

Лекции по общей биохимии (О. Тимин)
.pdfСтроение и обмен липидов |
240 |
|
|
|
|
ФОР М Ы ТР АН СПОР Т А Л ИПИД ОВ В К Р ОВИ
Липиды транспортируются в водной фазе крови в составе особых частиц – липопротеинов. Поверхность частиц гидрофильна и сформирована белками, фосфолипидами и свободным холестеролом. Триацилглицеролы и эфиры холестерола составляют гидрофобное ядро.
Белки в липопротеинах обычно называются апобелками, выделяют несколько их типов – А, В, С, D, Е. В каждом классе липопротеинов находятся соответствующие ему апобелки,
выполняющие структурную, ферментативную и кофакторную функции.
Липопротеины различаются по соотношению триацилглицеролов, холестерола и его эфиров, фосфолипидов и как сложные белки состоят из четырех классов.
o хиломикроны (ХМ),
o липопротеины очень низкой плотности
(ЛПОНП, пре-β-липопротеины, пре-β-ЛП), o липопротеины низкой плотности
(ЛПНП, β-липопротеины, β-ЛП),
o липопротеины высокой плотности
(ЛПВП, αлипопротеины, αЛП).
Хиломикроны и ЛПОНП ответственны, в первую очередь, за транспорт жирных кислот в составе ТАГ. Липопротеины высокой и низкой плотности – за транспорт холестерола и жирных кислот в составе эфиров ХС.
ТР АН СП ОР Т ТР И АЦ ИЛГЛ И Ц Е Р ОЛОВ
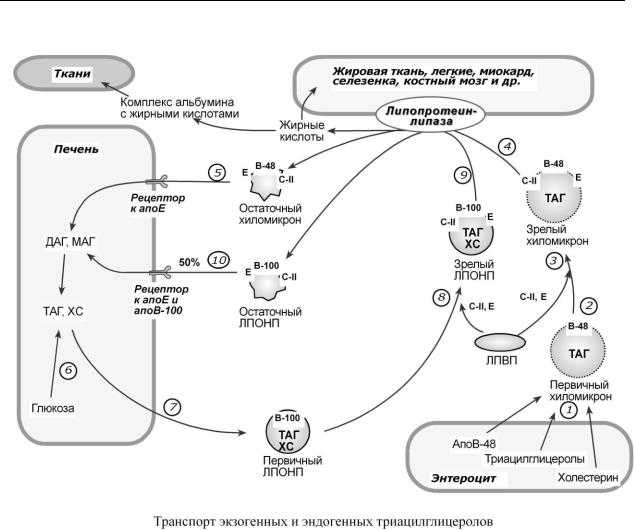
Транспорт ТАГ от кишечника к тканям (экзогенные ТАГ) осуществляется в виде хиломикронов, от печени к тканям (эндогенные ТАГ) – в виде липопротеинов очень низкой плотности.
В транспорте ТАГ к тканям в целом можно выделить последовательность следующих событий:
1.Образование незрелых первичных ХМ в кишечнике.
2.Движение первичных ХМ через лимфатические протоки в кровь.
3.Созревание ХМ в плазме крови – получение белков апоС-II и апоЕ от ЛПВП.
4.Взаимодействие с липопротеинлипазой эндотелия капилляров и потеря большей части ТАГ. Образование остаточных ХМ.
5.Переход остаточных ХМ в гепатоциты и полный распад их структуры.
6.Синтез ТАГ в печени из пищевой глюкозы и использование ТАГ, пришедших в составе остаточных ХМ.
7.Образование первичных ЛПОНП в печени.
8.Созревание ЛПОНП в плазме крови – получение белков апоС-II и апоЕ от ЛПВП.
9.Взаимодействие с липопротеинлипазой эндотелия капилляров и потеря большей части ТАГ. Образование остаточных ЛПОНП (по-другому липопротеины промежуточной плотности, ЛППП).
10.Остаточные ЛПОНП переходят в гепатоциты и полностью распадаются, либо остаются в плазме крови. После воздействия на них печеночной ТАГ-липазы в синусоидах печени ЛПОНП превращаются в ЛПНП.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
241 |
ХАРАКТЕРИСТИКА ХИЛОМИКРОНОВ
Общая характеристика
1.Формируются в кишечнике из ресинтезированных жиров.
2.В их составе преобладают ТАГ (до 90%), мало белка, фосфолипидов и холестерола.
3.Основным апобелком является апоВ-48, это структурный липопротеин. В плазме крови ХМ получают от ЛПВП белки апоС-II и апоЕ.
4.В норме натощак не обнаруживаются, в крови появляются после приема пищи, поступая из лимфы через грудной лимфатический проток, и исчезают через 10-12 часов.
5.Не атерогенны.
Функция
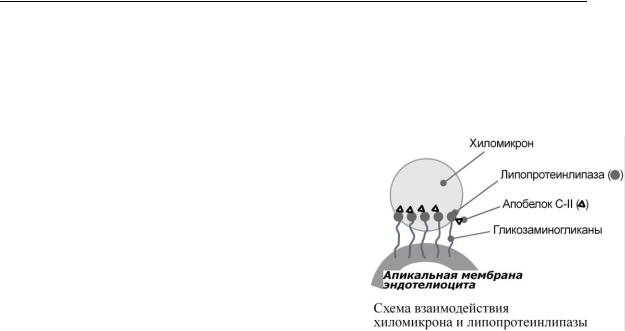
Транспорт экзогенных ТАГ из кишечника в ткани, запасающие или использующие жиры, в основном, в жировую ткань, миокард, скелетные мышцы, лактирующую молочную железу, в меньшей степени в костный мозг, легкие, почки, селезенку. На эндотелии капилляров этих тканей имеется фермент липопротеинлипаза.
Строение и обмен липидов |
242 |
Метаболизм
1.После ресинтеза жиров в эпителиоцитах кишечника формируются первичные хиломикроны, имеющие только апоВ-48.
2.Из-за большого размера они не проникают напрямую в кровеносное русло и эвакуируются через лимфатическую систему, попадая в кровь через грудной лимфатический проток.
3.В крови хиломикроны взаимодействуют с
ЛПВП и приобретают от них апоС-II и апоЕ, образуя зрелые формы. Белок апоС-II является кофактором фермента липопротеинлипазы, белок апоЕ необходим для удаления из крови остаточных хиломикронов в печень.
4.На эндотелии капилляров вышеперечисленных тканей находится фермент липопротеинлипаза (ЛПЛ). Количество фермента в жировой ткани увеличивается при действии
инсулина.
5.После взаимодействия хиломикрона с ферментом триацилглицеролы, находящиеся в
составе хиломикронов, гидролизуются с образованием свободных жирных кислот и моноацилглицеролов Жирные кислоты перемещаются в клетки органа, либо остаются в плазме крови и в комплексе с альбумином разносятся с кровью в другие ткани. Липопротеинлипаза способна расщепить до 90% всех ТАГ, находящихся в хиломикроне или ЛПОНП.
6.После окончания работы ЛПЛ остаточные хиломикроны попадают в гепатоциты посредством апоЕ-рецепторного эндоцитоза и разрушаются.
ХАРАКТЕРИСТИКА ЛИПОПРОТЕИНОВ ОЧЕНЬ НИЗКОЙ ПЛОТНОСТИ
Общая характеристика
1.Синтезируются в печени из эндогенных и экзогенных липидов,
2.В их составе преобладают ТАГ, около 40% от массы составляют белки, фосфолипиды и холестерол,
3.Основным белком является апоВ-100, выполняющий структурную функцию,
4.В норме концентрация 1,3-2,0 г/л,
5.Слабо атерогенны.
Функция
Транспорт эндогенных и экзогенных ТАГ от печени в ткани, запасающие и использующие жиры.
Метаболизм
1.Первичные ЛПОНП образуются в печени из пищевых жиров, достигающих гепатоцитов с остаточными хиломикронами, и новосинтезированных из глюкозы жиров, содержат только апоВ-100;
2.В крови первичные ЛПОНП взаимодействуют с ЛПВП и приобретают от них апоС-II и апоЕ, образуя зрелые формы.
3.Аналогично хиломикронам, на эндотелии капилляров ряда тканей зрелые ЛПОНП подвергаются воздействию липопротеинлипазы с образованием свободных жирных кислот. Жирные кислоты перемещаются в клетки органа, либо остаются в плазме крови и в комплексе с альбумином разносятся с кровью в другие ткани.
4.Остаточные ЛПОНП (также называемые липопротеины промежуточной плотности, ЛППП)
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
243 |
|
|
|
|
|
oлибо попадают в гепатоциты посредством эндоцитоза, связанного с рецепторами к апоЕ и апоВ-100-белкам,
oлибо после воздействия на них липазы печени (только в синусоидах печени) превращаются в следующий класс липопротеинов – липопротеины низкой плотности (ЛПНП).
ПР ОМ Е ЖУ ТО ЧНЫ Й ОБМ Е Н Л ИПИД О В
И С Т О Ч Н И К И И С У Д Ь Б А Ж И Р Н Ы Х К И С Л О Т
Жирные кислоты входят в состав большей части липидов. Поэтому вопросы метаболизма липидов – это часто вопросы обмена жирных кислот, их источники и пути дальнейших превращений.
Судьба жирной кислоты зависит от ее строения (насыщенная ЖК или полиненасыщенная ЖК) и от внутриклеточных условий (наличие или отсутствие энергии в клетке, гормональные влияния).
Абсорбтивный период
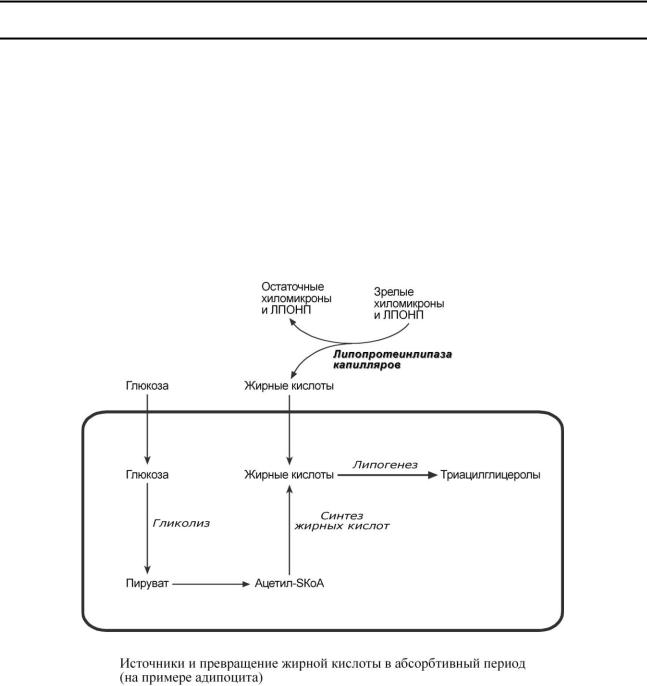
В течение нескольких часов после приема пищи насыщенные и мононенасыщенные жирные кислоты (НЖК) поступают от кишечника в определенные ткани из хиломикронов.
Параллельно в этот период времени в печени жирные кислоты способны синтезироваться из избытка экзогенной глюкозы и полученные эндогенные жирные кислоты этерифицируются с глицеролом в реакциях липогенеза с образованием ТАГ. Далее они транспортируются из печени в ткани, имеющие липопротеинлипазу, в составе ЛПОНП.
В клетках жировой ткани после приема пищи насыщенная жирная кислота либо синтезируется из глюкозы, либо поступает из хиломикронов и ЛПОНП. Далее она направляется в липогенез и запасается в составе ТАГ.
Строение и обмен липидов |
244 |
|
|
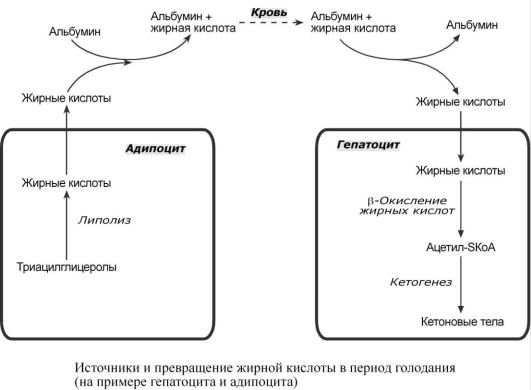
Если липопротеинлипазы нет в ткани, то жирная кислота доставляется сюда в комплексе с альбумином. Этот комплекс образуется после работы липопротеинлипазы в других тканях.
Полиненасыщенные жирные кислоты (ПНЖК) обычно поступают в клетки в виде эфиров холестерола и в составе фосфолипидов ЛПВП и ЛПНП. Эти кислоты необходимы для синтеза эйкозаноидов в некоторых видах клеток, либо они участвуют в синтезе фосфолипидов для мембран клетки.
Голодание и мышечная работа
При голодании хиломикроны и ЛПОНП в крови отсутствуют. Так как данному состоянию обычно сопутствует гипогликемия, то для ее компенсации из поджелудочной железы секретируется глюкагон. Под влиянием глюкагона и других гормонов в жировых депо активируется расщепление ТАГ до жирных кислот и глицерола (липолиз). Транспорт жирных кислот, вышедших в кровь, осуществляется альбумином.
Всостоянии покоя, когда процессы пищеварения уже закончились, при длительном голодании (более 20 часов) и при физической нагрузке в большинстве клеток, кроме нейронов
иэритроцитов, жирные кислоты используются для образования энергии в процессах -окис- ления и ЦТК (50% и более всей энергии клетки).
Впечени при длительном голодании жирная кислота направляется в кетогенез на синтез кетоновых тел.
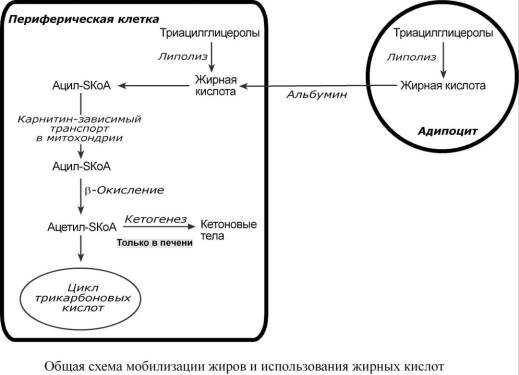
И С П О Л Ь З О В А Н И Е Т Р И А Ц И Л Г Л И Ц Е Р О Л О В
Распад ТАГ (липолиз, мобилизация жира) идет в жировых клетках постоянно и обычно существует равновесие между синтезом и распадом ТАГ. В состоянии покоя печень, сердце, скелетные мышцы и другие ткани (кроме эритроцитов и нейроцитов) более 50% энергии получают из окисления жирных кислот, поступающих из жировой ткани благодаря фоновому липолизу.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
245 |
|
|
|
|
|
Мобилизация жира активируется
oпри нормальных физиологических стрессовых ситуациях – эмоциональный стресс, мышечная работа, голодание,
oпри патологических состояниях – сахарный диабет I типа, другие гормональные заболевания: гиперкортицизм, гипертиреоз.
ОБЩАЯ ХАРАКТЕРИСТИКА МОБИЛИЗАЦИИ ЖИРОВ
Вцелом мобилизацию жира можно представить как последовательность следующих со-
бытий:
1.Липолиз – гормонзависимый распад ТАГ в жировой ткани (см ниже) или резервных ТАГ в самой клетке.
2.Транспорт жирных кислот из жировой ткани по крови в комплексе с альбумином.
3.Проникновение жирной кислоты в цитозоль клетки-мишени.
4.Активация жирной кислоты через присоединение HS-КоА.
5. Карнитин-зависимое перемещение жирной кислоты в митохондрию.
6.Окисление жирной кислоты с образованием ацетильных групп (ацетил-SКоА).
7.Сгорание ацетил-SКоА в цикле лимонной кислоты или синтез (только в печени) кетоновых тел.
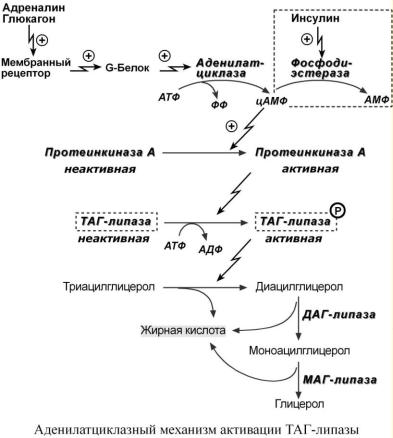
ГОРМОНЗАВИСИМАЯ АКТИВАЦИЯ ТАГ-ЛИПАЗЫ
Гормонзависимая активация ТАГ-липазы адипоцитов адреналином и глюкагоном происходит при напряжении организма (голодание, длительная мышечная работа, охлаждение). В первую очередь активность ТАГ-липазы зависит от соотношения инсулин/глюкагон.
Вцелом последовательность событий липолиза выглядит следующим образом:
1.Молекула гормона (адреналин, глюкагон, АКТГ, СТГ) взаимодействует со своим рецепто-
ром.
2.Активный гормон-рецепторный комплекс воздействует на мембранный G-белок.
3.G-белок активирует фермент аденилатциклазу.
Строение и обмен липидов |
246 |
|
|
4.Аденилатциклаза превращает АТФ в циклический АМФ (цАМФ) – вторичный посредник (мессенджер).
5.цАМФ аллостерически активирует фермент протеинкиназу А.
6.Протеинкиназа А фосфорилирует ТАГ-липазу и активирует ее.
7.ТАГ-липаза отщепляет от триацилглицеролов жирную кислоту в 1 или 3 положении с образованием ДАГ. Кроме ТАГ-липазы, в адипоцитах имеются еще ДАГ-липаза и МАГ-липаза, активность которых высока и постоянна, однако в покое она не проявляется из-за отсут-
ствия субстрата. Как только в клетке появляются ДАГ, начинает работать постоянно активная ДАГ-липаза, продукт ее реакции МАГ является субстратом для МАГ-липазы.
Кроме гормонов, влияющих на активность аденилатциклазы через G-белки, существуют иные механизмы регуляции. Например, соматотропный гормон увеличивает количество аденилатциклазы, глюкокортикоиды способствуют синтезу ТАГ-липазы.
Инсулин препятствует влиянию остальных гормонов на липолиз, т.к. он активирует фермент фосфодиэстеразу, которая гидролизует цАМФ и, следовательно, снижает активность ТАГ-липазы. Также он активирует протеинфосфатазы, дефосфорилирующие ТАГ-липазу.
Т Р А Н С П О Р Т Ж И Р Н Ы Х К И С Л О Т И Г Л И Ц Е Р О Л А
В результате липолиза в адипоцитах образуются свободный глицерол и жирные кислоты. Глицерол с кровью доставляется в печень и почки, здесь фосфорилируется и окисляется в метаболит гликолиза диоксиацетонфосфат. В зависимости от условий ДОАФ может вклю-
чаться в реакции глюконеогенеза (при голодании, мышечной нагрузке) или окисляться в гликолизе до пировиноградной кислоты.
Жирные кислоты транспортируются в крови в комплексе с альбуминами плазмы: o при физической нагрузке – в мышцы,
oв обычных условиях и при голодании – в мышцы и большинство тканей, однако при этом около 30% жирных кислот захватывается печенью.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
247 |
|
|
|
|
|
При голодании и физической нагрузке после проникновения в клетки жирные кислоты вступают на путь β-окисления.
- О К И С Л Е Н И Е Ж И Р Н Ы Х К И С Л О Т
Реакции β-окисления происходят в митохондриях большинства клеток организма (кроме нервных клеток). Для окисления используются жирные кислоты, поступающие в цитозоль из крови или появляющиеся при внутриклеточном липолизе ТАГ.
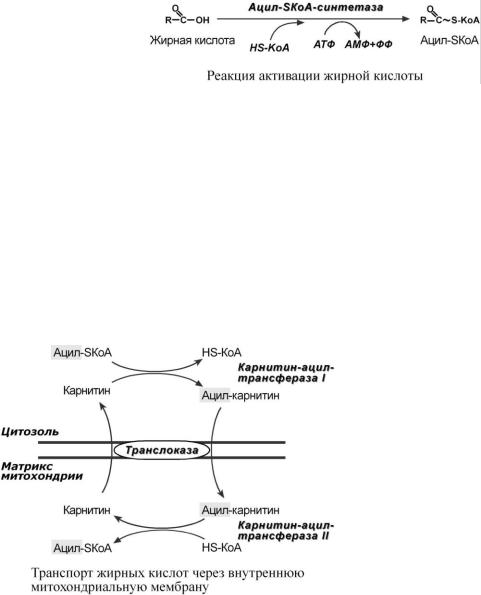
1. Прежде, чем проникнуть в матрикс митохондрий и окислиться, жирная кислота в цитозоле должна активироваться. Это осу-
ществляется присоединением коэнзима А с образованием ацил-SКоА. Ацил-SКоА является высокоэнергетическим соединением. Необратимость реакции достигается гидролизом дифосфата на две молекулы фосфорной кислоты.
2.Ацил-SКоА не способен проходить через митохондриальную мембрану, поэтому существует способ переноса жирной кислоты в комплексе с витаминоподобным веществом карнитином. На наружной мембране митохондрий имеется фермент карнитин-аци-
лтрансфераза I.
3.После связывания с карнитином жирная кислота переносится через мембрану транслоказой. Здесь на внутренней стороне мембраны фермент карнитин-ацилтрансфераза II вновь образует ацил-SКоА который вступает на путь β-окисления.
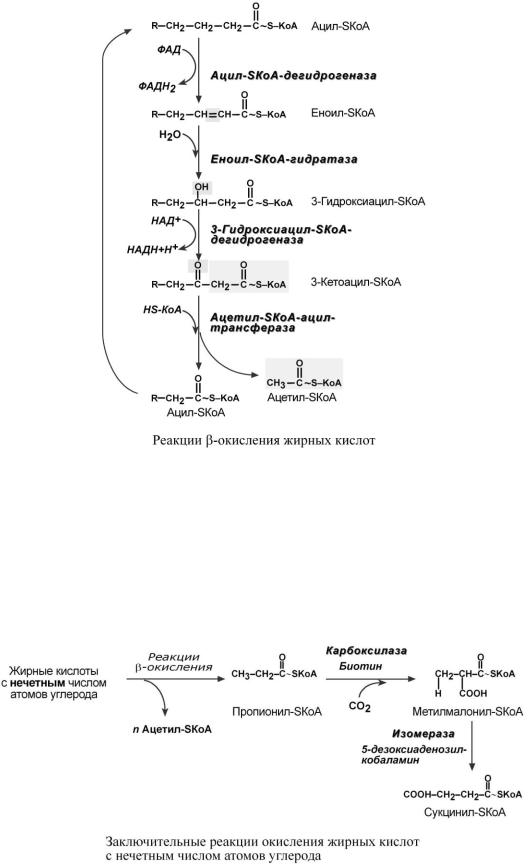
4.Процесс собственно β-окисления состоит из 4-х реакций, повторяющихся циклически. В них последовательно происходит окисление, гидратирование и вновь окисление 3-го атома углерода (β-положение). В последней реакции от жирной кислоты отщепляется ацетилSКоА. Оставшаяся укороченная на два углерода жирная кислота возвращается к первой реакции и все повторяется снова, до тех пор, пока в последнем цикле не образуются два ацетил-SКоА.
Строение и обмен липидов |
248 |
|
|
Окисление жирных кислот с нечетным числом углеродных атомов
Жирные кислоты с нечетным числом углеродов поступают в организм с растительной пищей и морепродуктами. Их β-окисление происходит по обычному пути до последней реакции, в которой образуется пропионил-SКоА. Суть превращений пропионил-SКоА сводится к его карбоксилированию, изомеризации и образованию сукцинил-SКоА, используемому в ЦТК. В этих реакциях участвуют биотин (витамин H) и дезоксиаденозилкобаламин (витамин В12).
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
249 |
|
|
|
|
|
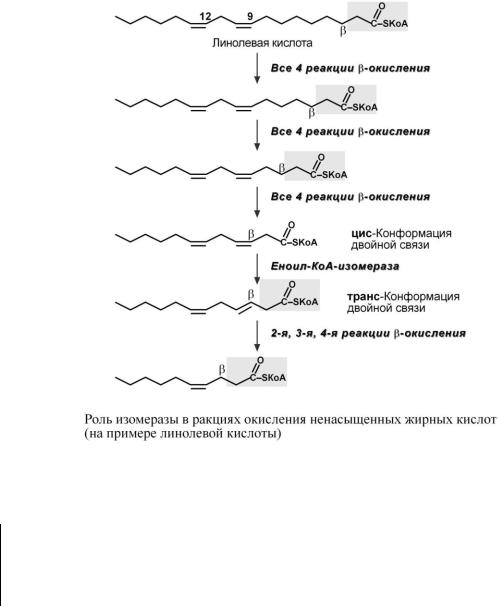
Окисление ненасыщенных жирных кислот
При окислении ненасыщенных жирных кислот возникает потребность клетки в дополнительных ферментах изомеразах. Эти изомеразы перемещают двойные связи в жирнокислотных остатках из - в β-положение, и переводят природные двойные связи из цис- в транс- положение.
Таким образом, уже имеющаяся двойная связь готовится к β-окислению и пропускается первая реакция цикла, в которой участвует ФАД.
Расчет энергетической ценности и коэффициента Р/О
при -окислении жирных кислот
Ранее при расчете эффективности окисления коэффициент P/O для НАДH принимался равным 3,0, для ФАДH2 – 2,0.
По современным данным значение коэффициента P/O для НАДH соответствует
2,5, для ФАДH2 – 1,5.
При расчете количества АТФ, образуемого при β-окислении жирных кислот необходимо учитывать:
oколичество образуемого ацетил-SКоА – определяется обычным делением числа атомов углерода в жирной кислоте на 2.
oчисло циклов β-окисления. Число циклов β-окисления легко определить исходя из представления о жирной кислоте как о цепочке двухуглеродных звеньев. Число разрывов между звеньями соответствует числу циклов β-окисления. Эту же величину можно под-
считать по формуле (n/2 -1), где n – число атомов углерода в кислоте.
oналичие двойных связей в жирной кислоте. В первой реакции β-окисления происходит образование двойной связи при участии ФАД. Если двойная связь в жирной кислоте уже имеется, то необходимость в этой реакции отпадает и ФАДН2 не образуется. Количество