

Лекции по общей биохимии (О. Тимин)
.pdfОбмен аминокислот и белков |
110 |
|
|
Кетогенными являются лизин и лейцин, при их окислении образуется только ацетилSКоА. Он в состоянии принять участие в синтезе кетоновых тел, жирных кислот и холестерола.
Также выделяют небольшую группу смешанных аминокислот, из них образуется пируват, метаболиты ЦТК и ацетил-SКоА (фенилаланин, тирозин, изолейцин, триптофан).
При направлении аминокислот на катаболизм пути их обмена сходятся к 6 продуктам, которые вступают в ЦТК и полностью окисляются до углекислого газа и воды с выделением энергии. Из общего количества энергии, образующейся в организме, на долю аминокислот приходится около 10%.
РАСЧЕТ ЭНЕРГЕТИЧЕСКОЙ ЦЕННОСТИ И КОЭФФИЦИЕНТА Р/О ПРИ ОКИСЛЕНИИ НЕКОТО РЫХ АМИНОКИСЛОТ
Ранее при расчете эффективности окисления коэффициент P/O для НАДH принимался равным 3,0, для ФАДH2 – 2,0.
По современным данным значение коэффициента P/O для НАДH соответствует 2,5, для ФАДH2 – 1,5.
При расчете энергетической ценности, т.е. количества АТФ, образующейся при полном окислении аминокислот, и коэффициента Р/О необходимо представлять себе весь путь аминокислоты до полного окисления ее углеродных атомов в СО2. При этом необходимо учитывать
число атомов углерода в молекуле.
Например, аспартат содержит 4 атома углерода и поэтому его остатку необходимо пройти 2 оборота ЦТК, прежде чем все они выделятся в виде СО2. Подсчитывая число восстановленных НАДН, ФАДН2 и ГТФ, образуемых в двух оборотах ЦТК, определяем сумму АТФ – 20 молекул. При расчете коэффициента Р/О учитываем только фосфат, включенный в АТФ ферментом АТФ-синтазой, т.е. в процессе окислительного фосфорилирования. Это значит, что фосфат, входящий в ГТФ, не учитывается!
И, наконец, помним, что каждая молекула восстановленного эквивалента (т.е. НАДН или ФАДН2) передает на дыхательную цепь по одной паре электронов, которые проходят разное расстояние: от НАДН – три комплекса ферментов, от ФАДН2 – 2 комплекса ферментов, но в любом случае восстанавливают до воды по одному атому кислорода.
Схожим образом рассчитываем требуемые значения суммы АТФ и коэффициента Р/О для аланина и глутамата.

biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
111 |
|
|
|
|
|
Два атома углерода аланина попадают в ЦТК после того, как безазотистый остаток аланина – пируват – окислится в пируват-дегидрогеназном комплексе с образованием НАДН. Образованный ацетил-SКоА входит в ЦТК и полностью окисляется в одном его обороте. В результате в дыхательную цепь направляется 4 молекулы НАДН и 1 молекула ФАДН2.

Обмен аминокислот и белков |
112 |
В случае с глутаматом надо учесть, что -кетоглутарат, образуемый из глутаминовой кислоты, является метаболитом цикла Кребса. Он сразу превращается в сукцинил-SКоА и далее, по реакциям ЦТК, в оксалоацетат. Для окисления оксалоацетата требуется два оборота ЦТК (см выше).
П Р Е В Р А Щ Е Н И Е А М И Н О К И С Л О Т П О К А Р Б О К С И Л Ь Н О Й Г Р У П П Е
Такое превращение связано с удалением карбоксильной группы от аминокислоты и об-
разованием биогенных аминов.
ГИСТАМИН
Реакция образования гистамина наиболее активно идет в тучных клетках легких, кожи, печени, в базофилах и эозинофилах. В них гистамин синтезируется и накапливается в секреторных гранулах.
В кровь гистамин выделяется при повреждении ткани, при ударе, при электрическом раздражении. В клинической практике секреция гистамина обычно связана с аллергиями – при повторном попадании антигена в ранее сенсибилизированный организм развивается аллергическая реакция.
Физиологические эффекты
1.Расширение артериол и капилляров и, как следствие, покраснение кожи, снижение артериального давления;
2.Повышение проницаемости стенки капилляров и, как следствие, выход жидкости в межклеточное пространство (отечность), снижение артериального давления;
3.Если п.п.1 и 2 наблюдаются в головном мозге – повышение внутричерепного давления;
4.Увеличивает тонус гладких мышц бронхов, как следствие – спазм и удушье;
5.Слабо повышает тонус мышц желудочно-кишечного тракта;
6.Стимулирует секрецию слюны и соляной кислоты желудочного сока.
СЕ Р ОТОНИН
Серотонин активно синтезируется в тучных клетках кожи, легких, печени, в селезенке,
ЦНС.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
113 |
Физиологические эффекты
1.Стимулирует сокращение гладких мышц желудочно-кишечного тракта и, как следствие, повышение перистальтики ЖКТ.
2.Выражено стимулирует сокращение гладких мышц кровеносных сосудов в тканях, кроме миокарда и скелетных мышц, и, как следствие, повышение артериального давления.
3.Слабо увеличивает тонус гладких мышц бронхов.
4.В центральной нервной системе является тормозным медиатором.
5.В периферических нервных окончаниях обусловливает возникновение боли и зуда (например, при укусе насекомых).
ГАММА -АМИНОМАСЛЯНАЯ КИСЛОТА
Синтез -аминомасляной кислоты (ГАМК) происходит исключительно в центральной нервной системе – в подкорковых образованиях головного мозга.
Физиологические эффекты
В центральной нервной системе ГАМК (наряду с глутаминовой кислотой) является тормозным медиатором. Наиболее высока ее роль в височной и лобной коре, гиппокампе, миндалевидных и гипоталамических ядрах, черной субстанции, ядрах мозжечка.
ДОФАМИН
Синтез дофамина происходит в основном в нейронах промежуточного и среднего мозга.
Физиологические эффекты
Является медиатором дофаминовых рецепторов в подкорковых образованиях ЦНС, в больших дозах расширяет сосуды сердца, стимулирует частоту и силу сердечных сокращений, расширяет сосуды почек, увеличивая диурез.
ОБЕЗВРЕЖИВАНИЕ БИОГЕН НЫХ АМИНОВ
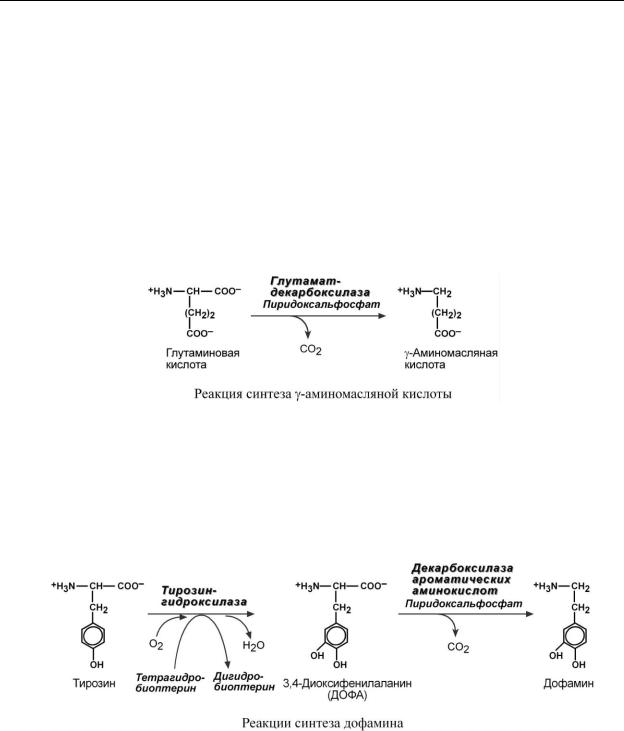
Существуют два типа реакций инактивация биогенных аминов – дезаминирование и метилирование.
Дезаминирование протекает с образованием свободного аммиака и с участием ФАД.
Катализирует реакцию моноаминоксидаза, она обнаружена во многих тканях, но наиболее активна в печени, желудке, почках, кишечнике, нервной ткани.
Обмен аминокислот и белков |
114 |
|
|
Метилирование биогенного амина происходит при наличии у него гидроксильной группы (дофамин, серотонин). В реакции принимает участие активная форма метионина – S-аденозилметионин (SAM), образуется метилированная форма амина и S-аденозилгомоци- стеин (SАГ).
П Р Е В Р А Щ Е Н И Е А М И Н О К И С Л О Т С У Ч А С Т И Е М А М И Н О Г Р У П П Ы
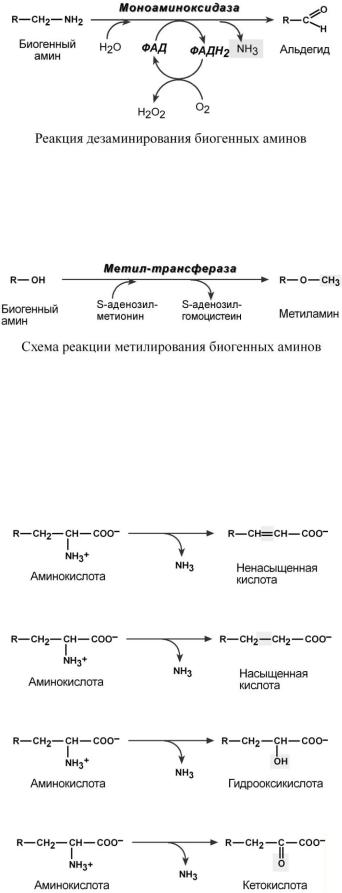
Превращение аминокислот с участием NH2-группы сводится к ее отщеплению от углеродного скелета – происходят реакции дезаминирования.
ТИПЫ ДЕЗАМИНИРОВАНИЯ
1.Внутримолекулярное – с образованием ненасыщенной жирной кислоты,
2.Восстановительное – с образованием насыщенной жирной кислоты,
3.Гидролитическое – с образованием карбоновой гидроксикислоты,
4.Окислительное – с образованием кетокислот.
Окислительное дезаминирование является основным путем катаболизма большинства аминокислот. Однако гистидин теряет аминогруппу с использованием внутримолекулярного

biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
115 |
|
|
|
|
|
дезаминирования, а треонин и серин сразу подвергаются прямому расщеплению до глицина и ацетальдегида (треонин) или гидроксиметила (серин).
ОКИСЛИТЕЛЬНОЕ ДЕЗАМИН ИРОВАНИЕ
Выделяют два варианта окислительного дезаминирования: прямое и непрямое.
Прямое окислительное дезаминирование
Прямое дезаминирование катализируется одним ферментом, в результате образуется NH3 и кетокислота. Прямое окислительное дезаминирование может идти в присутствии кислорода (аэробное) и не нуждаться в кислороде (анаэробное).
1. Аэробное прямое окислительное дезаминирование катализируется оксидазами D-аминокислот (D-оксидазы) в качестве кофермента использующими ФАД, и оксидазами L-аминокислот (L-оксидазы) с коферментом ФМН.
Наибольшая активность L-оксидаз обнаружена в печени и в почках, но роль их не совсем ясна, т.к. их оптимум рН находится около 10 и при внутриклеточных рН ферменты почти не активны.
Оксидазы D-аминокислот, имеющих только бактериальное происхождение, снижают их количество в тканях. Это модулирует обнаруженное недавно влияние D- аминокислот на активность эндокринных желез и ЦНС.
2. Анаэробное прямое окислительное дезаминирование существует только для глутаминовой кислоты, катализируется только глутаматдегидрогеназой, превращающей глутамат в-кетоглутарат. Фермент глутаматдегидрогеназа имеется в митохондриях всех клеток организма (кроме мышечных).
Этот тип дезаминирования теснейшим образом связан с трансаминированием аминокислот (см ниже) и формирует с ним процесс трансдезаминирования (см ниже).
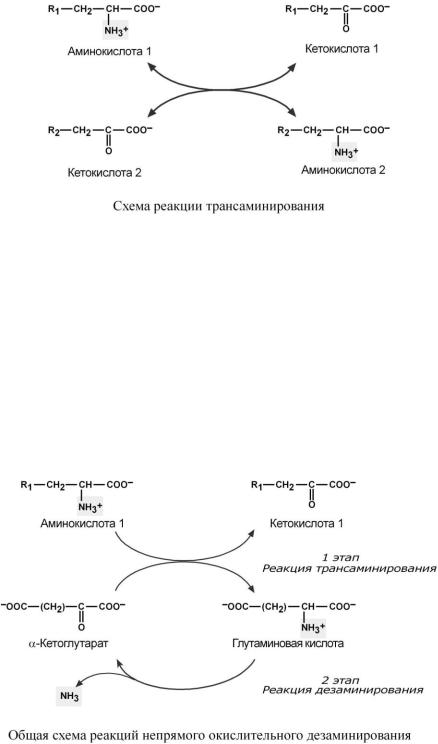
Непрямое окислительное дезаминирование (трансдезаминирование)
Непрямое окислительное дезаминирование включает 2 этапа и активно идет во всех клетках организма.
Обмен аминокислот и белков |
116 |
|
|
Первый этап заключается в обратимом переносе NH2-группы с аминокислоты на кетокислоту с образованием новой аминокислоты и новой кетокислоты – этот перенос называется трансаминирование (механизм реакции см ниже).
Вкачестве кетокислоты-акцептора ("кетокислота 2") в организме обычно используется-кетоглутаровая кислота, которая превращается в глутамат.
Врезультате трансаминирования свободные аминокислоты теряют -NH2-группы и превращаются в соответствующие кетокислоты. Далее их кетоскелет катаболизирует специфическими путями и вовлекается в цикл трикарбоновых кислот и тканевое дыхание, где сгорает до СО2 и Н2О. При необходимости (например, голодание) углеродный скелет глюкогенных аминокислот может использоваться для синтеза глюкозы.
Второй этап состоит в отщеплении аминогруппы от новообразованной аминокислоты (всегда глутамат) – происходит дезаминирование, которое осуществляется глутаматдегидрогеназой (реакцию см выше).
Учитывая тесную связь обоих этапов, непрямое окислительное дезаминирование назы-
вают трансдезаминирование.
Ниже подробно разбираются реакции трансаминирования и дезаминирования.
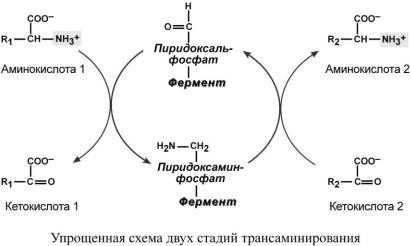
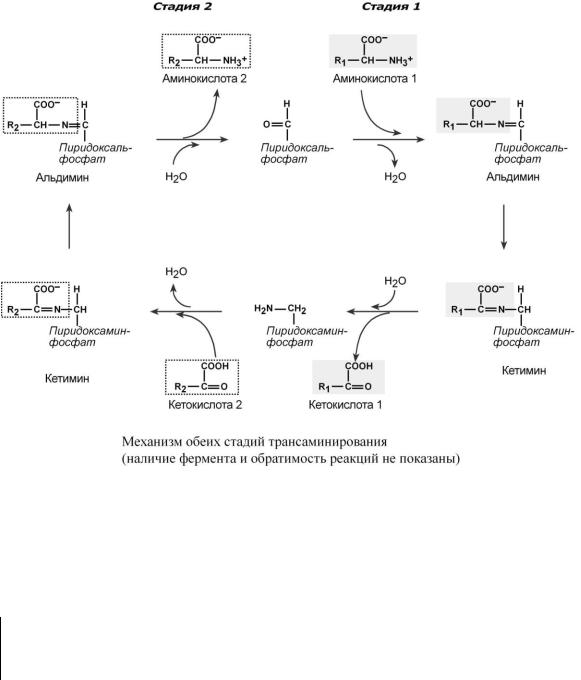
Механизм трансаминирования
Механизм реакции трансаминирования достаточно сложен. Катализируют реакцию ферменты аминотрансферазы, Они являются сложными ферментами, в качестве кофермента они имеют пиридоксальфосфат (активная форма витамина В6).
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
117 |
|
|
|
|
|
Весь перенос аминогруппы совершается в две стадии. К пиридоксальфосфату сначала присоединяется первая аминокислота, отдает аминогруппу, превращается в кетокислоту и отделяется. Аминогруппа при этом переходит на кофермент и образуется пиридоксаминфосфат. После этого на второй стадии присоединяется другая кетокислота, забирает аминогруппу, превращаясь в новую аминокислоту, пиридоксальфосфат регенерирует.
Роль и превращение пиридоксальфосфата сводится к образованию промежуточных соединений – шиффовых оснований (альдимин и кетимин). В первой реакции после отщепления воды образуется иминовая связь между остатком аминокислоты 1 и пиридоксальфосфатом. Полученное соединение называется альдимин. Перемещение двойной связи приводит к образованию кетимина, который гидролизуется водой по месту двойной связи. От фермента отщепляется готовый продукт – кетокислота 1.
Обмен аминокислот и белков |
118 |
|
|
После отщепления кетокислоты 1 к комплексу пиридоксамин-фермент присоединяется кетокислота 2, и процесс идет в обратном порядке: образуется кетимин, затем альдимин, после чего отделяется новая аминокислота 2.
Чаще всего аминокислоты взаимодействуют со следующими кетокислотами: пировиноградной (с образованием аланина), щавелевоуксусной (с образованием аспартата), -кето- глутаровой (с образованием глутамата). Однако аланин и аспартат в дальнейшем все равно передают свою аминогруппу на -кетоглутаровую кислоту.
В тканях насчитывают около 10 аминотрансфераз, которые обладают групповой специфичностью и вовлекают в реакции все аминокислоты, кроме пролина, лизина, треонина, которые не подвергаются трансаминированию.
Таким образом, в тканях осуществляется поток избыточных аминогрупп на один общий акцептор – -кетоглутаровую кислоту. В итоге образуется большое количество глутаминовой кислоты.
Дезаминирование
В организме коллектором всех аминокислотных аминогрупп (аминного азота) является глутаминовая кислота, и только она подвергается окислительному дезаминированию с образованием аммиака и -кетоглутаровой кислоты. Фермент глутаматдегидрогеназа имеется в митохондриях всех клеток организма (кроме мышечных) и катализирует реакцию дезаминирования глутамата.

biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
119 |
|
|
|
|
|
Так как НАДН используется в дыхательной цепи и -кетоглутарат вовлекается в реакции ЦТК, то реакция активируется при дефиците энергии при помощи АДФ и ингибируется избытком АТФ и НАДН.
Если реакция идет в митохондриях печени, аммиак используется для синтеза мочевины, которая в дальнейшем удаляется с мочой. В эпителии канальцев почек реакция необходима для удаления аммиака в процессе аммониегенеза.
ЭНЗИМОДИАГНОСТИКА С П ОМОЩЬЮ АМИНОТРАНСФЕР АЗ
Вмедицине нашло практическое применение определение активности двух аминотранс-
фераз – аланинаминотрансферазы (АЛТ) и аспартатаминтрансферазы (АСТ). Хотя актив-
ность обоих ферментов значительно возрастает при заболеваниях сердечной мышцы и печени, при поражении клеток миокарда наибольшая активность в сыворотке крови обнаруживается для АСТ, при гепатитах – для АЛТ.
Вклинической практике определение активности АЛТ и АСТ используется для дифференциальной диагностики болезней печени и миокарда, глубины поражения и контроля эф-
фективности их лечения.
Оба фермента обратимо взаимодействуют с -кетоглутаровой кислотой и переносят на нее аминогруппы от соответствующих аминокислот с образованием глутаминовой кислоты и кетокислот.