

Третий этап:
Все реакции этого этапа идут в митохондриях.
Ацетил-SКоА (и кетокислоты) включается в реакции цикла трикарбоновых кислот, где углероды веществ окисляются до углекислого газа.
Выделенные атомы водорода соединяются с НАД и ФАД и восстанавливают их. После этого НАДН и ФАДН2 переносят водород в цепь ферментов дыхательной цепи, расположенную на внутренней мембране митохондрий. Здесь в результате процесса под названием "окислительное фосфорилирование" образуется вода и главный продукт биологического окисления – АТФ.
Часть выделенной на этом этапе энергии молекулы рассеивается в виде тепла и около 46% энергии исходного вещества усваивается, т.е. запасается в связях АТФ и ГТФ.
Пировиноградная кислота(ПВК, пируват) является продуктом окисления глюкозы и некоторых аминокислот. Ее судьба различна в зависимости от доступности кислорода в клетке:
в анаэробных условиях она восстанавливается до молочной кислоты.
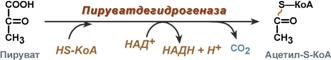
в аэробных условиях пируват симпортом с ионами Н+, движущимися по протонному градиенту, проникает в митохондрии. Здесь происходит его превращение до уксусной кислоты, переносчиком которой служит коэнзим А.
Пируватдегидрогеназный мульферментный комплекс
Суммарное уравнение отражает окислительное декарбоксилирование пирувата, восстановление НАД до НАДН и образование ацетил-SKoA.
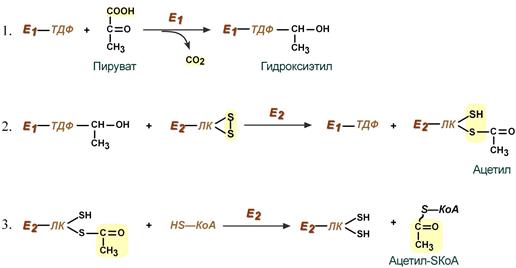
Превращение состоит из пяти последовательных реакций, осуществляется мультиферментным комплексом, прикрепленным к внутренней митохондриальной мембране со стороны матрикса.
В составе комплекса насчитывают 3 фермента и 5 коферментов:
Пируватдегидрогеназа (Е1, ПВК-дегидрогеназа), ее коферментом является тиаминдифосфат (ТДФ), катализирует 1-ю реакцию.
Дигидролипоат-ацетилтрансфераза (Е2), ее коферментом является липоевая кислота, катализирует 2-ю и 3-ю реакции.
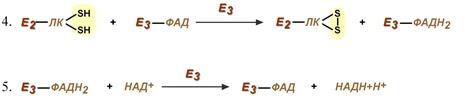
Дигидролипоат-дегидрогеназа (Е3), кофермент – ФАД, катализирует 4-ю и 5-ю реакции.
Помимо указанных коферментов, которые прочно связаны с соответствующими ферментами, в работе комплекса принимают участие коэнзим А и НАД.
Суть первых трех реакций сводится к:
декарбоксилированию пирувата (катализируется пируватдегидрогеназой, Е1),
окислению пирувата до ацетила,
переносу ацетила на коэнзим А (катализируется дигидролипоамид-ацетилтрансферазой, Е2).
Оставшиеся 2 реакции необходимы для возвращения липоевой кислоты и ФАД в окисленное состояние (катализируются дигидролипоат-дегидрогеназой, Е3).
При этом образуется НАДН.
Регуляция пируватдегидрогеназного комплекса:
Регулируемым ферментом ПВК-дегидрогеназного комплекса является первый фермент – пируватдегидрогеназа (Е1). Два вспомогательных фермента – киназа и фосфатаза обеспечивают регуляцию активности пируватдегидрогеназы путем ее фосфорилирования и дефосфорилирования.
Вспомогательный фермент киназа активируется при избытке конечного продукта биологического окисления АТФ и продуктов ПВК-дегидрогеназного комплекса – НАДН и ацетил-S-КоА. Активная киназа фосфорилирует пируватдегидрогеназу, инактивируя ее, в результате первая реакция процесса останавливается.
Фермент фосфатаза, активируясь ионами кальция или инсулином, отщепляет фосфат и активирует пируватдегидрогеназу.
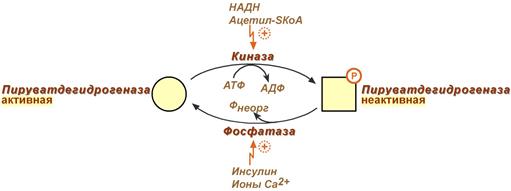
Таким образом,работа пируватдегидрогеназы подавляется при избытке в митохондрии (в клетке) АТФ и НАДН, что позволяет снизить окисление пирувата и, следовательно, глюкозы в случае когда энергии достаточно.
Если АТФ мало или имеется влияние инсулина, то образуется ацетил-SКоА. Последний в зависимости от условий будет направляться либо в цикл трикарбоновых кислот с образованием энергии АТФ, либо на синтез холестерина и жирных кислот.
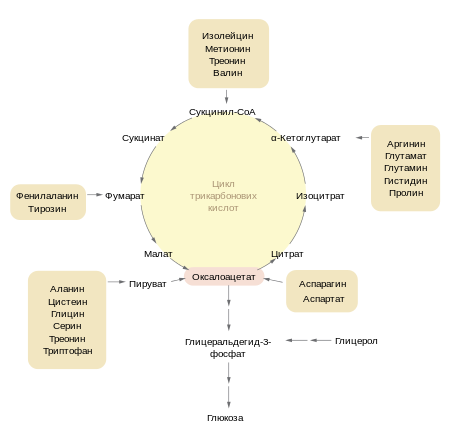
Образующийся в ПВК-дегидрогеназной реакции ацетил-SКоА далее вступает в цикл трикарбоновых кислот (ЦТК, цикл лимонной кислоты, цикл Кребса). Кроме пирувата, в цикл вовлекаются кетокислоты, поступающие из катаболизма аминокислот или каких-либо иных веществ.
Цикл протекает в матриксе митохондрий и представляет собой окисление молекулы ацетил-SКоА в восьми последовательных реакциях:
Связываются ацетил и оксалоацетат (щавелевоуксусная кислота) с образованием цитрата (лимонной кислоты),
Изомеризация лимонной кислоты до изоцитрата
Реакция дегидрирования с сопутствующим выделением СО2 и восстановлением НАД
Реакция дегидрирования с сопутствующим выделением СО2 и восстановлением НАД
Образуется ГТФ, это реакция субстратного фосфорилирования.
Последовательно происходит ФАД-зависимое дегидрирование сукцината(янтарной кислоты),
Гидратация фумаровой кислоты до малата (яблочная кислота),
НАД-зависимое дегидрирование с образованием в итоге оксалоацетата.
В итоге после восьми реакций цикла вновь образуется оксалоацетат.
Функции ЦТК
1. Энергетическая:
генерация атомов водорода для работы дыхательной цепи, а именно трех молекул НАДН и одной молекулы ФАДН2,
синтез одной молекулы ГТФ (эквивалентна АТФ).
Анаболическая:
В ЦТК образуются:
предшественник гема – сукцинил-SКоА,
кетокислоты, способные превращаться в аминокислоты – α-кетоглутарат для глутаминовой кислоты, оксалоацетат для аспарагиновой,
лимонная кислота, используемая для синтеза жирных кислот,
оксалоацетат, используемый для синтеза глюкозы.
2. Стадии свободно-радикального окисления липидов. Биосинтез ВЖК происходит во время приема пищи и в цитоплазме. Субстратом является ацетил-S-КоА, который образуется из
ПВК в процессе ресщепления глюкозы
Глюкагенных аминокислот-аланин, цистеин, серин, глицин, треонин, которые превращаются сначала в ПВК, а лизин- сразу в ацетил-S-КоА
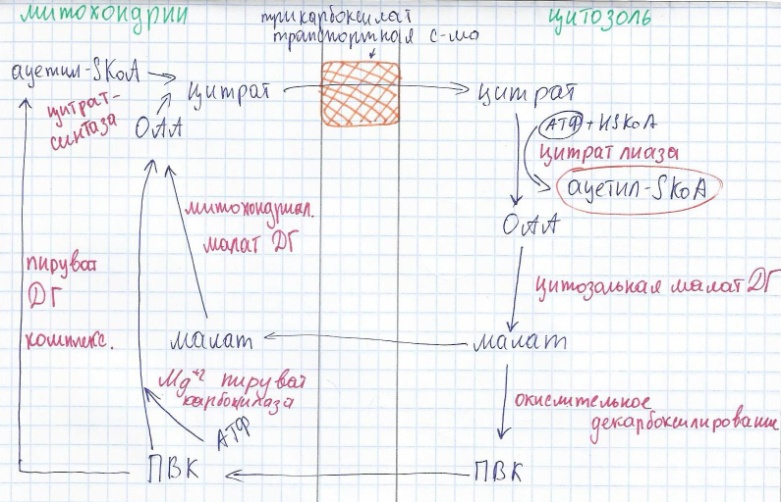
А) Транспорт ацетил-S-КоА из МХ в цитозоль
Происходит образование цитрата из ОАА и ацетил-СКоА. Далее в цитозоле происходит образование ОАА с выходом ацетил-СКоА
Б) Синтез малонин-КоА – в результате карбоксилирования ацетил-SKoA под действием ацетил-SKoА-карбоксилазы
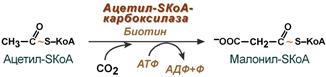
Положительными модуляторами ацетил-SKoА-карбоксилазыявляются:цитрат, АТФ
Отрицательными модуляторами являются:АДФ, АМФ, Ацетил-SКоА
В)Формирование цепи ВЖК
Катализирует синтез СИНТАЗА ВЖК:
Ферменты: ацетилтрансфераза, малонилтрансфераза, кетоацетилсинтетаза, кетоацилпедуктаза, гидратаза, редуктаза, тиоэстераза
Ацилпереносящий белок(АПБ)
Когда образуется С16:0 цикл останавливается, тк он не является субстратом для кондесирующего фермента
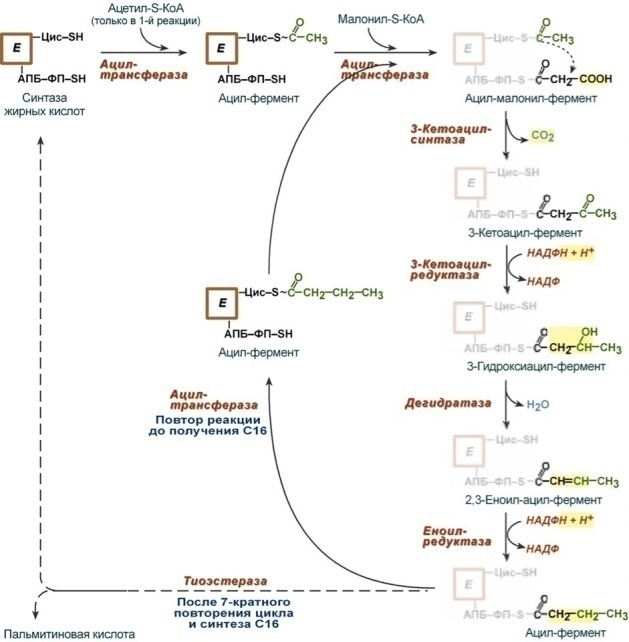
Далее из С16:0 образуются другие ВЖК (в основном С18:0) при помощи малонил-SKoA.
Включение двойных связей в радикалы ЖК называется десатурацией(происходит в ЭПС). В образовании двойных связей участвуют
Десатуразы ЖК-не могут образовывать двойные связи в радикалах жк дистальнее 9 атома, поэтому жк омега-3 и омега-6 не синтезируются в организме
Молекулярный кислород
НАДФН+Н+
Цитохром В5
ФАД-зависимая редуктаза цитохрома В5
Метаболическая регуляция
1)ассоциация – диссоциация. В неактивной форме фермент(ацетил-КоА карбоксилаза) представляет собой отдельные комплексы из которых состоит 4 субъединицы
Активаторы фермента:цитрат
Ингибитор фермента: пальмитоил – КоА
2)фосфорилирование-дефосфорилирование. Фермент активен в дефосфорилированном состоянии и наоборт.
Гормональная регуляция
В постабсортивном периоде или при физической нагрузке глюкагон или адреналин стимулируют фосфорилирование фермента и синтез прекращается
В абсорбтивном периоде инсулин способствует дефосфорилированию, и синтез усиливается
Перекисное(свободно-радикальное) окисление липидов-это многоступенчатый процессокисления ПНЖК, протекающий через образование короткоживущих свободных радикалов.
В результате ПОЛ образуются «срезанный хвостики» ПНЖК и из них получаются первичные продукты ПОЛ:оксикислоты, этилен, альдегиды. Из «срезанных хвостиков»,имеющих две и более двойные связи, образуется вторичный продукт ПОЛ-малоновый диальдегид. Он легко взаимодействует с белками, липидами, белково-липидными образованиями, образуя внутри- и межмолекулярные сшивки.
Первичные продукты ПОЛ гидрофильны и поэтому выталкиваются на поверхность мембраны вместе с ЖК, на которой находится. А далее фосфолипаза А2(находится на поверхности мембраны) «выстригает» радикалы ПНЖК из ФЛ. ФЛ, утратившие остатки ЖК, превращаются в лизофосфатиды ⇒в мембране повляются ОН-группы,образующие гидрофильные поры⇒нарушается избирательная проницаемость
При повышении интенсивности ПОЛ и появлении гидрофильных пор, Н+ может попадать в клетку,минуя АТФ-синтетазы⇒оксиление субстратов дых.цепи идет в холостую,тк АТФ не синтезируется.
Также через эти поры может проникать Na+ . Это приводит к нарушению симпорта АК, глюкозы.
В результате работы Na/Ca-обменника усиливается поступление Ca⇒ активация C-зависимых гидролаз,разрушающие собственные структуры клеток.