

Лекции по общей биохимии (О. Тимин)
.pdfМатричные биосинтезы |
170 |
|
|
o субстрат для синтеза – рибонуклеотиды (УТФ, ГТФ, ЦТФ, АТФ), o источник энергии – УТФ, ГТФ, ЦТФ, АТФ.
o ферменты РНК-полимеразы и белковые факторы транскрипции.
Биосинтез РНК происходит в участке ДНК, который называется транскриптон, с одного края он ограничен промотором (начало), с другого – терминатором (конец).
РНК-полимеразы эукариот имеют по две больших субъединицы и несколько малых субъединиц.
СТАДИИ ТРАНСКРИПЦИИ
Выделяют три стадии транскрипции: инициация, элонгация и терминация.
Инициация
Промотор содержит стартовый сигнал транскрипции – ТАТА-бокс – определенную последовательность нуклеотидов ДНК, связывающую первый фактор инициации ТАТА-фактор. Этот ТАТА-фактор обеспечивает присоединение РНК-полимеразы к той нити ДНК, которая будет использоваться в качестве шаблона для транскрипции (матричная нить ДНК). Так как промотор ассиметричен ("ТАТА"), то он связывает РНК-полимеразу только в одной ориентации, что определяет направление транскрипции от 5'-конца к 3'-концу (5' 3'). Для связывания РНК-полимеразы с промотором необходим еще один фактор инициации – -фактор (греч. – "сигма"), но сразу после синтеза затравочного фрагмента РНК (длиной 8-10 рибонуклеотидов) -фактор отрывается от фермента.
Другие факторы инициации раскручивают спираль ДНК перед РНК-полимеразой.
Элонгация
Белковые факторы элонгации обеспечивают продвижение РНК-полимеразы вдоль ДНК и расплетают молекулу ДНК на протяжении примерно 17 нуклеотидных пар. РНК-полимераза продвигается со скоростью 40-50 нуклеотидов в секунду в направлении 5' 3'. Фермент использует АТФ, ГТФ, ЦТФ, УТФ одновременно в качестве субстрата и в качестве источника энергии.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
171 |
|
|
|
|
|
Терминация
РНК-полимераза остановится, когда достигнет терминирующих кодонов. С помощью белкового фактора терминации, так называемого -фактора (греч. – "ро"), от матрицы ДНК отделяются фермент и синтезированная молекула РНК, которая является первичным транскриптом, предшественником мРНК или тРНК или рРНК.
П Р О Ц Е С С И Н Г Р Н К
Сразу после синтеза первичные транскрипты РНК по разным причинам еще не имеют активности, являются "незрелыми" и в дальнейшем претерпевают ряд изменений, которые называются процессинг. У эукариот процессингу подвергаются все виды пре-РНК, у прокариот – только предшественники рРНК и тРНК.
ПРОЦЕССИНГ ПРЕДШЕСТВЕ ННИКА МРНК
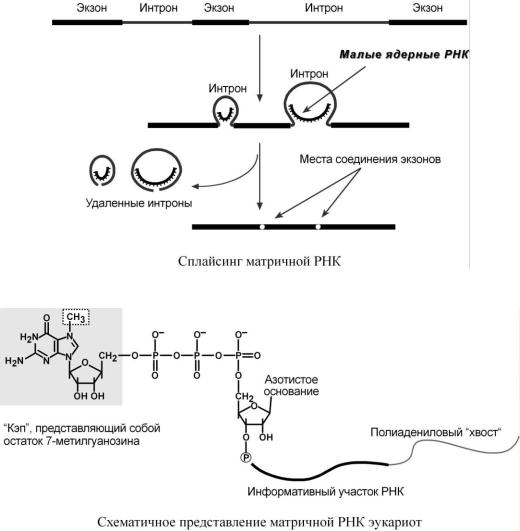
При транскрипции участков ДНК, несущих информацию о белках, образуются гетерогенные ядерные РНК, по размеру намного превосходящие мРНК. Дело в том, что из-за мозаичной структуры генов эти гетерогенные РНК включают в себя информативные (экзоны) и неинформативные (интроны) участки.
1. Сплайсинг (англ. splice - склеивать встык) – особый процесс, в котором при участии малых ядерных РНК происходит удаление интронов и сохранение экзонов.
2. Кэпирование (англ. cap – шапка) – происходит еще во время транскрипции. Процесс состоит в присоединении к 5'-трифосфату концевого нуклеотида пре-мРНК 5'-углерода
Матричные биосинтезы |
172 |
N7-метил-гуанозина. "Кэп" необходим для защиты молекулы РНК от экзонуклеаз, работающих с 5'-конца, а также для связывания мРНК с рибосомой и для начала трансляции.
3. Полиаденилирование – при помощи полиаденилат-полимеразы с использованием молекул АТФ происходит присоединение к 3'-концу РНК от 100 до 200 адениловых нуклеотидов, формирующих поли (А)-хвост. Поли (А)-хвост необходим для защиты молекулы РНК от экзонуклеаз, работающих с 3'-конца.
ПРОЦЕССИНГ ПРЕДШЕСТВЕ ННИКА Р РНК
Предшественники рРНК являются более крупными молекулами по сравнению со зрелыми рРНК. Их созревание сводится к разрезанию прерибосомной РНК на более мелкие формы, которые уже непосредственно участвуют в формировании рибосомы. У эукариот существуют 5S-, 5,8S-, 18S-, и 28S-рРНК. При этом 5S-рРНК синтезируется отдельно, а большая прерибосомная 45S-РНК расщепляется специфичными нуклеазами с образованием 5,8S-рРНК, 18S-рРНК, и 28S-рРНК.
Упрокариот молекулы рибосомальной РНК совсем иные по своим свойствам (5S-, 16S-
,23S-рРНК), что является основой изобретения и использования ряда антибиотиков в медицине.
ПРОЦЕССИНГ ПРЕДШЕСТВЕ ННИКА ТРНК
1.Формирование на 3'-конце последовательности Ц-Ц-А. Для этого у одних пре-тРНК
с3'-конца удаляются лишние нуклеотиды до "обнажения" триплета Ц-Ц-А, у других идет присоединение этой последовательности.
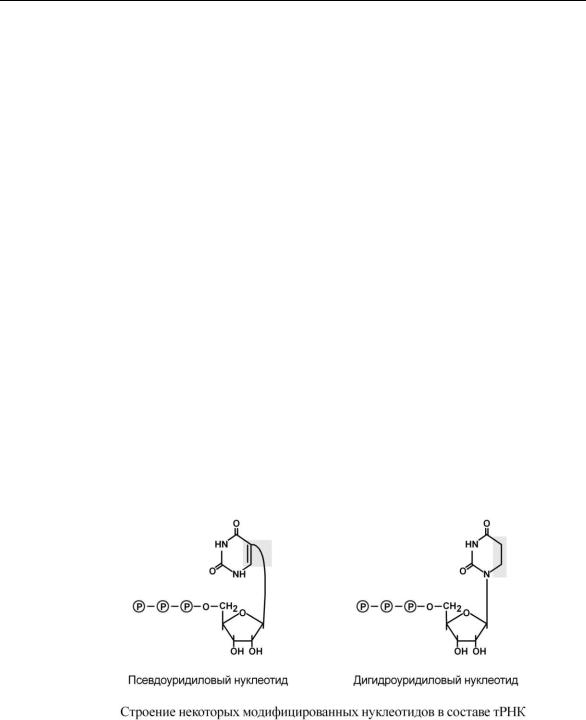
2.Формирование антикодоновой петли происходит путем сплайсинга и удаления интрона в средней части пре-тРНК.
3.Модификация нуклеотидов в молекуле путем дезаминирования, метилирования, восстановления. Например, образование псевдоуридина и дигидроуридина.
РЕГУЛЯЦИЯ ТРАНСКРИПЦИИ
Так как транскрипция связывает ядро – "мозг" клетки, ее "банк знаний" и белки, "рабочих лошадок" клетки, то от качества и активности транскрипции зависит объем синтеза тех или иных белков, жизнедеятельность клетки, ее способность адаптироваться к окружающей обстановке.
У прокариот и эукариот регуляция транскрипции происходит, естественно, по-разному, хотя некоторые моменты похожи.
Прокариоты
Регуляция биосинтеза белка у прокариот осуществляется на уровне транскрипции мРНК. В настоящее время принята теория оперона, сформулированная Франсуа Жакобом и Жаком Моно. В основе теории лежат следующие понятия:
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
173 |
|
|
|
|
|
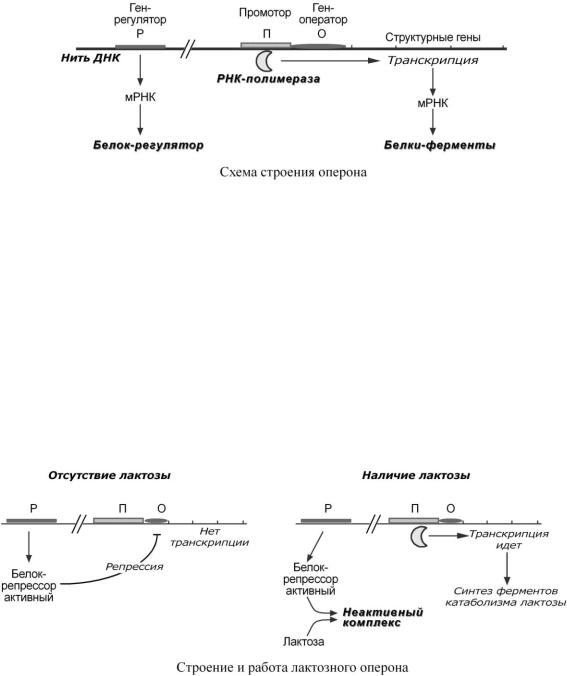
oоперон – группа тесно связанных между собой генов, которые программируют образование структурных белков и ферментов в клетке,
oконституитивные гены – те, которые активны в клетках всегда, независимо от ее активности,
o |
индуцибельные гены – те, которые считываются при необходимости, |
o |
ген-регулятор – ген, регулирующий работу оперона, но не входящий в его состав. Он |
|
синтезирует белок-регулятор (чаще называемый белок-репрессор), который может быть |
|
в активной или неактивной форме, |
oген-оператор – участок ДНК, способный связываться с белком-регулятором, и "решающий" нужно работать РНК-полимеразе или нет.
Лактозный оперон
Лактозный оперон в целом отвечает за катаболизм лактозы.
При изучении E.coli было замечено, что в клетке может быть две взаимоисключающие ситуации:
oактивность одного из ферментов катаболизма лактозы низка, если в среде имеется глю-
коза.
oно активность этого фермента резко повышается в обратной ситуации, т.е. при отсут-
ствии глюкозы и при наличии лактозы.
На основании этих наблюдений была предложена схема регуляции оперона по меха-
низму индукции:
1.В отсутствие лактозы активный белок-репрессор связывается с оператором и блокирует синтез мРНК, кодирующей ферменты катаболизма лактозы. В результате эти ферменты не образуются.
Матричные биосинтезы |
174 |
|
|
2. Если глюкозы нет, а лактоза есть, то последняя связывается с белком-репрессором и ингибирует его, не давая связаться с геном-оператором. Это позволяет РНК-полимеразе считывать информацию, отвечающую за синтез ферментов катаболизма лактозы, и синтезировать мРНК.
Таким образом, лактоза является индуктором транскрипции.
Триптофановый оперон
Триптофановый оперон в целом отвечает за синтез триптофана.
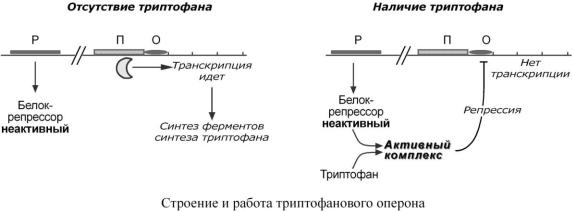
Функционирование триптофанового оперона в некотором смысле противоположно лактозному. Регуляция осуществляется по механизму репрессии.
1.В отличие от лактозного оперона, белок-репрессор синтез ируется в неактивном состоянии
ине может заблокировать транскрипцию генов, кодирующих ферменты синтеза триптофана. Синтез этой аминокислоты будет в клетке продолжаться до тех пор, пока в питательной среде не появится триптофан.
2.Триптофан соединяется с белком-репрессором и активирует его. Далее такой активный комплекс присоединяется к гену-оператору и блокирует транскрипцию. Таким образом, при наличии триптофана в среде прекращается его внутриклеточный синтез, экономятся ресурсы и энергия бактериальной клетки.
Вэтом случае триптофан является репрессором транскрипции.
Эукариоты
Внутриклеточная регуляция
Существенное усложнение эукариотических организмов повлекло за собой появление новых способов регуляции активности транскрипции.
Амплификация – это увеличение количества генов, точнее многократное копирование одного гена. Естественно, все полученные копии равнозначны и одинаково активно обеспечивают транскрипцию.
Энхансеры (англ. to enhance - усиливать) – это участки ДНК в 10-20 пар оснований, способные значительно усиливать экспрессию генов той же ДНК. В отличие от промоторов они значительно удалены от транскрипционного участка и могут располагаться от него в любом направлении (к 5'-концу или к 3'-концу). Сами энхансеры не кодируют какие-либо белки, но способны связываться с регуляторными белками (подавляющими транскрипцию).
Сайленсеры (англ. silence – молчание) – участки ДНК, в принципе схожие с энхансерами, но они способны замедлять транскрипцию генов, связываясь с регуляторными белками (которые ее активируют).
Перестройка генов. К подобным процессам относится кроссинговер – обмен участками гомологичных хромосом, и более сложный процесс – сайт-специфичная рекомбинация, которая изменяет положение и порядок нуклеотидных последовательностей в геноме.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
175 |
|
|
|
|
|
Процессинг мРНК – некоторые пре-мРНК подвергаются разным вариантам сплайсинга (альтернативный сплайсинг) в результате чего образуются разные мРНК, и соответственно, белки с разной функцией.
Изменение стабильности мРНК – чем выше продолжительность жизни мРНК в цитозоле клетки, тем больше синтезируется соответствующего белка.
Лекарственная регуляция
Ингибирование
1.Гетероциклические соединения доксорубицин, дауномицин и актиномицин D обла-
дают способностью интеркалировать (встраиваться между нитей молекулы ДНК) между двумя соседними парами оснований Г-Ц. В результате возникает препятствие для движения РНК-полимеразы ("заедание молнии") и остановка транскрипции.
2.Рифампицин связывается с -субъединицей РНК-полимеразы прокариот и ингибирует ее. Благодаря такой избирательности действия рифампицин действует только на бактерии
иявляется препаратом для лечения туберкулеза.
3.-Аманитин, октапептид бледной поганки (Amanita phalloides) блокирует РНК-полимеразу II эукариот и предотвращает продукцию мРНК.
Активация
Активация транскрипции используется в клинике намного реже и заключается в применении аналогов стероидных гормонов для достижения анаболического эффекта в органе-ми- шени (см "Гормоны"/"Механизм действия стероидных гормонов").
Г Е Н Е Т И Ч Е С К И Й К О Д
Генетический (биологический) код – это способ перевода четырехзначного (А, Г, У, Ц) языка нуклеотидов в двадцатизначный язык аминокислотной последовательности.
Свойства биологического кода
Триплетность – три нуклеотида формируют кодон, кодирующий аминокислоту. Всего насчитывают 61 смысловой кодон.
Специфичность (или однозначность) – каждому кодону соответствует только одна аминокислота.
Вырожденность – одной аминокислоте может соответствовать несколько кодонов. Универсальность – биологический код одинаков для всех видов организмов на Земле
(однако в митохондриях млекопитающих есть исключения).
Колинеарность – последовательность кодонов соответствует последовательности аминокислот в кодируемом белке.
Неперекрываемость – триплеты не накладываются друг на друга, располагаясь рядом. Отсутствие знаков препинания – между триплетами нет дополнительных нуклеотидов
или каких-либо иных сигналов.
Однонаправленность – при синтезе белка считывание кодонов идет последовательно, без пропусков или возвратов назад.
А Д А П Т О Р Н А Я Р О Л Ь Т Р А Н С П О Р Т Н Ы Х Р Н К
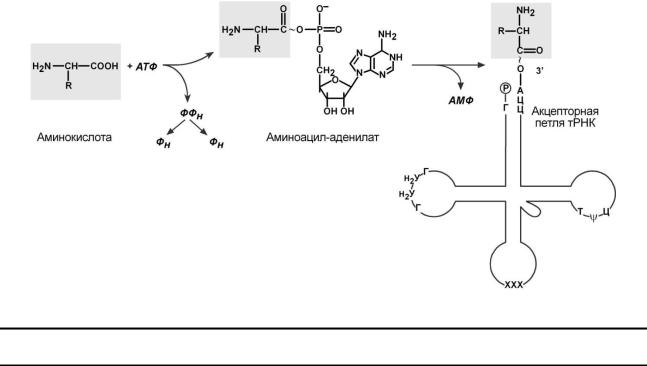
Транспортные РНК являются единственным посредником между 4-х буквенной последовательностью нуклеиновых кислот и 20-ти буквенной последовательностью белков. Именно от наличия того или иного антикодона в тРНК зависит, какая аминокислота включится в белковую молекулу, т.к. ни рибосома, ни мРНК не узнают аминокислоту. Таким образом, адапторная роль тРНК заключается: 1) в специфичном связывании с аминокислотами, 2) во включении аминокислот в белковую цепь в соответствии с матрицей мРНК.
Избирательное присоединение аминокислоты к тРНК осуществляется ферментом ами- ноацил-тРНК-синтетазой, имеющей специфичность одновременно к двум соединениям: ка-
Матричные биосинтезы |
176 |
|
|
кой-либо аминокислоте и соответствующей ей тРНК. Для реакции требуется две макроэргические связи АТФ. Аминокислота присоединяется к 3'-концу акцепторной петли тРНК через-карбоксильную группу, и связь между аминокислотой и тРНК становится макроэргической. -Аминогруппа остается свободной.
Так как существует около 60 различных тРНК, то некоторым аминокислотам соответствует по две или более тРНК. Различные тРНК, присоединяющие одну аминокислоту, назы-
вают изоакцепторными.
СИН ТЕ З БЕ ЛК А – ТР АН С ЛЯ Ц ИЯ
Трансляция (англ. translation – перевод) – это биосинтез белка на матрице мРНК. После переноса информации с ДНК на матричную РНК начинается синтез белков. Каж-
дая зрелая мРНК несет информацию только об одной полипептидной цепи. Если клетке необходимы другие белки, то необходимо транскрибировать мРНК с иных участков ДНК.
Биосинтез белков или трансляция происходит на рибосомах, внутриклеточных белоксинтезирующих органеллах, и включает 5 ключевых элементов:
o матрица – матричная РНК,
o растущая цепь – полипептид,
o субстрат для синтеза – 20 протеиногенных аминокислот, o источник энергии – ГТФ,
o рибосомальные белки, рРНК и белковые факторы.
Выделяют три основных стадии трансляции: инициация, элонгация, терминация.
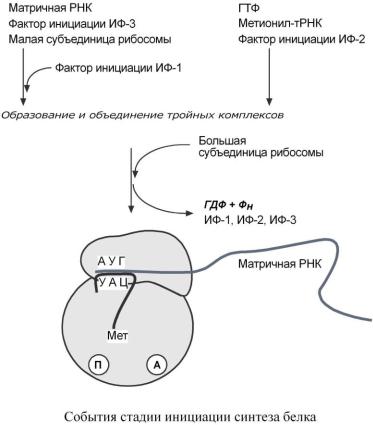
ИНИЦИАЦИЯ
Для инициации необходимы мРНК, ГТФ, малая и большая субъединицы рибосомы, три белковых фактора инициации (ИФ-1, ИФ-2, ИФ-3), метионин и тРНК для метионина.
В начале этой стадии формируются два тройных комплекса: o первый комплекс – мРНК + малая субъединица + ИФ-3,
o второй комплекс – метионил-тРНК + ИФ-2 + ГТФ.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
177 |
|
|
|
|
|
После формирования тройные комплексы объединяются с большой субъединицей рибосомы. В этом процессе активно участвуют белковые факторы инициации, источником энергии служит ГТФ. После сборки комплекса инициирующая метионил-тРНК связывается с первым кодоном АУГ матричной РНК и располагается в П-центре (пептидильный центр) большой субъединицы. А-центр (аминоацильный центр) остается свободным, он будет задействован на стадии элонгации для связывания аминоацил-тРНК.
После присоединения большой субъединицы начинается стадия элонгации.
ЭЛОНГ АЦИЯ
Для этой стадии необходимы все 20 аминокислот, тРНК для всех аминокислот, белковые факторы элонгации, ГТФ. Удлинение цепи происходит со скоростью примерно 20 аминокислот в секунду.
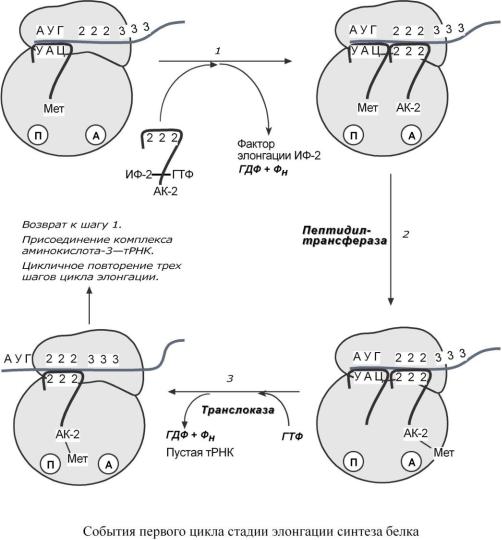
Элонгация представляет собой циклический процесс. Второй цикл (и следующие циклы) элонгации включает три шага:
1.Присоединение аминоацил-тРНК (еще второй) к кодону мРНК (еще второму) – аминокислота при этом встраивается в А-центр рибосомы. Источником энергии служит ГТФ.
2.Фермент пептидилтрансфераза осуществляет перенос метионина с метионил-тРНК (из П-центра) на вторую аминоацил-тРНК (в А-центре) с образованием пептидной связи между метионином и второй аминокислотой. При этом уже активированная СООН-группа метионина связывается со свободной NH2-группой второй аминокислоты. Здесь источником энергии служит макроэргическая связь между аминокислотой и тРНК.
Матричные биосинтезы |
178 |
|
|
3.Фермент транслоказа перемещает мРНК относительно рибосомы таким образом, что первый кодон АУГ оказывается вне рибосомы, второй кодон становится напротив П-центра, напротив А-центра оказывается третий кодон. Для этих процессов необходима затрата энергии ГТФ. Так как вместе с мРНК перемещаются закрепленные на ней тРНК, то инициирующая первая тРНК выходит из рибосомы, вторая тРНК с дипептидом помещается в П-центр.
4.Второе повторение цикла – начинается с присоединения третьей аминоацил-тРНК к третьему кодону мРНК, аминокислота-3 становится в А-центр. Далее трансферазная реакции повторяется и образуется трипептид, занимающий А-центр, после чего он смещается в П- центр в транслоказной реакции.
5.В пустой А-центр входит четвертая аминоацил-тРНК и все повторяется.
Цикл элонгации (реакции 1,2,3) повторяется столько раз, сколько аминокислот необходимо включить в полипептидную цепь.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
179 |
|
|
|
|
|
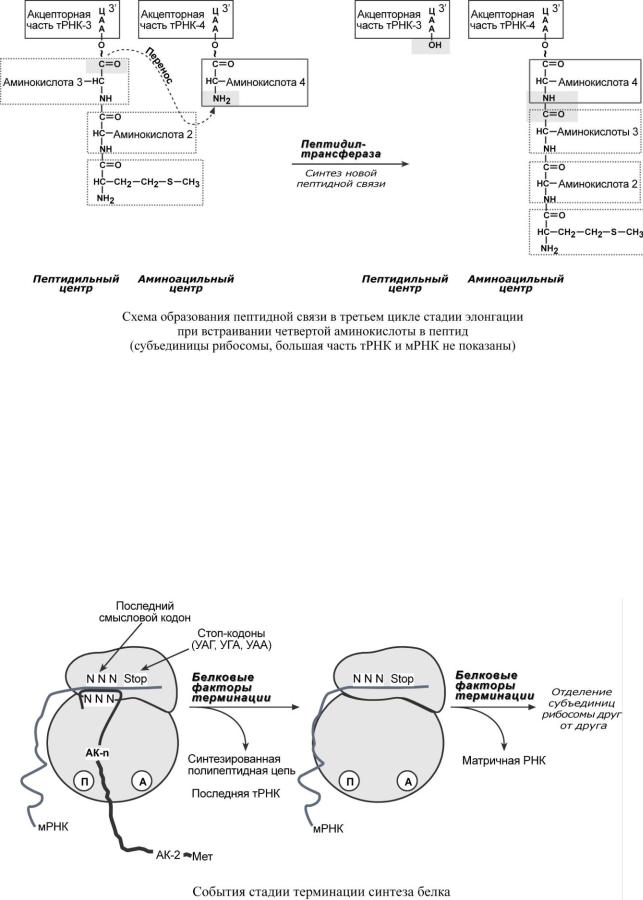
ТЕ Р МИНАЦИЯ
Синтез белка продолжается до тех пор, пока рибосома не достигнет на мРНК особых терминирующих кодонов – стоп-кодонов УАА, УАГ, УГА. Данные триплеты не кодируют ни одной из аминокислот, их также называют нонсенс-кодоны. При вхождении этих кодонов внутрь рибосомы происходит активация белковых факторов терминации, которые последовательно катализируют:
1.Гидролитическое отщепление полипептида от конечной тРНК.
2.Отделение от П-центра последней, уже пустой, тРНК.
3.Диссоциацию рибосомы.
Источником энергии для завершения трансляции является ГТФ.