

Ответы на экзаменационные вопросы по биологии. Экзамен
.pdfактивируемым видимым светом, циклобутановых димеров, возникающих в ДНК под действием ультрафиолетового излучения.
Пострепликативная репарация - пострепликативная репарация «включается» тогда, когда эксцизионная репарация «не справляется» с устранением всех повреждений, возникших в ДНК до её репликации. При репликации во второй спирали ДНК возникают бреши - однонитевые пробелы, соответствующие изменённым нуклеотидам первой спирали. Бреши заполняются участками цепи с нормальной последовательностью нуклеотидов уже в ходе пострешгикативной репарации при участии ДНК-полимеразы.
Позднее была обнаружена и темновая репарация, т. е. свойство клеток ликвидировать повреждения ДНК без участия видимого света. Темновая репарация осуществляется комплексом из пяти ферментов:
·узнающего химические изменения на участке цепи ДНК;
·осуществляющего вырезание поврежденного участка;
·удаляющего этот участок;
·синтезирующего новый участок по принципу комплементарности взамен удаленного фрагмента;
·соединяющего концы старой цепи и восстановленного участка.
При световой репарации исправляются повреждения, возникшие только под воздействием ультрафиолетовых лучей, при темновой — повреждения, появившиеся под влиянием жесткой радиации, химических веществ и других факторов. Темновая репарация обнаружена как у прокариот, так и в клетках эукариот. У последних она изучается в культурах тканей. Вопрос о том, почему одни повреждения репарируются, а другие нет, остается открытым. Если репарация не наступает, то клетка либо гибнет, либо наступает мутация.
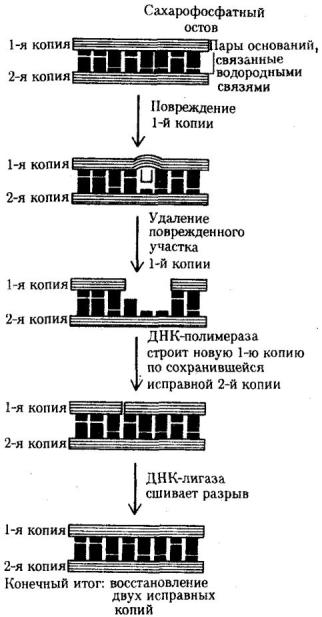
Механизм репарации основан на наличии в молекуле ДНК двух комплементарных цепей. Искажение последовательности нуклеотидов в одной из них обнаруживается специфическими ферментами. Затем соответствующий участок удаляется и замещается новым, синтезированным на второй комплементарной цепи ДНК. Такую репарацию называют эксцизионной, т.е. с «вырезанием» (рис. 3.15). Она осуществляется до очередного цикла репликации, поэтому ее называют также дорепликативной.
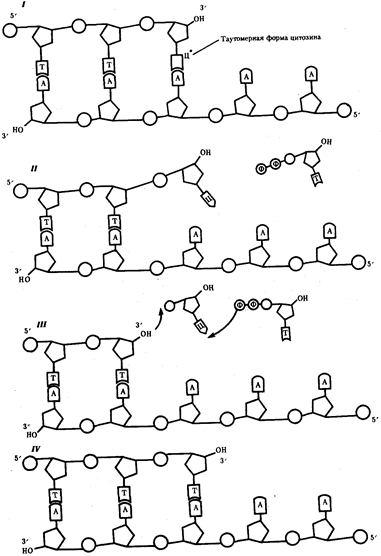
Рис. 3.14. Схема процесса коррекции при синтезе ДНК:
I—включение в цепь ДНК нуклеотида с измененной (таутомерной) формой цитоэина, который «незаконно» спаривается с аденином; II — быстрый переход цитозина в обычную форму нарушает его спаривание с аденином; неспаренный 3'—ОН-конец синтезируемой цепи препятствует дальнейшему ее удлинению под действием ДНК-полимеразы; III — ДНК-полимераза удаляет незаконный нуклеотид, в результате чего вновь появляется спаренный с матрицей 3 '—ОН-конец; IV — ДНК-полимераза продолжает наращивание цепи на 3'—ОН-конце
Восстановление исходной структуры ДНК требует участия ряда ферментов. Важным моментом в запуске механизма репарации является обнаружение ошибки в структуре ДНК. Нередко такие ошибки возникают во вновь синтезированной цепи в процессе репликации. Ферменты репарации должны обнаружить именно эту цепь.
У многих видов живых организмов вновь синтезированная цепь ДНК отличается от материнской степенью метилирования ее азотистых оснований, которое отстает от синтеза. Репарации при этом подвергается неметилированная цепь. Объектом узнавания ферментами репарации могут также служить разрывы в цепи ДНК. У высших организмов, где синтез
ДНК происходит не непрерывно, а отдельными репликонами, вновь синтезируемая цепь ДНК имеет разрывы, что делает возможным ее узнавание.
Восстановление структуры ДНК при утрате пуриновых оснований одной из ее цепей предполагает обнаружение дефекта с помощью фермента эндонуклеазы, которая разрывает фосфоэфирную связь в месте повреждения цепи. Затем измененный участок с несколькими примыкающими к нему нуклеотидами удаляется ферментом экзонуклеазой, а на его месте в соответствии с порядком оснований комплементарной цепи образуется правильная нуклеотидная последовательность (рис. 3.15).
Рис. 3.15. Схема эксцизионной, дорепликативной репарации ДНК
При изменении одного из оснований в цепи ДНК в восстановлении исходной структуры принимают участие ферменты ДНК-гликозилазы числом около 20. Они специфически узнают повреждения, обусловленные дезаминированием, алкилированием и другими структурными преобразованиями оснований. Такие модифицированные основания
удаляются. Возникают участки, лишенные оснований, которые репарируются, как при утрате пуринов. Если восстановление нормальной структуры не осуществляется, например в случае дезаминирования азотистых оснований, происходит замена одних пар комплементарных оснований другими —пара Ц—Г может заменяться парой Т—А и т.п. (см. разд. 3.4.2.3).
Образование в полинуклеотидных цепях под действием УФ-лучей тиминовых димеров (Т—Т) требует участия ферментов, узнающих не отдельные измененные основания, а более протяженные повреждения структуры ДНК. Репаративный процесс в этом случае также связан с удалением участка, несущего димер, и восстановлением нормальной последовательности нуклеотидов путем синтеза на комплементарной цепи ДНК.
В том случае, когда система эксцизионной репарации не исправляет изменения, возникшего в одной цепи ДНК, в ходе репликации происходит фиксация этого изменения и оно становится достоянием обеих цепей ДНК. Это приводит к замене одной пары комплементарных нуклеотидов на другую либо к появлению разрывов (брешей) во вновь синтезированной цепи против измененных участков. Восстановление нормальной структуры ДНК при этом может произойти и после репликации.
Пострепликативная репарация осуществляется путем рекомбинации (обмена фрагментами) между двумя вновь образованными двойными спиралями ДНК. Примером такой пострепликативной репарации может служить восстановление нормальной структуры ДНК при возникновении тиминовых димеров (Т—Т), когда они не устраняются самопроизвольно под действием видимого света (световая репарация) или в ходе дорепликативной эксцизионной репарации.
Ковалентные связи, возникающие между рядом стоящими остатками тимина, делают их не способными к связыванию с комплементарными нуклеотидами. В результате во вновь синтезируемой цепи ДНК появляются разрывы (бреши), узнаваемые ферментами репарации. Восстановление целостности новой полинуклеотидной цепи одной из дочерних ДНК осуществляется благодаря рекомбинации с соответствующей ей нормальной материнской цепью другой дочерней ДНК. Образовавшийся в материнской цепи пробел заполняется затем путем синтеза на комплементарной ей полинуклеотидной цепи (рис. 3.16). Проявлением такой пострепликативной репарации, осуществляемой путем рекомбинации между цепями двух дочерних молекул ДНК, можно считать нередко наблюдаемый обмен материалом между сестринскими хроматидами (рис. 3.17).
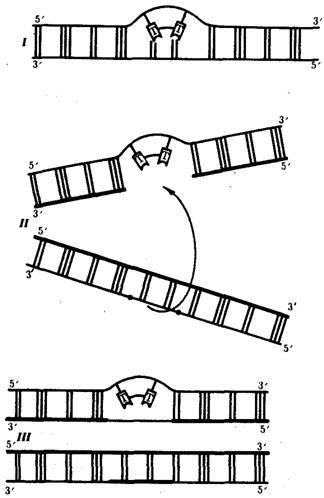
Рис. 3.16. Схема пострепликативной репарации ДНК:
I — возникновение тиминового димера в одной из цепей ДНК;
II — образование «бреши» во вновь синтезируемой цепи против измененного участка материнской молекулы после репликации (стрелкой показано последующее заполнение «бреши» участком из соответствующей цепи второй дочерней молекулы ДНК);
III— восстановление целостности дочерней цепи верхней молекулы за счет рекомбинации
ив нижней молекуле за счет синтеза на комплементарной цепи
В ходе дорепликативной и пострепликативной репарации восстанавливается большая часть повреждений структуры ДНК. Однако, если в наследственном материале клетки возникает слишком много повреждений и часть из них не ликвидируется, включается система индуцируемых (побуждаемых) ферментов репарации (SOS-система). Эти ферменты заполняют бреши, восстанавливая целостность синтезируемых полинуклеотидных цепей без точного соблюдения принципа комплементарности. Вот почему иногда сами процессы репарации могут служить источником стойких изменений в структуре ДНК (мутаций). Названная реакция также относится к SOS-системе.
Если в клетке, несмотря на осуществляемую репарацию, количество повреждений структуры ДНК остается высоким, в ней блокируются процессы репликации ДНК. Такая клетка не делится, а значит, не передает возникших изменений потомству.
Вызываемая повреждениями ДНК остановка клеточного цикла в сочетании с невозможностью молекулярной репарации измененного наследственного материала может с участием белка, синтез которого контролируется геном р53, приводить к активации процесса самоликвидации (апотпоз) дефектной клетки с целью устранения ее из организма.
Таким образом, обширный набор различных ферментов репарации осуществляет непрерывный «осмотр» ДНК, удаляя из нее поврежденные участки и способствуя поддержанию стабильности наследственного материала. Совместное действие ферментов репликации (ДНК-полимераза и редактирующая эндонуклеаза) и ферментов репарации обеспечивает достаточно низкую частоту ошибок в молекулах ДНК, которая поддерживается на уровне 1 · 10-9 пар измененных нуклеотидов на геном. При размере генома человека 3 · 109 нуклеотидных пар это означает появление около 3 ошибок на реплицирующийся геном. Вместе с тем даже этот уровень достаточен для образования за время существования жизни на Земле значительного генетического разнообразия в виде генных мутаций.
Связь мутаций с репарацией ДНК.
Спонтанные повреждения ДНК встречаются довольно часто, такие события имеют место в каждой клетке. Для устранения последствий подобных повреждений имеется специальные репарационные механизмы (например, ошибочный участок ДНК вырезается и на этом месте восстанавливается исходный). Мутации возникают лишь тогда, когда репарационный механизм по каким-то причинам не работает или не справляется с устранением повреждений. Мутации, возникающие в генах, кодирующих белки, ответственные за репарацию, могут приводить к многократному повышению (мутаторный эффект) или понижению (антимутаторный эффект) частоты мутирования других генов. Так, мутации генов многих ферментов системы эксцизионной репарации приводят к резкому повышению частоты соматических мутаций у человека, а это, в свою очередь, приводит к развитию пигментной ксеродермы и злокачественных опухолей покровов.
Дополнение:
Мутация и репарация
Генетическая информация кодируется последовательностью оснований ДНК и поэтому изменения в структуре или последовательности азотистых оснований приводят к мутациям. Многие мутагены вызывают нарушения регуляции роста и деления клеток и поэтому являются канцерогенными. Изменение в структуре генов (мутация) — важный фактор биологической эволюции. В то же время слишком высокая скорость мутаций ставит под вопрос существование индивидуальных организмов или целых видов. Поэтому
клетки обладают механизмами восстановления (репарации), которые корректируют большинство изменений ДНК, вызываемых мутациями.
Механизмы репарации
Важным механизмом удаления повреждений в ДНК является эксцизионная репарация (1). Специфическая нуклеаза удаляет небольшой сегмент ДНК, включающий поврежденный участок. Удаленный участок восстанавливается ДНК-полимеразой, использующей в качестве матрицы комплементарную цепь. Наконец, оставшийся одноцепочечный разрыв закрывается ДНК-лигазой. Тиминовые димеры могут быть удалены фотореактивацией (2). Специфическая фотолиаза связывается с дефектным участком ДНК и после облучения расщепляет димер с образованием отдельных нуклеиновых оснований. Третий механизм — это репарация в результате рекомбинации (3, показано в упрощенном виде). В этом процессе участок, содержащий повреждение, пропускается во время репликации. Образующаяся брешь закрывается путем сдвига соответствующего сегмента из правильно реплицированной второй цепи. Новая брешь ликвидируется с участием полимераз и ДНК-лигаз. В завершение первоначальный дефект (1) устраняется путем вырезания.
17. Изменчивость, её формы. Модификационная изменчивость; адаптивный характер модификаций. Норма реакции генетически детерминированных признаков. Морфозы и фенокопии. Примеры.
Изменчивость - это возникновение индивидуальных различий. На основе изменчивости организмов появляется генотипическое разнообразие форм, к-ые в результате действия ЕО преобразуются в новые подвиды и виды. Различают модификационную (фенотипическая), мутационная (генотипическая): мутационная, комбинативная, соотносительная (коррелятивная).
Наследственная (генотипическая, неопределённая): затрагивает генотип, имеет индивидуальный характер, носит внезапный скачкообразный характер, изменения неадекватны условия среды.
Ненаследственная (фенотипическая, определённая): затрагивает только фенотип, имеет групповой характер, форм-ся в онтоногенезе, зависит от условий среды (изм-ия адекватны условия среды), изм-ия в пределах нормы р-ии, изм-ия носят постепенный характер.
Модификационная изменчивость не вызывает изменений генотипа, она связана с реакцией данного, одного и того же генотипа на изменения внешней среды: в оптимальных условиях выявляется максимум возможностей, присущих данному генотипу (например, в условиях улучшенного содержания и ухода коров повышается надои молока, нагул мяса; в этом случае все особи с одинаковым генотипом отвечают на внешние условия одинаково. Однако другой признак -жирность молокаслабо подвержен изменениям, а масть животного ещё более устойчивый признак). Норма р-ии- степень варьирования признака у организма, т.е. пределы модификационной изменчивости. Широкая Н.Р.- удои молока, размеры листьев; узкая Н.Р.-жирность молока, яйценоскость у кур; однозначная Н.Р.- (большинство качественных признаков - цвет волос, глаз). Фенотип форм-ся в результате взаим-ия генотипа и факторов среды. Фенотипические признаки не передаются от родителей

потомкам, наслед-ся лишь норма р-ии, т.е. характер реагирования на изм-ия окружающих условий. Св-ва модификаций: 1) ненаследуемость; 2) групповой характер изменений; 3) соотнесение изменений действию определённого фактора среды; 4) обусловленность пределов изменчивости генотипом. Приспособительный (адаптивный) характер имеют все наиболее распространенные модификации. Так повышение числа эритроцитов и содержание Hb в крови животных и человека в горах представляют приспособление для лучшего использования кислорода. Загар кожи - приспособление воздействия чрезмерной инсоляции. Установлено, что адаптивными бывают только те модификации, которые вызываются обычными изменениями природных условий. Не имеют приспособительного значения модификации, вызываемые различными химическими и физическими факторами.
Морфоз — ненаследственное изменение фенотипа организма в онтогенезе под влиянием экстремальных факторов среды. Морфозы возникают в критические периоды онтогенеза и не носят приспособительного характера. Фенотипически морфозы сходны с мутациями и в таких случаях они называются фенокопиями. Механизмом фенокопий является нарушение реализации наследственной информации. Они возникают вследствие подавления функции определенных генов. По своему проявлению они напоминают функцию известных генов, но не наследуются.
Фенокопии— изменение признака под влиянием внешних факторов в процессе его развития, зависящего от определенного генотипа, ведущего к копированию признаков, характерных для другого генотипа или его отдельных элементов. Такие изменения вызваны факторами внешней среды, однако их фенотип напоминает (копирует) проявление наследственных синдромов.
Итак, фенокопия прикидывается мутацией, вызванной генотипом, а морфоз - просто необратимо искореженный фенотип» (с)
Возникшие фенотипические модификации не наследуются (генотип не изменяется). Примером проявления фенокопий могут служить заболевания, приводящие к кретинизму, которые могут обусловливаться наследственными и передовыми (в частности, отсутствием йода в рационе ребенка, независимо от его генотипа) факторами.
18. Комбинативная изменчивость, причины возникновения. Значение комбинативной изменчивости в обеспечении генетического разнообразия людей.
Комбинативная изменчивостьизменчивость, приводящая к новому сочетанию признаков вследствие комбинации генов; приводит к появлению нестабильных изменений. Источники: конъюгация и кроссинговер в профазе мейоза 1; независимое расхождение хромосом в анафазе мейоза; случайное слияние гамет.
Комбинативная наследственная изменчивость возникает в результате обмена гомологичными участками гомологичных хромосом в процессе мейоза, а также как следствие независимого расхождения хромосом при мейозе и случайного их сочетания при скрещивании. Изменчивость может быть обусловлена не только мутациями, но и сочетаниями отдельных генов и хромосом, новая комбинация к-ых при размножении приводит к изменению определённых признаков и св-в организма.
Причина: возникает стихийно в рамках популяции при скрещивании, когда у потомков появляются новые комбинации генов.Значение: распространение в популяции новых наследственных изменений, к-ые служат материалом для отбора.
Примеры: появление розовых цветков при скр-ии белоцветковой и красноцветковой примул. При скр-ии белого и серого кроликов может появиться чёрное потомство.
19. Мутационная изменчивость. Классификация мутаций (общие принципы). Соматические и генеративные мутации.
Мутационная изменчивость — изменчивость, вызванная действием на организм мутагенов, вследствие чего возникают мутации.
Мутациями наз-ся скачкообразные и устойчивые изм-ия единиц наследственности - генов, влекущие за собой изм-ия наследственных признаков. Термин мутация был впервые введён де Фризом. Мутации обязательно вызывают изм-ия генотипа, к-ые наслед-ся потомством и не связаны со скрещиванием и рекомбинацией генов.
Классификация мутаций. По характеру проявления:
1)доминантные и рецессивные;
2)полулетальные (мутации, резко снижающие жизнеспособность, частично или полностью останавливающие развитие) и летальные (несовместимые с жизнью).
По месту их возникновения:
1)генеративные (мутация, возникшая в половых клетках, не влияет на признаки данного орг-ма, а проявляется только в следующем поколении);
2)соматические (если изм-ся гены в соматических клетках, такие мутации проявляются у данного орг-ма не передаются потомству при половом размножении. Но при бесполом размножении, если орг-м разв-ся из кл или группы кл, имеющих мутировавший ген, мутации могут передаваться потомству).
По адаптивному значению (условнно):
1)Полезные мутации: мутации, которые приводят к повышенной устойчивости организма (устойчивость тараканов к ядохимикатам).
2)Вредные мутации: глухота, дальтонизм.
3)Нейтральные мутации: мутации никак не отражаются на жизнеспособности организма (цвет глаз, группа крови).
По уровню их возникновения:
1) хромосомные (связаны с изм-ем стр-ры хром-м);
2)генные (затрагивают стр-ру самого гена и влекут за собой изм-ие св-в орг-ма (гемофилия, дальтонизм, альбинизм));
3)геномные мутации приводят к изменению числа хромосом. Различают анеуплоидию и полиплоидию. Анеуплоидия связана с изменением числа хромосом на несколько хромосом
(1, 2, 3):
А) моносомия общая формула 2n-1 (45, Х0), болезнь – синдром Шерешевского-Тернера.
Б) трисомия общая формула 2n+1 (47, ХХХ или 47, ХХУ) болезнь – синдром Клайнфельтра.
В) полисомия.
По локализации:
1)ядерные (в ядре);
2)цитоплазматические (в митохондриях и хлоропластах).
По направлению:
1)прямые (переход гена от дикого типа к новому состоянию (мутантному));
2)обратные (переход гена от мутантного состояния к дикому типу).
По способу возникновения:
1) индуцированные (возникли при воздействии мутагенов)
Мутагеныфакторы, вызывающие мутации:
Физические (радиация, электро-магнитное излучение, температура);
Химические (гербициды, ароматические углеводороды);
Биологические (бактерии, вирусы)