

Ответы на экзаменационные вопросы по биологии. Экзамен
.pdfназывают полуконсервативным. Также стоит сказать, что наряду с принципом комплиментарности, при удвоении ДНК соблюдается и еще один – принцип антипараллельности, это связано с разнонаправленностью цепей ДНК (матричная цепь ДНК 3’ – 5’, а синтезируемая ДНК 5’ – 3’).
Если в клетке, несмотря на осуществляемую репарацию, количество повреждений структуры ДНК остается высоким, в ней блокируются процессы репликации
ДНК. Такая клетка не делится, а значит, не передает возникших изменений потомству.
На геном любой неделящейся клетки постоянно оказывает влияние окружающая среда, при этом вполне вероятны повреждения в составе генома: изменение нуклеотида (например, дезаминирование), сшивки азотистых оснований друг с другом, разрывы цепей, отрыв пуриновых нуклеотидов и т.п. Такие изменения быстро определяются специальными ферментами, пораженный участок удаляется экзонуклеазами, заполняется ДНК-полимеразой β и сшивается ДНК-лигазой. В делящейся клетке мутации могут также возникать во время синтеза ДНК. Поэтому в клетках существует двойная система проверки точности репликации: одна непосредственно при ДНК-полимеразной реакции, другая – анализ уже синтезированной ДНК.
Вызываемая повреждениями ДНК остановка клеточного цикла в сочетании с невозможностью молекулярной репарации измененного наследственного материала может с участием белка, синтез которого контролируется геном р53, приводить к активации процесса самоликвидации (апоптоз) дефектной клетки с целью устранения ее из организма.
Несмотря на высокую точность процессов репликации и эффективно работающую систему коррекции во вновь синтезированных нитях ДНК всегда имеются нарушения. Эти дефекты именуют чаще всего как «генные мутации». Подсчёты показали, что они происходят с частотой1 ошибка на 1010 пар нуклеотидов. Редупликация, происходящая с ошибками носит название конвариантная редупликация.
9. Особенности организации генома эукариот. Регуляция экспрессии эукариотических генов.
Особенности организации генома эукариот:
У эукариот объем наследственного материала значительно больше, чем у прокариот.
Состав транскрибируемых последовательностей и их количество зависят от типа клетки и стадии онтогенеза. Значительная часть нуклеотидных последовательностей у эукариот не транскрибируется вообще — молчащая ДНК.
Большой объем наследственного материала эукариот объясняется существованием в нем помимо уникальных также умеренно и высоко повторяющихся последовательностей.
Так, около 10% генома мыши составляют друг за другом расположенные короткие нуклеотидные последовательности, повторенные до 106 раз. Эти высоко повторяющиеся последовательности ДНК располагаются в основном в гетерохроматине, окружающем центромерные участки. Они не транскрибируются.
Около 20% генома мыши образовано умеренными повторами, встречающимися с частотой 103—105 раз. Такие повторы распределены по всему геному и транскрибируются в РНК. К ним относятся гены, контролирующие синтез гистонов, тРНК, рРНК и
некоторые другие.
Остальные 70% генома мыши представлены уникальными нуклеотидными последовательностями.
Избыточность генома эукариот объясняется также экзон(кодирующие участки)-интронной(не кодирующие участки) организацией большинства эукариотических генов, при которой значительная часть транскрибированной РНК удаляется в ходе процессинга и не используется для кодирования аминокислотных последовательностей белков.
Функция молчащей ДНК не изучена до конца, однако высказываются положения о ее роли в структурной организации хроматина и регуляции экспрессии некоторых генов.
В эукариотических клетках внехромосомная ДНК представлена генетическим аппаратом органелл — митохондрий и пластид, а также ДНК и РНК вирусоподобных частиц. Наследственный материал органелл представлен несколькими копиями кольцевых молекул ДНК, не связанных с гистонами. В отличие от хромосомной ДНК, мтДНК характеризуется высокой «плотностью генов». В них нет интронов, а межгенные промежутки невелики.
Совокупность генов, расположенных в цитоплазматических молекулах ДНК, называют
плазмоном. Он определяет особый тип наследования признаков — цитоплазматическое наследование.
Регуляция экспрессии эукариотических генов:
Гены для синтеза ферментов к одной цепи биохимический реакций могут
располагаться в геноме рассеянно(в отличие прокариотического оперона) и не иметь единой системы регуляции. Поэтому отдельно синтезированные мРНК являются матрицами для отдельных пептидных цепей.
Комбинационная регуляция - активность каждого гена регулируется большим спектром генов-регуляторов.
Умногих эукариотических генов, кодирующих белки и транскрибируемых РНК-полимеразой II, в ДНК имеется несколько областей, которые узнаются разными
белками-регуляторами:
-область вблизи промотора(100 пар нуклеотидов): ТАТА-блок(25 пар нуклеотидов от точки начала транскрипции) - к нему присоединяется фактор транскрипции, этот комплекс затем узнается РНК-полимеразой II
-другая область располагается на большом расстоянии(несколько тысяч пар нуклеотидов) от промотора - энхансер, к которому присоединяется свой регуляторный белок
-в результате взаимодействия этих центров с белками происходит включение или выключение генов
Уэукариот существуют такие белки-регуляторы, которые могут контролировать транскрипцию многих генов(которые сами могут кодировать другие белки-регуляторы) -
плейотропный эффект действия.
Регуляция транскрипции подчинена влиянию гормонов организма, которые могут являться, в том числе, индукторами транскрипции.
Как особенность регуляции можно выделить существование хроматина - комплекса ДНК
сбелками.
Возможность осуществления регуляции экспрессии генов у эукариот продолжается на протяжении не только транскрипции, но на всех этапах реализации наследственной информации.
10. Множественные аллели как результат изменения нуклеотидной последовательности гена. Полиморфизм гена как вариант нормы и патологии. Примеры.
Множественный аллелизм — один из видов взаимодействия аллельных генов, при котором ген может быть представлен не двумя аллелями (как в случаях полного или неполного доминирования), а гораздо большим их числом; при этом члены одной серии аллелей могут находиться в различных доминантно-рецессивных отношениях друг с другом. Возникает полиаллелизм в следствие многократных генных мутаций одного локуса гена. У одной особи из огромного числа комбинаций аллельных генов может быть
только одна пара таких генов.
Примеры: У кроликов сплошная чёрная окраска обусловлена доминантным геном А, гомозиготные рецессивные животные (аа) — белые. Но существуют еще несколько состояний этого гена, имеющих собственный фенотип в гомозиготе — шиншилловой (achach) и гималайской (ahah) окраски. Шиншилла — сплошная серая масть, а гималайский
— белый, но кончики ушей, хвоста, ног и носа окрашены.
Полиморфными принято называть гены, которые представлены в популяции несколькими аллелями, что обусловливает разнообразие признаков внутри вида.
Существует много разных видов полиморфизма: некоторые из них - следствие генными мутациями многих (миллионов) пар нуклеотидов ДНК и не связаны с какими-либо патологическими процессами; другие изменения аналогичного масштаба оказываются
редкими вариантами, вызывающими тяжелые болезни организма; они могут быть связаны с изменениями в одном или нескольких основаниях ДНК, расположенными между генами или в интронах, не связанными с прямым функционированием генов и
обнаруживаемыми только при прямом анализе ДНК. Но также изменения могут находиться и в составе кодирующих генов, влияющих на процессы синтеза белка и соответственно на фенотип организма.
Примеры: муковисцидоз, мышечная дистрофия - моногенные болезни; предрасположенности к - злокачественным новообразованиям, сердечно-сосудистым заболеваниям, аллергии и так далее.
11. Ген, его свойства: дискретность, стабильность, лабильность, полиаллелизм, специфичность, плейотропия. Примеры.
Ген – элементарная функциональная единица наследственного материала, участок молекулы ДНК, кодирующий первичную последовательность аминокислот в структуре белка, т-/и(м)-/р-РНК.
Дискретность - наличие в гене субъединиц, минимальной из которых является пара комплементарных нуклеотидов, а минимальная функциональная единица - кодон.
Пример: строение генов(как такового примера нет :( ).
Стабильность - способность сохранять структуру и целостность гена и относительно
редкое явление мутации, наследование в ряду поколений в неизменном виде. Пример: перенос целостного гена через многие поколения.
Лабильность - способность гена изменяться, мутировать(в том числе, многократно).
Пример: появление различных мутаций в структуре гена.
Полиаллелизм(множественный аллелизм) - существование в популяции
множества(более двух) аллелей одного гена.
Пример: окраска шерсти у кроликов(черный, шиншилловый, гималайский, альбинос и т.д.), группы крови по системе АВ0.
Специфичность - каждый ген кодирует один собственный признак. Пример: ген кодирует один определенный фермент(пример "из головы", можете поискать в интернете).
Плейотропия - влияние одного гена на развитие разных признаков. Пример: ген платиновой окраски у лисиц в гомозиготном состоянии - летален; ген карликовости у мышей(рецессивная аллель) определяет ненормальное развитие гипофиза(что влияет на многие системы органов).
12. Фенотип как результат реализации генотипа в конкретных условиях среды. Среда первого, второго (а и б) и третьего порядка. Экспрессивность и пенетрантность гена.
Фенотип - совокупность индивидуальных характеристик организма, возникающие в процессе онтогенеза и представляющие собой сложный комплекс взаимодействия наследственной информации(генотипа) организма и окружающей среды. Фенотипическая изменчивость организма ограничена нормой реакции, которая, в зависимости от признака,
может иметь узкий либо широкий диапазон.
Среды различных порядков(по лекции Рамазановой):
-среда 1 порядка - совокупность генов организма
-среда 2 порядка - внутриорганизменные факторы:
-среда 2а порядка - внутренняя среда развивающегося организма(плода)
-среда 2б порядка - внутренняя среда материнского организма
-среда 3 порядка - внешние факторы(очень важные факторы, для человека - абиотические, социальные(интересны в качестве анамнеза пациента) факторы)
Экспрессивность гена - степень проявления признака в фенотипе организма. Можно сравнить с тяжестью болезни в клинической практики(для удобства понимания). В основе -
генетические факторы, факторы внешней среды. Количественно степень экспрессивности гена определяют, используя статистический показатель. Генетический признак может даже не проявляться в некоторых случаях. Если ген есть в генотипе, но он вовсе не проявляется – он пенетрирован.
Пенетрантность гена - количество особей(%) в популяции, у которых проявляется данный ген от количества особей, имеющих этот ген(в которых он мог бы проявиться). Пенетрантность свойственна проявлению многих генов. Важен принцип "всё или ничего" -
либо ген проявляется, либо нет.
13. Определение пола. Типы определения пола: прогамный, эпигамный, сингамный. Роль генотипа и среды в развитии признаков пола. Хромосомный механизм определения пола у разных организмов.
Пол у большинства животных и растений определяется генетически в момент оплодотворения. При исследовании кариотипов многих животных было установлено,что у женского организма каждая хромосома имеет парную (идентичную по размерам ,морфологии и содержанию генов),а у мужских организмов имеются две непарные хромосомы, которые резко отличаются по величине, морфологии и заключенной в них генетической информации. При дальнейшем исследовании было показано,что эти непарные хромосомы и определяют пол огранизма. Их назвали половыми хромосомами, в отличие от других – аутосом.Большую из непарных хромосом, одинаковую у мужского и женского организмов, назвали Х-хромосомой, а меньшую, имеющуюся только у мужского организма - Y-хромосомой.
Пол организма представляет собой важную фенотипическую характеристику, которая проявляется в совокупности свойств, обеспечивающих воспроизведение потомства и передачу ему наследственной информации. В зависимости от значимости этих свойств различают первичные и вторичные половые признаки.
Половые признаки — совокупность признаков, по которым мужской и женский пол отличают друг от друга. Половые признаки бывают первичными, или основными, и вторичными. Первые — это половые железы — яичники или семенники. Под первичными половыми признаками понимают морфофизиологические особенности организма, обеспечивающие образование половых клеток — гамет, сближение и соединение их в процессе оплодотворения. Это наружные и внутренние органы размножения. Вторичные половые признаки формируются в течение роста и полового созревания организма. У мужчин они проявляются в росте бороды, усов, появлении низкого тембра голоса и др., у женщин — в развитии молочных желез, в появлении определенных особенностей телосложения и других признаков. У человека и позвоночных животных вторичные половые признаки являются функцией деятельности половых желез. Интенсивность полового созревания людей зависит от социально-бытовых условий, наследственности и других причин.
Половой диморфизм — анатомические различия между самцами и самками одного и того же биологического вида, исключая различия в строении половых органов. Половой диморфизм может проявляться в различных физических признаках:
·Размер. У большинства млекопитающих и многих видов птиц самцы более крупные
итяжёлые, чем самки. У земноводных и членистоногих самки, как правило, крупнее самцов. У черепах и у змей самки крупнее самцов; у ящериц, напротив, как правило, самцы крупнее самок; исключение составляют гекконы, у которых большая величина также свойственна самкам.
·Волосяной покров. Борода у мужчин, грива у львов или бабуинов.
·Окраска. Цвет оперения у птиц, особенно у утиных.
·Кожа. Характерные наросты или дополнительные образования, такие как рога у оленевых, гребешок у петухов.
·Зубы. Бивни у самцов индийского слона, более крупные клыки у самцов моржей и кабанов.
Типы определения пола:
·Прогамный; прогамное определение пола происходит до оплодотворения, в процессе формирования яйцеклеток. Это имеет место, например, у коловраток. Они образуют яйцеклетки двух сортов: крупные, с двумя наборами хромосом (диплоидные) и большим объёмом цитоплазмы и мелкие, с одним набором хромосом — гаплоидные. Из гаплоидных неоплодотворённых яиц развиваются гаплоидные самцы, продуцирующие гаплоидные гаметы. Из крупных диплоидных яиц также развиваются самки, но в этом случае они появляются не в результате оплодотворения, а партеногенетически, то есть без оплодотворения. Таким образом, пол особи, развивающейся из диплоидного яйца, определяется ещё на стадии формирования яйцеклетки (на этом этапе закладывается её диплоидность), а пол особи, развивающейся из мелкого яйца, зависит от того, будет оно оплодотворено или нет;
·Эпигамный; под влиянием внешней среды – например, температуры; У некоторых рептилий пол определяется температурой, в которой развивалось яйцо в течение температурочувствительного периода. Это явление носит название температурозависимого определения пола; При эпигамном способе мужской или женский пол определяется факторами внешней среды, например, температурой в кладке яиц: у многих видов черепах при температуре ниже 27 °С развиваются только самцы, свыше 30 °С - только самки, в интервале 27-30 °С - самцы и самки. У большинства видов животных, размножающихся половым путем, в процессе исторического развития закрепились разные варианты «надежного» генотипического способа определения пола.
·Сингамный; сингамное определение пола происходит при оплодотворении, и пол определяется генетически.
·При эусингамном варианте (пчелы, муравьи), самцы первично гаплоидны, поскольку они развиваются из неоплодотворенных яйцеклеток, тогда как самки диплоидны. Напомним, что в процессе развития соматические клетки самцов таких животных становятся диплоидными. У плодовых мух наблюдается «балансовый» вариант генотипического способа, при котором пол определяется отношением числа хромосом Х к числу наборов аутосом (А). Если указанное отношение равно единице (ХХ/2А), развивается самка, при значении отношения 0,5 (ХY/2А) - самец, особи с кариотипом ХХ/3А (отношение меньше единицы, но больше 0,5) или ХY/3А (отношение меньше 0,5) - интерсексы. Исследования, выполненные также на плодовых мухах, заставляют думать, что генотипический механизм формирования комплекса фенотипических признаков женского или мужского пола у них более сложен. Так, у дрозофил на хромосоме 3 обнаружен локус tra или t с геном, изменяющим пол организма в сторону мужского вне зависимости от значений отношения числа хромосом Х и числа наборов аутосом. Особи как с генотипом ХY/2Аtt, так и с генотипом ХХ/2Аtt - фенотипически самцы, однако первые плодовиты
(образуют сперматозоиды), а вторые стерильны. Можно заключить, что для сперматогенеза хромосома Y необходима.
Хромосомный механизм определения пола у разных организмов
Предположительно с точки зрения интересов эволюционного процесса оптимален хромосомный вариант, получивший повсеместное распространение среди высокоорганизованных многоклеточных животных (амниоты - птицы, млекопитающие, включая человека), однако встречающийся у анамниа (земноводные) и среди членистоногих (некоторые виды клопов). Для этого варианта генотипического способа характерно, что один из полов (гомогаметный) образует одинаковые гаметы, тогда как второй (гетерогаметный) - разные. У млекопитающих гомогаметны женские особи, имеющие в кариотипе пару одинаковых половых хромосом (ХХ), а гетерогаметны - мужские особи, имеющие в кариотипе пару разных половых хромосом (ХY). У земноводных и птиц гомогаметны мужские особи (пара одинаковых половых хромосом - ZZ), тогда как гетерогаметны женские особи (пара разных половых хромосом
Источник KingMed.info 244
- ZW). Легко видеть, что при таком варианте генотипического способа определения один из полов (гомогаметный) по паре половых хромосом характеризуется как гомозиготный, тогда как второй (гетеро-гаметный) - как гетерозиготный.
Убольшинства организмов женский пол гомогаметный (хх),а мужской – гетерогаметный (ху). У птиц, бабочек, пресмыкающихся, гомогаметен (хх) мужской пол. У многих насекомых, например, у кузнечика, у некоторых моллюсков, у кенгуру Y-хромосомы вообще нет. У них клетки женских особей содержат две Х хромосомы, а клетки мужской - одну Х-хромосому (хо).
Умлекопитающих развитие половых признаков мужского пола обусловлено значительным количеством генов, большая часть которых расположена в аутосомах. Но начинают они работать только при наличии фактора, который расположен на маленькой Y-хромосоме. Развитие зародышей крокодила у мужских и женских особей определяется температурой, при которой происходит инкубация яиц.32-33 градуса - самцы, выше или ниже - самки.
Развитие пола морского беспозвоночного бонелии определяется влиянием внешних факторов. Этот фактор – наличие взрослой самки. Если ее нет, то личинки развиваются только как самки. Если есть - только как самцы.
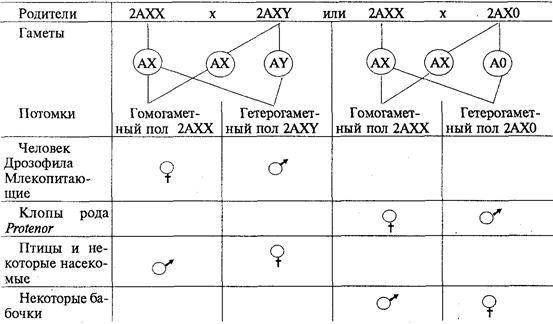
У разных видов организмов хромосомный механизм определения пола реализуется по-разному. У человека и других млекопитающих, а также у дрозофилы гомогаметным является женский пол (XX), а гетерогаметным — мужской (XY). У некоторых насекомых (клопы рода Protenor) гетерогаметный мужской пол имеет лишь одну Х-хромосому (ХО). У птиц и некоторых насекомых женский пол является гетерогаметным (XY), а мужской — гомогаметным (XX). У некоторых бабочек гетерогаметный женский пол имеет одну Х-хромосому (ХО). В большинстве выше описанных случаев пол вновь образующегося организма определяется сочетанием половых хромосом, возникающим в зиготе при оплодотворении (табл. 6.2).
Таблица 6.2. Хромосомный механизм определения полов
Таким образом, хромосомный механизм определения половой принадлежности организмов обеспечивает равновероятность встречаемости представителей обоих полов. Это имеет большой биологический смысл, так как обусловливает максимальную вероятность встречи самки и самца, потомки получают более разнообразную наследственную информацию, поддерживается оптимальная численность особей в популяции.
Исследования, проведенные на дрозофиле, обнаружили незначительную роль Y-хромосомы в определении пола. Особи с кариотипом ХО внешне являются типичными самцами, но стерильными, а особи с кариотипом XXY — плодовитые самки. Y-хромосома у дрозофилы по размеру близка к Х-хромосоме, однако она генетически инертна, так как состоит в основном из гетерохроматина. В связи с этим первичное определение пола у дрозофилы обусловлено не столько сочетанием в зиготе Х- и Y-хромосом, сколько соотношением числа Х-хромосом и наборов аутосом (половой индекс). У нормальных самок половой индекс равен 1 (2Х:2А), у нормальных самцов — 0,5 (XY:2A). Увеличение
полового индекса более 1 при увеличении дозы Х-хромосомы (ЗХ:2А) приводит к развитию сверхсамок, а величина этого показателя ниже 0,5 (XY:3A) обеспечивает появление самцов. При значениях полового индекса более 0,5, но менее 1 (2Х:ЗА) развиваются интерсексы.
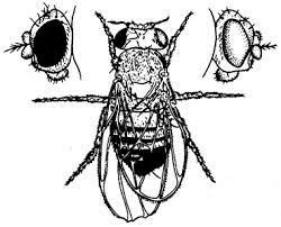
О значении дозы Х-хромосом в определении пола у дрозофилы свидетельствует развитие латерального гинандроморфизма у мух при утрате Х-хромосомы одной из клеток на стадии первого деления зиготы. В результате половина тела, развивающаяся из клетки с нормальным кариотипом (2АХХ), несет признаки самки, а другая половина, клетки которой лишены одной Х-хромосомы (2АХО), имеет признаки самца (рис. 6.2).
Рис. 6.2. Латеральный гинадроморфизм у Drosophila melanogaster, гетерозиготной по гену окраски глаз, находящемуся в Х-хромосоме
Левая половина мухи, клетки которой несут 2Х, имеет признаки самки и красный глаз; правая половина мухи, клетки которой несут одну Х-хромосому, имеет признаки самца и белый глаз
Обнаружение зависимости половой принадлежности развивающегося организма от дозы Х-хромосом у дрозофилы и некоторых других насекомых привело американского генетика С. Бриджеса (1922) к формулировке гипотезы генного баланса, в соответствии с которой организм изначально бисексуален, т.е. несет в себе задатки и мужского, и женского полов. Развитие признаков одного из них в ходе онтогенеза определяется балансом женских и мужских генов — детерминаторов пола. У дрозофилы эти гены сосредоточены не только в половой Х-хромосоме, но и в аутосомах. Поэтому пол организма у них зависит от соотношения этих хромосом. У плодовой мухи Y-хромосома, будучи генетически инертной,
вопределении признаков пола не участвует.
Учеловека Y-хромосома играет важную роль в детерминации пола. Она содержит определенное количество генов, часть из которых гомологична генам Х-хромосомы, а часть не имеет в ней гомологов и наследуется только по мужской линии. Некоторые из этих генов непосредственно связаны с детерминацией мужского пола. Поэтому у человека присутствие Y-хромосомы в кариотипе независимо от количества Х-хромосом (2AXXY, 2AXXXY) обеспечивает развитие мужского пола. Особи с кариотипом 2АХО являются женщинами,
несмотря на уменьшение дозы Х-хромосомы. Однако значение баланса генов в определении половой принадлежности организма подтверждается тем, что особи с кариотипами 2АХО, 2АХХХ, 2AXXY, 2AXXXY и т.д. отличаются наличием пороков развития и часто стерильны.
Как было отмечено, у большинства видов животных пол организма определяется сочетанием или количеством половых хромосом в диплоидном наборе аутосом. Однако у некоторых насекомых (пчелы, муравьи, осы) представители разных полов отличаются числом наборов хромосом. Самки у них развиваются из оплодотворенных яиц и поэтому диплоидны, самцы — из неоплодотворенных яиц и первично гаплоидны. В такой ситуации соотношение по полу зависит от того, происходит оплодотворение или нет. Половой состав популяции может быть различным в разные периоды ее жизни.
Зависимость половой принадлежности организмов от сочетания или количества половых хромосом в его кариотипе свидетельствует о том, что гены, определяющие пол, располагаются главным образом в этих хромосомах. Однако известен целый ряд примеров, указывающих на то, что в развитии признаков пола принимают участие также гены, локализованные в аутосомах. В частности, в геноме дрозофилы присутствует ген, называемый трансформатором. Рецессивный аллель этого гена в гомозиготном состоянии tt превращает женские зиготы 2Х в фенотипических самцов, которые оказываются стерильными. Самки1 XXt+t+ или XXt+t плодовиты, особи с кариотипом XYtt — нормальные самцы, организмы с кариотипом XXtt — превращенные стерильные самцы. Другим примером влияния аутосомных генов на развитие признаков пола может служить существование у кукурузы генов, мутантные аллели которых превращают ее из однодомного растения в двудомное, препятствуя развитию органов либо мужского, либо женского пола. Рецессивный аллель гена sk(silkless) в гомозиготном состоянии вызывает недоразвитие семяпочек и превращает растение в мужское. Мутантный аллель другого гена ts (tassel seed) в гомозиготном состоянии вызывает развитие семяпочек и семян в метелке, а пыльники не развиваются.
У человека развитие организма по мужскому типу обеспечивается не только геном, расположенным в Y-хромосоме и определяющим способность к синтезу мужского полового гормона — тестостерона, но и Х-сцепленным геном, контролирующим синтез белка-рецептора этого гормона. Мутация упомянутого Х-сцепленного гена приводит к развитию синдрома тестикулярной феминизации (см. разд. 3.6.5.2).
Приведенные примеры доказывают, что пол организмов как признак развивается на основе полученной от родителей наследственной информации и находится под контролем группы взаимодействующих генов, расположенных как в половых хромосомах, так и в аутосомах.