

2. Строение и состав мембран: структура и свойства липидов,белков, углеводов мембран. Общие свойства мембран и их функции.
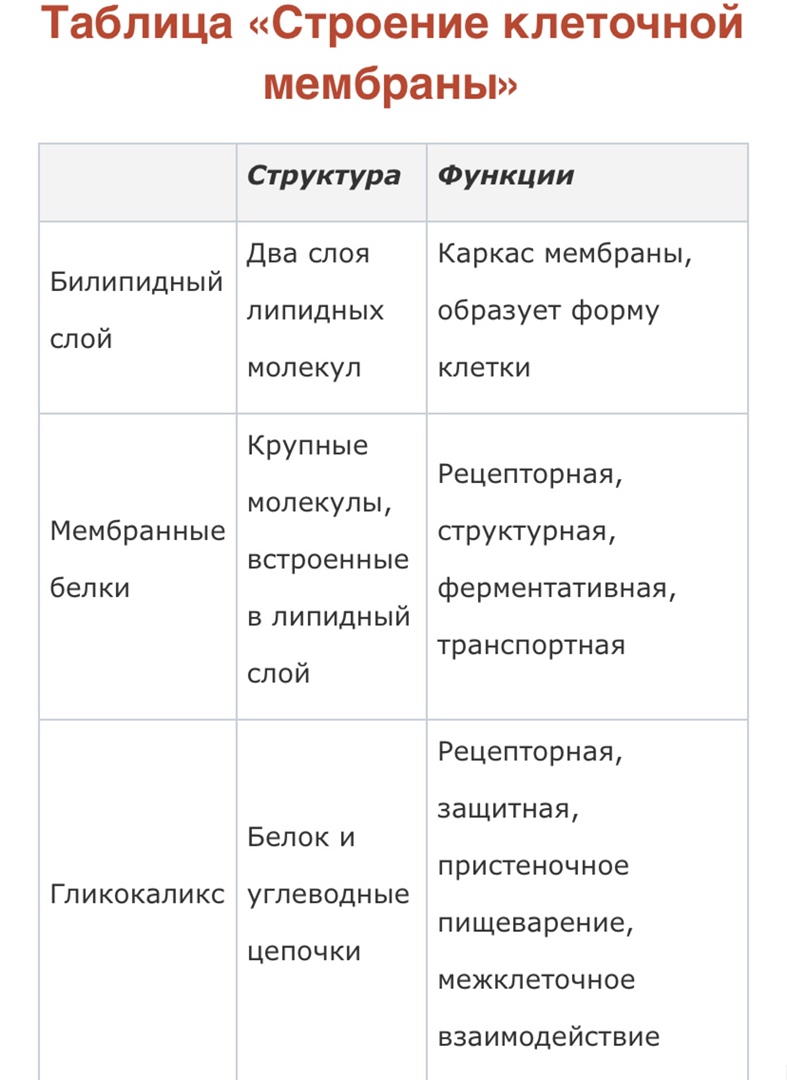
Мембрана – это сверхтонкая структура, образующая поверхности органоидов и клетки в целом. Все мембраны имеют сходное строение и связаны в одну систему. Мембраны клетки химически однородны и состоят из белков и липидов различных групп: фосфолипидов; галактолипидов; сульфолипидов. Также в их состав входят нуклеиновые кислоты, полисахариды и другие вещества. Физические свойства При нормальной температуре мембраны находятся в жидкокристаллическом состоянии и постоянно колеблется. Их вязкость близка к вязкости растительного масла. Мембрана способна к восстановлению, прочна, эластична и имеет поры. Толщина мембран 7 – 14 нм. Модель Обычно строение мембран описывается при помощи жидкостно-мозаичной модели. Мембрана имеет каркас – два ряда липидных молекул, плотно, как кирпичики прилегающих друг к другу. С обеих сторон поверхность липидов покрыта белками. Мозаичная картина образуется неравномерно распределёнными на поверхности мембраны молекулами белков. Все мембраны клетки по локализации можно разделить на следующие типы: наружная; ядерная; мембраны органоидов. Наружная цитоплазматическая мембрана, или плазмолемма, является границей клетки. Соединяясь с элементами цитоскелета, она поддерживает её форму и размеры. Ядерная мембрана, или кариолемма, является границей ядерного содержимого. Она построена из двух мембран, очень похожих на наружную. Внешняя мембрана ядра связана с мембранами эндоплазматической сети (ЭПС) и, через поры, с внутренней мембраной. Гликокаликс Животные клетки имеют на поверхности наружной мембраны углеводный слой, называемый гликокаликсом. Наиболее развит гликокаликс в клетках кишечного эпителия, где он создаёт условия для пищеварения и защищает плазмолемму. Строительным "кирпичиком" мембраны являются фосфолипиды. Вследствие своего строения фосфолипиды амфифильны, поскольку обладают полярной "головой", образованной заряженной группой фосфата (иногда ионизированной группировкой, присоединенной к ней, пример- фосфатидилхолин). Полярной частью молекула хорошо взаимодействует с водной средой. Кроме того, у фосфолипидов имеется довольно длинный неполярный (гидрофобный) "хвост". Хвост образован остатками карбоновых кислот, очень часто- ненасыщенных, содержащих кратную связь примерно в середине цепи. Кратная связь чаще всего находится в цис-конфигурации, поэтому "ножка" не прямая, а как бы изогнутая . В мембранах содержатся липиды трех классов: фосфолипиды, холестерин и гликолипиды. Наиболее важная группа, фосфолипиды, включает фосфатидилхолин (лецитин). Холестерин присутствует во внутриклеточных мембранах животных клеток (за исключением внутренней мембраны митохондрий). Гликолипиды входят в состав многих мембран (например, во внешний слой плазматических мембран). В состав гликолипидов входят углеводные функциональные группы, которые ориентируются в водную фазу. Липиды мембран представляют собой амфифильные молекулы с полярной гидрофильной головкой (голубого цвета) и неполярным липофильным хвостом (желтого цвета). В водной среде они агрегируют за счет гидрофобных взаимодействий.
Мембранные белки Протеины могут связываться с мембраной различным путем. Интегральные мембранные белки имеют трансмембранные спирализованные участки (домены), которые однократно или многократно пересекают липидный бислой. Такие белки прочно связаны с липидным окружением. Периферические мембранные белки удерживаются на мембране с помощью липидного «якоря»и связаны с другими компонентами мембраны; например, они часто бывают ассоциированы с интегральными мембранными белками. У интегральных мембранных белков фрагмент пептидной цепи, пересекающий липидный бислой, обычно состоит из 21-25 преимущественно гидрофобных аминокислот, которые образуют правую α-спираль с 6 или 7 витками (трансмембранная спираль).