих в получении, передаче, хранении и воспроизведении информации.
Нервная система человека может быть представлена в виде нервной сети, т.е. системы нейронных цепочек, передающих возбуждающие и тормозные сигналы. Нейронные сети построены из трех главных компонентов: входных волокон, вставочных нейронов и эфферентных нейронов. Самыми простыми и элементарными нейронными цепями являются локальные сети, или микросети
Локальные сети имеются в разных отделах мозга. Они служат: 1) для усиления слабых сигналов; 2) уменьшения и фильтрации слишком интенсивной активности; 3) выделения контрастов; 4) поддержания ритмов или сохранения рабочего состояния нейронов путем регулировки их входов. Микросети могут оказывать на нейроны-мишени возбуждающее или тормозящее действие.
31. Нервный центр: определение, структура, функциональные свойств Нервный центр (по Павлову) – совокупность нейронов, связанных между собой синапсами, расположенных в одном или нескольких отделах ЦНС и объединенных в систему обеспечением определённых функций организма. Любой нервный центр состоит из ядра и периферии. Ядерная часть нервного центра представляет собой функциональное объединение нейронов, в которое поступает основная информация от афферентных путей. Периферическая часть нервного центра получает небольшую порцию афферентной информации, и ее повреждение вызывает ограничение или уменьшение объема выполняемой функции. Временная суммация возникает как в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда возбуждающего постсинаптического потенциала. Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторных нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются, и в мембране нейрона генерируется распространяющийся потенциал действия. Лекция Слабые раздражители одного или нескольких участков рецептивного поля в отдельности не способны реализовать рефлекс, но, действуя на нервный центр одновременно, они генерируют ВПСП, которые суммируются, приводя к формированию на мембране нервных клеток потенциалов действия, распространяющихся по эфферентным проводникам, вызывая рефлекторную реакцию. Это явление называется пространственной суммацией. При увеличении частоты подпороговых афферентных сигналов, приходящих к одному и тому же входному нейрону, суммация ВПСП на постсинаптической мембране приводит к достижению КУД, что также вызывает возбуждение нервного центра и возникновение рефлекторной реакции на слабые, но частые стимулы. Это явление временной суммации 1. Одностороннее проведение возбуждения в НЦ- через нервные центры нервные импульсы передаются с афферентных волокон на эфферентные. 2. Центральная задержка – более медленное проведение в НЦ из-за передачи медиаторов, чем в волокнах 3. Посттетаническая потенциация – становление подпороговых раздражителей надпорговыми после высокочастотных стимуляций; усиление рефлекторного ответа под влиянием предшествующих частых ритмических (т. н. тетанизирующих) раз-дражений. 4.Последействие - сохранение возбуждения в нервном центре после действия раздражителя. 5.Трансформация ритма возбуждения – изменение активности после прохождения возбуждения через НЦ 6.Тонус НЦ – постоянное поддержание электрической активности нейронов 7. Утомляемость НЦ – снижение функции при чрезмерной работе
32. Механизмы взаимодействия (интеграции) нервных центров: реципрокность, общий конечный путь, субординация, обратная афферентация, доминанта, индукция. Механизм взаимодействия НЦ были изучены Введенским, Ухтомским 1. Принцип РЕЦИПРОКНОСТИ (сопряженное торможение) – многие НЦ связаны друг с другом посредством тормозных нейронов. Возбуждение одних – тормоз других (сгибатели-разгибатели) 2. Принцип ОБЩЕГО КОНЕЧНОГО ПУТИ – ( мотонейроны спинного мозга являются общим конечным путём многочисленных рефлексов) на одном Альфа-мотонейроне конвергируют множество нисходящих путей, выходит всего 1 аксон, нисходящие пути борются за этот аксон 3. ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ – в результате открытия проприоцепторов. любой эффектор рефлекторной реакции имеет рецепторы, сигнализирующие об исполнении команды. 4. ПРИНЦИП СУБОРДИНАЦИИ НЦ – иерархия взаимоотношений между разными центрами (вышележащие НЦ подчиняют себе нижележащие) 5. ПРИНЦИП ДОМИНАНТЫ – устанавливает соответствие нервной деятельности с ведущими потребностями организма в данный момент. Основные черты доминант: доминирующий нервный центр обладает повышенной возбудимостью среди всех НЦ разных уровней, доминирующий НЦ имеет стойкое возбуждение, тормозит НЦ несовместимые с его деятельностью, домин НЦ инертен, притягивает нервные импульсы от информационных каналов, не имеющих с ним прямой связи 6. Индукция (от лат. inductio — побуждение) — это свойство нервных процессов возбуждения и торможения вызывать после себя противоположный процесс. Во время движения нервных процессов происходит взаимовлияние возбуждения и торможения, где торможение вызывает (индуцирует) возбуждение (положительная индукция), а процесс возбуждения индуцирует торможение (отрицательная индукция).
33.
Торможение в центральной нервной
системе: определение. Классификация
торможения по локализации (пре- и
постсинаптическое), нейрональной
организации (первичное и вторичное) и
форме (возвратное, латеральное,
реципрокное).
Торможение –
активный нервный процесс, проявл в
подавлении возникновения и распространение
возбуждения и его остановки.
По
локализации:
