

Вопросы для подготовки к экзамену
Вопросы из программы осеннего семестра
Строение, функции клеточных мембран и ионных каналов клеточной мембраны. Виды активного и пассивного транспорта веществ через клеточную мембрану.
Клеточная мембрана (цитолемма, плазмалемма) – это трехслойная липопротеиновая (жиро-белковая) оболочка, отделяющая каждую клетку от соседних клеток и окружающей среды, и осуществляющая управляемый обмен между клетками и окружающей средой.
В составе клеточных мембран присутствуют белки трех видов:
Периферические – крепятся на поверхности пленки;
Полуинтегральные – частично проникают внутрь билипидного слоя;
Интегральные – полностью пронизывают мембрану.
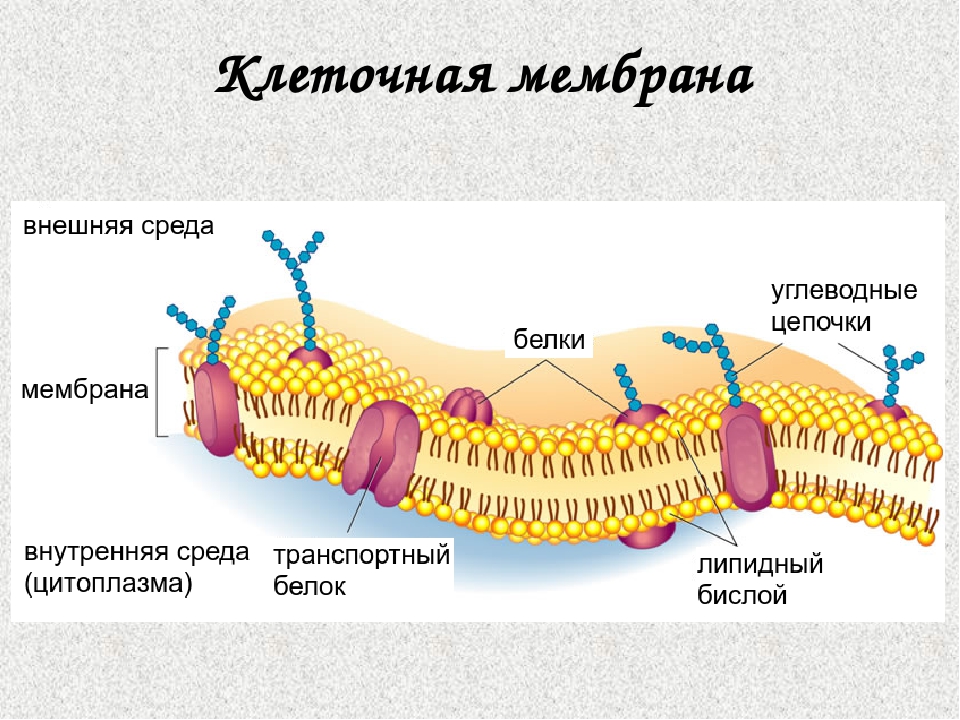
Периферические белки связаны с головками мембранных липидов посредством электростатического взаимодействия, и они никогда не образуют сплошной слой, как принято было считать ранее.А полуинтегральные и интегральные белки служат для транспортировки внутрь клетки кислорода и питательных веществ, а также для вывода из нее продуктов распада
Клеточная мембрана выполняет следующие функции:
Барьерную – проницаемость мембраны для разных типов молекул неодинакова.Чтобы миновать оболочку клетки, молекула должна иметь определенный размер, химические свойства и электрический заряд. Вредные или неподходящие молекулы, благодаря барьерной функции клеточной мембраны, просто не могут проникнуть внутрь клетки;
Транспортную – сквозь мембрану проходит пассивный, активный, регулируемый и избирательный обмен. Пассивный обмен подходит для жирорастворимых веществ и газов, состоящих из очень маленьких молекул. Такие вещества проникают внутрь и выходят из клетки без затрат энергии, свободно, методом диффузии. Активная транспортная функция клеточной мембраны задействуется тогда, когда в клетку или из нее нужно провести необходимые, но трудно транспортируемые вещества. Например, обладающие большим размером молекул, или неспособные пересечь билипидный слой из-за гидрофобности. Тогда начинают работать белки-насосы, в том числе АТФаза, которая отвечает за всасывание в клетку ионов калия и выбрасывание из нее ионов натрия. Регулируемый транспортный обмен необходим для осуществления функций секреции и ферментации, например, когда клетки производят и выделяют гормоны или желудочный сок. Все эти вещества выходят из клеток через специальные каналы и в заданном объеме. А избирательная транспортная функция связана с теми самыми интегральными белками, которые пронизывают мембрану и служат каналом для входа и выхода строго определенных типов молекул;
Матричную – клеточная мембрана определяет и фиксирует расположение органоидов относительно друг друга (ядра, митохондрий, хлоропластов) и регулирует взаимодействие между ними;
Механическую – обеспечивает ограничение одной клетки от другой, и, в то же время,— правильное соединение клеток в однородную ткань и устойчивость органов к деформации;
Защитную – как у растений, так и у животных, клеточная мембрана служит основой для построения защитного каркаса;
Энергетическую — процессы фотосинтеза и клеточного дыхания были бы невозможны без участия белков клеточной мембраны, ведь именно с помощью белковых каналов клетки обмениваются энергией;
Рецепторную— белки, встроенные в клеточную мембрану, могут обладать ещё одной важной функцией. Они служат рецепторами, благодаря которым клетка получает сигнал от гормонов и нейромедиаторов. А это, в свою очередь, необходимо для проведения нервных импульсов и нормального течения гормональных процессов;
Ферментативную — ещё одна важная функция, присущая некоторым белкам клеточных мембран. Например, в эпителии кишечника с помощью таких белков синтезируются пищеварительные ферменты;
Биопотенциальную – концентрация ионов калия внутри клетки значительно выше, чем снаружи, а концентрация ионов натрия, наоборот, снаружи больше, чем внутри. Этим и объясняется разность потенциалов: внутри клетки заряд отрицательный, в снаружи положительный, что способствует движению веществ внутрь клетки и наружу при любом из трех типов обмена – фагоцитозе, пиноцитозе и экзоцитозе;
Маркировочную – на поверхности клеточных мембран имеются так называемые «ярлыки» - антигены, состоящие из гликопротеинов (белков с присоединенными к ним разветвленными олигосахаридными боковыми цепями). Поскольку боковые цепи могут иметь огромное множество конфигураций, каждый тип клеток получает свой уникальный ярлык, который позволяет другим клеткам организма узнавать их «в лицо» и правильно на них реагировать. Вот почему, например, иммунные клетки человека, макрофаги, без труда распознают чужака, проникшего в организм (инфекцию, вирус) и пытаются его уничтожить. То же самое происходит с больными, мутировавшими и старыми клетками – ярлык на их клеточной мембране меняется, и организм избавляется от них.
Строение и функции ионных каналов. Ионы Na+, K+, Са2+, Сl- проникают внутрь клетки и выходят наружу через специальные, заполненные жидкостью каналы. Размер каналов довольно мал.
Все ионные каналы подразделяются на следующие группы:
По избирательности:
a) Селективные, т.е. специфические. Эти каналы проницаемы для строго определенных ионов.
b) Малоселективные, неспецифические, не имеющие определенной ионной избирательности. Их в мембране небольшое количество.
По характеру пропускаемых ионов:
a) калиевые
b) натриевые
c) кальцевые
d) хлорные
По скорости инактивации, т.е. закрывания:
a) быстроинактивирующиеся, т.е. быстро переходящие в закрытое состояние. Они обеспечивают быстро нарастающее снижение МП и такое же быстрое восстановление.
b) медленноинактирующиеся. Их открывание вызывает медленное снижение МП и медленное его восстановление.
4. По механизмам открывания:
a) потенциалзависимые, т.е. те которые открываются при определенном уровне потенциала мембраны.
b) хемозависимые, открывающиеся при воздействии на хеморецепторы мембраны клетки физиологически активных веществ (нейромедиаторов, гормонов и т. д).
В настоящее время установлено, что ионные каналы имеют следующее строение:
1.Селективный фильтр, расположенный в устье канала. Он обеспечивает прохождение через канал строго определенных ионов.
2.Активационные ворота, которые открываются при определенном уровне мембранного потенциала. Активационные ворота потенциалзависимых каналов имеется сенсор, который открывает их на определенном уровне МП.
3.Инактивационные ворота, обеспечивающие закрывание канала и прекращение проведения ионов по каналу на определенном уровне МП.
Неспецифические ионные каналы не имеют ворот.
Селективные ионные каналы могут находиться в трех состояниях, которые определяются положением активационных (м) и инактивационных (h) ворот:
1.Закрытом, когда активационные закрыты, а инактивационные открыты.
2.Активированном, и те и другие ворота открыты.
3.Инактивированном, активационные ворота открыты, а инактивационные закрыты
Функции ионных каналов:
1. Калиевый (в покое) – генерация потенциала покоя
2. Натриевый – генерация потенциала действия
3. Кальциевый - генерация медленных действий
4. Калиевый (задержанное выпрямление) – обеспечение реполяризации
5. Калиевый кальций-активируемый – ограничение деполяризации, обусловленной током Са+2
Пассивный транспорт– транспорт веществ через мембрану, осущестляемый без затрат энергии.
1. Простая диффузия. Небольшие нейтральные молекулы (Н2О, СО2, О2, NH3, мочевина, этанол) диффундируют через мембрану без участия специальных механизмов. Перенос веществ осуществляется по градиенту концентрации и с низкой скоростью.
2. Облегченная диффузия. Для более крупных полярных молекул (глюкоза, аминокислоты), а также для ионов липидный бислой практически непроницаем, так как его внутренняя часть гидрофобна. Такие вещества переносятся через мембрану также по градиенту концентрации, но с участием мембранных белков.
Активный транспорт
Активный транспорт веществ протекает против концентрационного градиента и требует затрат энергии. Таким способом происходит перенос многих минеральных ионов из межклеточной жидкости в клетку или в обратном направлении, перенос аминокислот из просвета кишечника в клетки кишечника, перенос глюкозы из первичной мочи через клетки канальцев почки в кровь. Основным источником энергии для активного транспорта является АТФ. Поэтому, как правило, эти системы представляют собой АТФазы.
В зависимости от количества веществ, переносимых через один белок-переносчик, и направления транспорта, различают:
унипорт - транспорт одного вещества;
симпорт - транспорт двух веществ в одном направлении через один переносчик;
антипорт - перемещение двух веществ в разных направлениях через один переносчик.
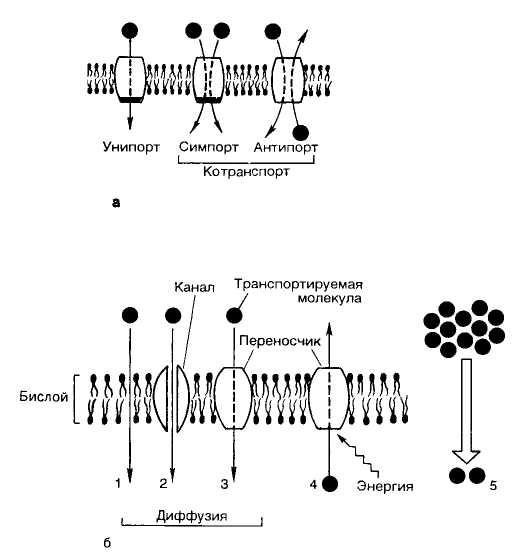
Эта классификация справедлива как для пассивного, так и для активного транспорта. Примером пассивного антипорта является перенос ионов хлора и гидрокарбонат-ионов.
Примером активного антипорта яляется натрий–калиевая АТФаза. Она переносит в клетку ионы калия, а из клетки - ионы натрия.
АТФ-аза присоединяет с внутренней стороны мембраны три иона Na+. Эти ионы изменяют конформацию активного центра АТФазы, и она гидролизует одну молекулу АТФ, присоединяя к себе фосфат. Выделившаяся энергия расходуется на изменение конформации АТФазы, после чего три иона натрия оказываются на внешней стороне мембраны, а фосфат замещается на 2 иона K+ из внешней среды. Затем конформация переносчика изменяется на первоначальную, и ионы K+ оказываются на внутренней стороне мембраны. Здесь ионы K+ отщепляются.
Работа Na+,K+-ATФазы создает не только разность концентраций ионов, но и потенциал на мембране. На внешней стороне мембраны создается положительный заряд, на внутренней - отрицательный.
Вторично-активный транспорт. Градиент одного вещества используется для транспорта другого. Переносчик в этом случае имеет специфические центры связывания для обоих веществ. Вещество транспортируется против градиента своей концентрации путем симпорта или антипорта. Симпорт и антипорт, например, могут происходить за счет энергии градиента концентрации ионов Na+, создаваемого Na+,K+-ATФазой. Таким способом происходит всасывание аминокислот из кишечника и глюкозы из первичной мочи икишечника.
Пример вторично-активного симпорта – транспорт глюкозы и ионов натрия; вторично-активного антипорта – транспорт ионов кальция и натрия.
Для переноса углеводов, аминокислот и других метаболитов вторично-активный транспорт имеет, по-видимому, наибольшее значение по сравнению с другими механизмами.
Структура и функции мембран нарушаются при ряде заболеваний.
Потенциал покоя, его происхождение и ионные механизмы. Фазы потенциала действия, их происхождение.
Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин:
1) неодинакового распределения ионов по обе стороны мембраны. Внутри клетки находится больше всего ионов К, снаружи его мало. Ионов Na и ионов Cl больше снаружи, чем внутри. Такое распределение ионов называется ионной асимметрией;
2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ.
За счет этих двух факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузией в результате разности концентрации ионов. Ионы K выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны, ионы Cl пассивно переходят внутрь клетки, что приводит к увеличению положительного заряда на наружной поверхности клетки. Ионы Na накапливаются на наружной п оверхности мембраны и увеличивают ее положительный заряд. Органические соединения остаются внутри клетки. В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны может не быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточной мембраны называется состоянием поляризации. Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов на мембране, т. е. не наступит электрохимическое равновесие. Момент равновесия зависит от двух сил:
1) силы диффузии;
2) силы электростатического взаимодействия.
Значение электрохимического равновесия:
1) поддержание ионной асимметрии;
2) поддержание величины мембранного потенциала на постоянном уровне.
В возникновении мембранного потенциала участвуют сила диффузии (разность концентрации ионов) и сила электростатического взаимодействия, поэтому мембранный потенциал называется концентрационно-электрохимическим.
Для поддержания ионной асимметрии электрохимического равновесия недостаточно. В клетке имеется другой механизм – натрий-калиевый насос. Натрий-калиевый насос – механизм обеспечения активного транспорта ионов. В клеточной мембране имеется система переносчиков, каждый из которых связывает три иона Na, которые находятся внутри клетки, и выводит их наружу. С наружной стороны переносчик связывается с двумя ионами K, находящимися вне клетки, и переносит их в цитоплазму. Энергия берется при расщеплении АТФ. Работа натрий-калиевого насоса обеспечивает:
1) высокую концентрацию ионов К внутри клетки, т. е. постоянную величину потенциала покоя;
2) низкую концентрацию ионов Na внутри клетки, т. е. сохраняет нормальную осмолярность и объем клетки, создает базу для генерации потенциала действия;
3) стабильный концетрационный градиент ионов Na, способствуя транспорту аминокислот и сахаров.
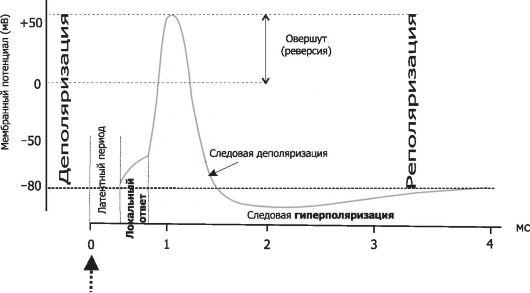
Фазы потенциала действия
Восходящая ветвь графика:
потенциал покоя – исходное обычное поляризованное электроотрицательное состояние мембраны (–70 мВ);
нарастающий локальный потенциал – пропорциональная раздражителю деполяризация;
критический уровень деполяризации (–50 мВ) – резкое ускорение деполяризации (за счет самораскрытия натриевых каналов), с этой точки начинается спайк – высокоамплитудная часть потенциала действия;
самоусиливающаяся круто нарастающая деполяризация;
переход нулевой отметки (0 мВ) – смена полярности мембраны;
«овершут» – положительная поляризация (инверсия, или реверсия, заряда мембраны);
пик (+30 мВ) – вершина процесса изменения полярности мембраны, вершина потенциала действия.
Нисходящая ветвь графика:
реполяризация – восстановление прежней электроотрицательности мембраны;
переход нулевой отметки (0 мВ) – обратная смена полярности мембраны на прежнюю, отрицательную;
переход критического уровня деполяризации (–50 мВ) – прекращение фазы относительной рефрактерности (невозбудимости) и возврат возбудимости;
следовые процессы (следовая деполяризация или следовая гиперполяризация);
восстановление потенциала покоя – норма (–70 мВ).
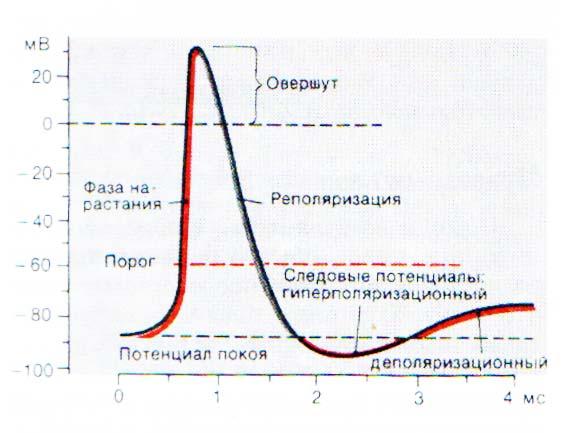
Фазовые изменения возбудимости клеток при генерации потенциала действия. Критерий возбудимости (порог раздражения, хронаксия, лабильность).
Потенциал действия – это скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно.
При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается в 400–500 раз, и градиент нарастает быстро, для ионов К – в 10–15 раз, и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный.
Возбуждение нервной клетки под действием химического сигнала (реже электрического импульса) приводит к возникновению потенциала действия.
Процесс начинается с открывания Nа+-канала (1). Ионы Na+устремляются в клетку (по градиенту концентрации), что вызывает локальное обращение знака мембранного потенциала (2). При этом Na+-каналы тотчас закрываются, т. е. поток ионов Na+ в клетку длится очень короткое время (3). В связи с изменением мембранного потенциала открываются (на несколько мс) потенциал-управляемые К+-каналы (2) и ионы К+ устремляются в обратном направлении, из клетки. В результате мембранный потенциал принимает первоначальное значение (3), и даже превышает на короткое время потенциал покоя (4). После этого нервная клетка вновь становится возбудимой.
За один импульс через мембрану проходит небольшая часть ионов Na+ и К+, и концентрационные градиенты обоих ионов сохраняются (в клетке выше уровень К+, а вне клетки выше уровень Na+). Поэтому по мере получения клеткой новых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно. Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращение мембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку.
К параметрам возбудимости относятся:
Порог раздражения - это минимальная сила раздражителя, при которой возникает возбуждение.
Реобаза - это минимальная сила раздражителя, вызывающая возбуждение при его действии в течение неограниченно долгого времени. На практике порог и реобаза имеют одинаковый смысл. Чем ниже порог раздражения или меньше реобаза, тем выше возбудимость ткани. Величина реобазы определяется разностью между критическим потенциалом и мембранным потенциалом покоя.
Полезное время - минимальное время действия раздражителя силой в одну реобазу, за которое возникает возбуждение. Порог времени определяется емкостной и резистивной характеристикой клеточной мембраны.
Хронаксия - это минимальное время действия раздражителя силой в две реобазы, необходимое для возникновения возбуждения. Использование этого критерия позволяет точно измерить временные характеристики возбудимых структур, поскольку измерение происходит на крутом изгибе гиперболы. Хронаксиметрия используется при оценке функционального состояния нервно-мышечной системы у человека. При ее органических поражениях величина хронаксии и реобазы нервов и мышц значительно возрастает.
Лабильность – способность возбудимой ткани реагировать на раздражение с определенной скоростью. Лабильность характеризуется максимальным числом волн возбуждения, возникающих в ткани в единицу времени (1 с) в точном соответствии с ритмом наносимых раздражений без явления трансформации. Лабильность определяется продолжительностью рефрактерного периода (чем короче рефрактерный период, тем больше лабильность).
Законы раздражения возбудимых тканей (силы, длительности, скорости нарастания раздражения). Законы действия постоянного тока на возбудимые ткани.
Закон силы раздражения – чем больше сила раздражителя, тем больше ответная реакция (но до определённого предела)
Принцип «все или ничего» (закон Боудича) частный случай. «Клетка не отвечает на подпороговый раздражитель, но на пороговые и сверхпороговые раздражители – возникает максимально возможная ответная реакция»
2) Закон длительности – чем больше длительность раздражения пороговой величины, тем больше ответная реакция (до определённого предела).
3) Закон нарастания силы раздражения – чем больше крутизна нарастания силы раздражения (скорость нарастания), тем больше ответная реакция, но до определенного предела. При снижении крутизны нарастания силы раздражителя возникает аккомодация (снижение возбудимости)
Законы действия посстоянного тока на возбудимые ткани
В 19 веке Пфлюгер установил, что при замыкании цепи постоянного тока, под отрицательным электродом, т.е. катодом возбудимость повышается, а под положительным – анодом, снижается. Это называется законом действия постоянного тока. Изменение возбудимости ткани (например, нерва) под действием постоянного тока в области анода или катода называется физиологическим электротоном. В настоящее время установлено, что под действием отрицательного электрода - катода потенциал мембраны клеток снижается. Это явление называется физическим катэлектротоном. Под положительным - анодом,» он возрастает. Возникает физический анэлектртон. Так как, под катодом мембранный потенциал приближается к критическому уровню деполяризация, возбудимость клеток и тканей повышается. Под анодом мембранный потенциал возрастает и удаляется от критического уровня деполяризации, поэтому возбудимость клетки, ткани падает. Следует отметить, что при очень кратковременном действии постоянного тока (1 мсек и менее)МП не успевает измениться, поэтому не изменяется и возбудимость ткани под электродами.
Постоянный ток широко используется в клинике для лечения и диагностики. Например, с помощью него производится электростимуляция нервов и мышц, физиопроцедуры: ионофорез и гальванизация.
Классификация, физиологические свойства и функции нейронов. Механизм возбуждения нейронов.
Нейро́н, или невро́н (от др.-греч. νεῦρον — волокно, нерв) — структурно-функциональная единица нервной системы. Нейрон — электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов
Классификация:
1.В зависимости от нейромедиатора, используемого для передачи:
- холинергические – медиатор ацетилхолин (АХ);
- адренергические – норадреналин (НА);
- серотонинергические – серотонин (СТ);
- глицинергические – аминокислота глицин (ГЛИ);
- ГАМК-ергические – гамма-аминомасляная кислота (ГАМК);
- дофаминергические – дофамин (ДА);
- пептидергические – медиаторами являются нейропептиды (вещество Р, опиоидный пептид в-эндорфин и др.)
2. По форме (до 80 вариантов нейронов):
- звёздчатые
- пирамидальные,
- грушевидные
- веретеновидные
- паукообразные
- овальные
3. Функциональная (в зависимости от выполняемой функции и места в рефлекторной дуге):
- рецепторные (чувствительные, афферентные нейроны с помощью дендритов воспринимают воздействия внешней или внутренней среды, генерируют нервный импульс и передают его другим типам нейронов, встречаются только в спинальных ганглиях и чувствительных ядрах черепномозговых нервов)
- эффекторные (эфферентные нейроны передают возбуждение на рабочие органы, например, мышцы или железы, располагаются в передних рогах спинного мозга и вегетативных нервных ганглиях)
- вставочные (ассоциативные нейроны располагаются между рецепторными и эффекторными нейронами; по количеству их больше всего, особенно в ЦНС)
- секреторные (нейросекреторные клетки, специализированные нейроны, по своей функции напоминающие эндокринные клетки, синтезируют и выделяют в кровь нейрогормоны, расположены в гипоталамической области головного мозга.
4. По физиологической роли:
- возбуждающие
- тормозные
5. По количеству и характеру отростков:
- униполярные (имеющие один отросток)
- псевдоуниполярные (от тела отходит один отросток, который затем делится на 2 ветви)
- биполярные (2 отростка, один дендритоподобный, другой аксон)
- мультиполярные (имеют 1 аксон и много дендритов)
Физиологические функции:
1) возбудимость – способность приходить в состояние возбуждения в ответ на раздражение;
2) проводимость – способность передавать нервные возбуждение в виде потенциала действия от места раздражения по всей длине;
3) рефрактерность (устойчивость) – свойство временно резко снижать возбудимость в процессе возбуждения.
Нервная ткань имеет самый короткий рефрактерный период. Значение рефрактерности – предохранять ткань от перевозбуждения, осуществляет ответную реакцию на биологически значимый раздражитель;
4) лабильность – способность реагировать на раздражение с определенной скоростью. Лабильность характеризуется максимальным числом импульсов возбуждения за определенный период времени (1 с) в точном соответствии с ритмом наносимых раздражений.
Механизм возбуждения нейронов
Место возникновения ПД нейрона.
Формирование ПД начинается на мембране аксонного холмика (генераторный пункт нейрона). Синапсы на нем отсутствуют, возбудимость мембраны аксонного холмика в 3—4 раза превосходит возбудимость мембраны тела нейрона, что объясняется более высокой (примерно в 7 раз) концентрацией Na- каналов на аксонном холмике (на нем отсутствует миелиновая оболочка). ВПСП, возникающие в любых участках сомы нейрона суммируются, их суммарное электрическое поле достигает аксонного холмика и вызывает его деполяризацию, в результате чего начинает развиваться ПД на аксонном холмике. Это связано с тем, что постоянная длины мембраны тела нейрона составляет 1-2 мм, а диаметр тела нейрона — всего лишь 10-80 мкм.
На восходящей части ПД нейрона регистрируется перегиб на уровне 30— 40 мВ (задержка в развитии ПД), вследствие того, что возбудимость мембраны тела нейрона в 3—4 раза ниже возбудимости мембраны аксонного холмика, что также является подтверждением описанного механизма формирования ПД нейрона (рис. 33).
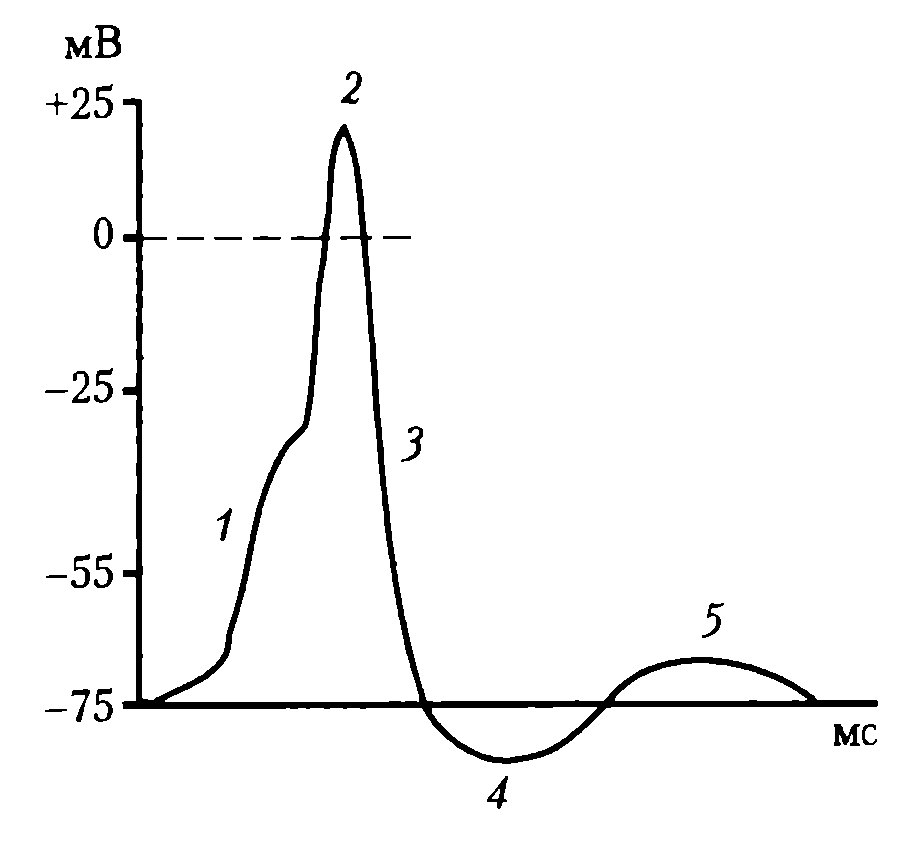 Рис.
33. Типичный ПД нейронов ЦНС: 1 –
деполяризация; 2 – инверсия; 3 –
реполяризация; 4 – следовая гиперполяризация;
5 – следовая деполяризация
Рис.
33. Типичный ПД нейронов ЦНС: 1 –
деполяризация; 2 – инверсия; 3 –
реполяризация; 4 – следовая гиперполяризация;
5 – следовая деполяризация
Проведение возбуждения в немиелинизированных и миелинизированных нервных волокнах. Функциональная классификация нервных волокон.
Миелиновые волокна:
Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
|
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А – Г); соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000) 1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно |
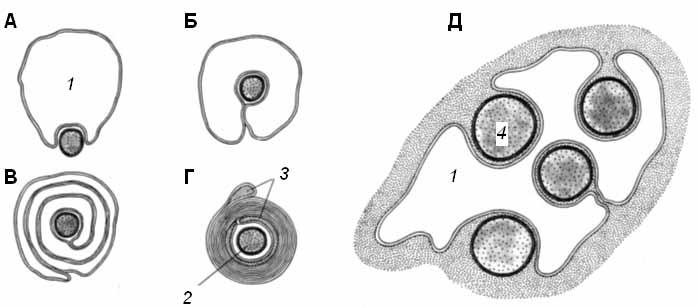
Безмиелиновые волокна:
Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
Механизм проведения возбуждения по безмиелиновым нервным волокнам:
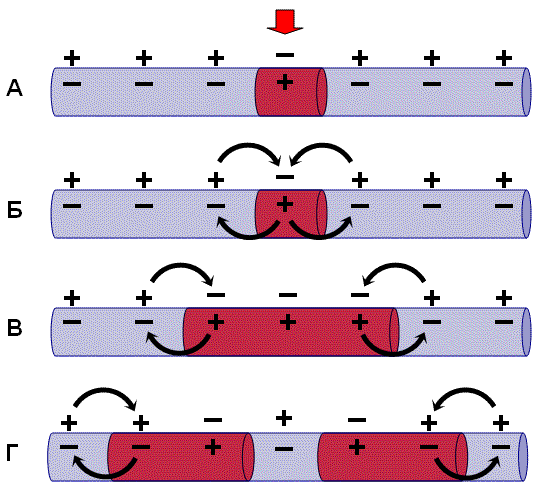
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
|
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте |
Механизм проведения возбуждения по миелиновым нервным волокнам:
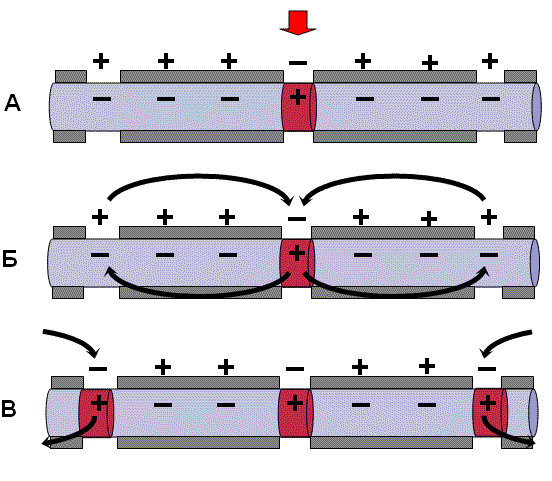
В миелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только в участках мембраны, расположенных в области перехватов Ранвье.
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
|
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте |
Функциональная классификация нервных волокон
Нервные волокна (neurofibrae) – это отростки нервных клеток, окруженные оболочками из нейроглиальных клеток.
Существуют различные классификации нервных волокон.
По наличию (или отсутствию) миелиновой оболочки нервные волокна делят на миелиновые и безмиелиновые.
Классификация нервных волокон по Эрлангеру – Гассеру основана на исследовании составного потенциала действия смешанного периферического нерва (например, седалищного). После электрической стимуляции такого нерва регистрируется целый спектр потенциалов действия, отражающий наличие в нерве волокон с разными скоростями проведения импульса. Согласно этой классификации выделяют три основных типа волокон – А, В, С, соответствующие трем волнам составного потенциала действия. Волокна типа А подразделяются на 4 подтипа – α, β, γ, δ. Данная классификация учитывает как чувствительные, так и двигательные волокна.
Классификация нервных волокон по Ллойду – Ханту учитывает диаметр волокна и скорость проведения импульса, но применяется только для чувствительных волокон. Согласно этой классификации выделяют четыре класса волокон – I,II,III,IV. ВолокнаIкласса подразделяют на 2 группы – а иb. Эти классификации не противоречат друг другу и представлены в сводной таблице.
Тип волокна |
Тип волокна (только чувствительные) |
Диаметр волокна (мкм) |
Скорость (м/с) |
Функция, тип обслуживаемого рецептора |
Чувствительные нервные волокна |
||||
Аα |
Ia, Ib |
13 – 20 |
80 – 120 |
Первичные афферентные волокна мышечных веретен и от сухожильных органов Гольджи. |
Аβ |
II |
6 – 12 |
35 – 75 |
Вторичные афферентнфе волокна мышечных веретен, кожные механорецепторы |
Аδ |
III |
1 – 5 |
5 – 30 |
Кожные механорецепторы, терморецепторы, ноцицепторы |
С |
IV |
0,2 – 1,5 |
0,5 – 2 |
Кожные механорецепторы, терморецепторы, ноцицепторы (немиелинизированные волокна) |
Двигательные нервные волокна |
||||
Аα |
− |
12 – 20 |
72 – 120 |
Экстрафузальные волокна скелетных мышц |
Аγ |
− |
2 – 8 |
12 – 48 |
Интрафузальные мышечные волокна |
В |
− |
1 – 3 |
6 – 18 |
Преганглионарные вегетативные волокна |
С |
− |
0,2 – 2 |
0,5 – 2 |
Постганглионарные вегетативгые (немиелинизированные волокна) |
Закон анатомической и функциональной целостности нервного волокна. Парабиоз по Н.Е. Введенскому, фазы парабиоза. Практическое применение парабиоза в медицине.
Закон анатомической и физиологической целостности нервного волокна.Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву. Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е., к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается.
Парабиоз
Явление парабиоза открыто русским физиологом Н.Е.Введенским в 1901 г. при изучении возбудимости нервно-мышечного препарата. Состояние парабиоза могут вызвать различные воздействия – сверхчастые, сверхсильные стимулы, яды, лекарства и другие воздействия как в норме, так и при патологии.
Н. Е. Введенский обнаружил, что если участок нерва подвергнуть воздействию повреждающего агента, то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется. Такое состояние пониженной лабильности и было названо Н. Е. Введенским парабиозом. Состояние парабиоза возбудимой ткани возникает под влиянием сильных раздражителей и характеризуется фазными нарушениями проводимости и возбудимости.
Выделяют 3 фазы: первичную, фазу наибольшей активности (оптимум) и фазу сниженной активности (пессимум). Третья фаза объединяет 3 последовательно сменяющие друг друга стадии: уравнительную (провизорная, трансформирующая – по Н.Е.Введенскому), парадоксальную и тормозную.
Первая фаза (примум) характеризуется снижением возбудимости и повышением лабильности. Во вторую фазу (оптимум) возбудимость достигает максимума, лабильность начинает снижаться. В третью фазу (пессимум) возбудимость и лабильность снижаются параллельно и развивается 3 стадии парабиоза.
Первая стадия - уравнительная по И.П.Павлову - характеризуется выравниванием ответов на сильные, частые и умеренные раздражения. В уравнительную фазу происходит уравнивание величины ответной реакции на частые и редкие раздражители. В нормальных условиях функционирования нервного волокна величина ответной реакции иннервируемых им мышечных волокон подчиняется закону силы: на редкие раздражители ответная реакция меньше, а на частые раздражители—больше. При действии парабиотического агента и при редком ритме раздражении (например, 25 Гц) все импульсы возбуждения проводятся через парабиотический участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражении (100Гц) последующие импульсы могут поступать в тот момент, когда нервное волокно еще находится в состоянии относительной рефрактерности, вызванной предыдущим потенциалом действия. Поэтому часть импульсов не проводится. Если проводится только каждое четвертое возбуждение (т.е. 25 импульсов из 100) ,то амплитуда ответной реакции становится такой же, как на редкие раздражители (25Гц)—происходит уравнивание ответной реакции.
Вторая стадия характеризуется извращенным реагированием – сильные раздражения вызывают меньший ответ, чем умеренные. В эту - парадоксальную фазу происходит дальнейшее снижение лабильности. При этом на редкие и частые раздражители ответная реакция возникает, но на частые раздражители она значительно меньше, т. к. частые раздражители еще больше снижают лабильность, удлиняя фазу абсолютной рефрактерности. Следовательно, наблюдается парадокс— на редкие раздражители ответная реакция больше, чем на частые.
В тормозную фазу лабильность снижается до такой степени, что и редкие, и частые раздражители не вызывают ответной реакции. При этом мембрана нервного волокна деполяризована и не переходит в стадию реполяризации, т. е. не восстанавливается ее исходное состояние. Ни сильные, ни умеренные раздражения не вызывают видимой реакции, в ткани развивается торможение.
Парабиоз— явление обратимое. Если парабиотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, но в обратной последовательности. Однако, при действии сильных раздражителей за тормозной стадией может наступить полная потеря возбудимости и проводимости, а в дальнейшем – гибель ткани.
Работы Н.Е.Введенского по парабиозу сыграли важную роль в развитии нейрофизиологии и клинической медицины, показав единство процессов возбуждения, торможения и покоя, изменили господствовавший в физиологии закон силовых отношений, согласно которому реакция тем больше, чем сильнее действующий раздражитель.
Явление парабиоза лежит в основе медикаментозного локального обезболивания. Влияние анестезирующих веществ связано с понижением лабильности и нарушением механизма проведения возбуждения по нервным волокнам.
Физиологические свойства и функции поперечно-полосатых мышечных клеток. Механизм сокращения поперечно-полосатых мышечных клеток.
Физиологические свойства поперечнополосатых мышц:
- возбудимость (ниже, чем в нервном волокне, что объясняется низкой величиной мембранного потенциала)
- низкая проводимость, порядка 10–13 м/с
- рефрактерность (занимает по времени больший отрезок, чем у нервного волокна)
- лабильность
- сократимость (способность укорачиваться или развивать напряжение)
Физиологические функции поперечнополосатых мышц:
- двигательная (динамическая и статическая)
- обеспечения дыхания
- мимическая
- рецепторная
- депонирующая
- терморегуляторная
Механизм сокращения поперечно- полосатой мускулатуры
При световой микроскопии было замечено, что в момент сокращения ширина А-диска не уменьшается, а Iдиски и Н-зоны саркомеров суживаются. При электронной, микроскопии было установлено, что длина нитей актина и миозина в момент соскращения не изменяется. Поэтому Хаксли и Хэнсон разработали теорию скольжения нитей. Согласно этой теории, мышца укорачивается в результате движения тонких актиновых нитей в промежутки между миозиновыми. Это приводит к укорочению каждого саркомера, образующего миофибриллы. Скольжение же нитей обусловлено тем, что при переходе в активное состояние головки отростков миозина связываются с центрами актиновых нитей и вызывают их движение относительно себя (гребковые движения). Но это последний этап всего сократительного механизма.
Сокращение начинается с того, что в области концевой пластинки двигательного нерва возникает ПД. Он с большой скоростью распространяется по сарколемме и переходит с неѐ на продольные трубочки и цистерны. Возникает деполяризация мембраны цистерн и из них в саркоплазму высвобождаются ионы кальция. На нитях актина расположены молекулы еще двух белков – тропонина и тропомиозина. При низкой (менее 10-8 моль) концентрации кальция, т.е. в состоянии покоя, тропомиозин блокирует присоединение мостиков миозина к нитям актина. Когда ионы кальция начинают выходить из СР, молекула тропонина изменяет свою форму таким образом, что освобождает активные центры актина от тропомиозина. К этим центрам присоединяются головки миозина и начинается скольжение за счет ритмического прикрепления и разъединения поперечных мостиков с нитями актина. При этом головки ритмически продвигаются по нитям актина к Z-мембранам. Для полного сокращения мышцы необходимо 50 таких циклов.
Передача сигнала от возбужденной мембраны к миофибриллам называется электромеханическим сопряжением. Когда генерация ПД прекращается и мембранный потенциал возвращается к исходному уровню, начинает работать Са-насос (фермент Са-АТФаза). Ионы кальция вновь закачиваются в цистерны саркоплазматического ретикулума и их концентрация падает ниже 10-8 моль. Молекулы тропонина приобретают исходную форму и тропомиозин вновь начинает блокировать активные центры актина. Головки миозина отсоединяются от них, и мышца за счет эластичности приходит в исходное расслабленное состояние.
Одиночное сокращение скелетных мышц, его фазы. Тетаническое сокращение скелетных мышц. Зубчатый и гладкий тетанус мышц.
При нанесении на двигательный нерв или мышцу одиночного порогового или сверхпорогового раздражения, возникает одиночное сокращение. При его графической регистрации, на полученной кривой можно выделить три последовательных периода:
1) Латентный период: это время от момента нанесения раздражения до начала сокращения. Его длительность около 1-2 мс. Во время латентного периода генерируется и распространяется ПД, происходит высвобождение кальция из СР, взаимодействие актина с миозином и т.д.
2) Период укорочения: в зависимости от типа мышцы (быстрая или медленная) его продолжительность от 10 до 100 мсек.,
3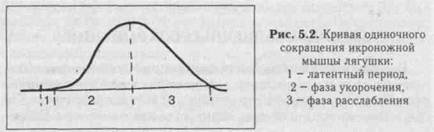 )
Период
расслаблени:
его длительность несколько больше, чем
укорочения (0,05-0.06 сек.)
)
Период
расслаблени:
его длительность несколько больше, чем
укорочения (0,05-0.06 сек.)
В режиме одиночного сокращения мышца способна работать длительное время без утомления, но его сила незначительна. Поэтому в организме такие сокращения встречаются редко, например, так могут сокращаться быстрые глазодвигательные мышцы, мышцы сгибателей пальцев. Чаще одиночные сокращения суммируются.
Суммация – это сложение двух последовательных сокращений при нанесении на нее двух пороговых или сверхпороговых раздражений, интервал между которыми меньше длительности одиночного сокращения, но больше продолжительности рефракторного периода.
Виды:
- неполная суммация: возникает в том случае, если повторное раздражение наносится на мышцу, когда она уже начала расслабляться
- полная возникает тогда, когда повторное раздражение действует на мышцу до начала периода расслабления, т.е. в конце периода укорочения.
Амплитуда сокращения при полной суммации выше, чем неполной. Если интервал между двумя раздражениями еще больше уменьшить, например, нанести второе в середине периода укорочения, то суммации не будет, потому что мышца находится в состоянии рефрактерности.
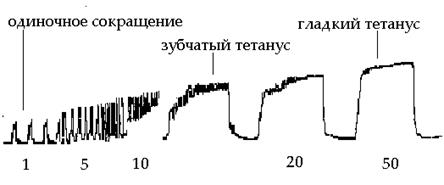
Тетаническое сокращение
Тетанус – это длительное сокращение мышцы, возникающее в результате суммации нескольких одиночных сокращений, развивающихся при нанесении на нее ряда последовательных раздражений.
Формы
- зубчатый тетанус: наблюдается в том случае, если каждое последующее раздражение действует на мышцу, когда она уже начала расслабляться. Т.е. наблюдается неполная суммация.
- гладкий тетанус: возникает тогда, когда каждое последующее раздражение наносится в конце периода укорочения. Т.е. имеет место полная суммация отдельных сокращений. Амплитуда гладкого тетануса больше, чем зубчатого. В норме мышцы человека сокращаются в режиме гладкого тетануса. Зубчатый возникает при патологии, например, тремор рук при алкогольной интоксикации и болезни Паркинсона.
Физиологические свойства и функции гладкомышечных клеток. Виды хеморецепторов мембраны гладкомышечных клеток.
Функции:
1.поддержание давления в полых органах
2.регуляция давления в кровеносных сосудах
3.опорожнение полых органов и продвижение их содержимого
Физиологические особенности гладких мышц
Гладкие мышцы имеют те же физиологические свойства,что и скелетные мышцы ,но имеют и свои особенности :
1.нестабильный мембранный потенциал , который поддерживает мышцы в состоянии постоянного частичного сокращения – тонуса
2.самопроизвольную автоматическую активность
3.сокращение в ответ на растяжение
4.пластичность
5.высокую чувствительность к химическим веществам