

Ответы на экзаменационные вопросы по биологии. Экзамен
.pdf

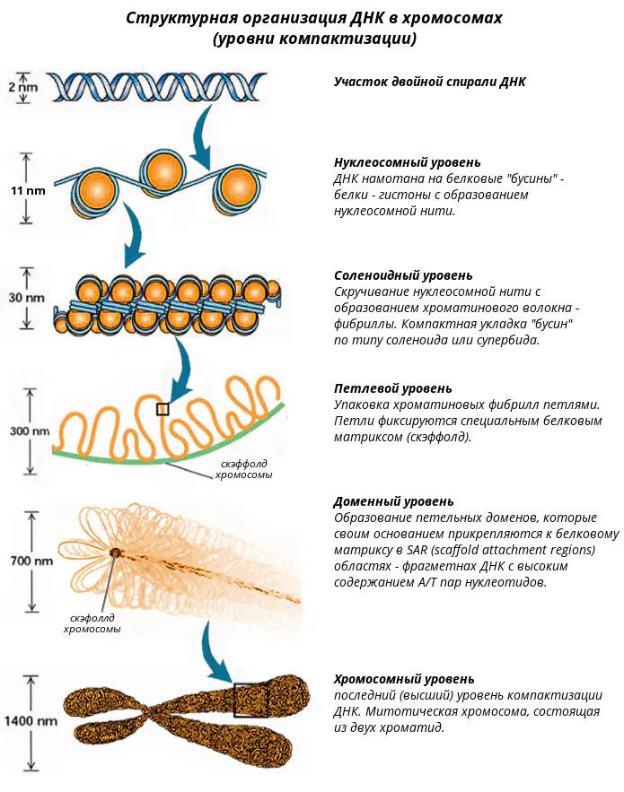
6. Функции нуклеиновых кислот в процессе реализации наследственной информации. Кодирование наследственной информации в клетке. Генетический код и его свойства.
Исследования, направленные на выяснение химической природы наследственного материала, неопровержимо доказали, что материальным субстратом наследственности и изменчивости являются нуклеиновые кислоты, которые были обнаружены Ф. Мишером (1868) в ядрах клеток гноя.Среди нуклеиновых кислот различают два вида
соединений:дезоксирибонуклеиновую (ДНК) и рибонуклеиновую (РНК) кислоты. Изучение состава основных носителей наследственного материала — хромосом обнаружило, что их наиболее химически устойчивым компонентом является ДНК, которая представляет собой субстрат наследственности и изменчивости.Наследственная информация, записанная с помощью генетического кода, хранится в молекулах ДНК и размножается для того, чтобы обеспечить вновь образуемые клетки необходимыми «инструкциями» для их нормального развития и функционирования.
Функции ДНК
1.ДНК является носителем генетической информации. Функция обеспечивается фактом существования генетического кода.
2.Воспроизведение и передача генетической информации в поколениях клеток и организмов. Функция обеспечивается процессом репликации.
3.Реализация генетической информации в виде белков, а также любых других соединений, образующихся с помощью белков-ферментов. Функция обеспечивается процессами транскрипции и трансляции.
Вместе с тем непосредственного участия в жизнеобеспечении клеток ДНК не принимает. Роль посредника, функцией которого является перевод наследственной информации, сохраняемой в ДНК, в рабочую форму, играют рибонуклеиновые кислоты — РНК.
Вотличие от молекул ДНК рибонуклеиновые кислоты представлены одной полинуклеотидной цепью, которая состоит из четырех разновидностей нуклеотидов, содержащих сахар, рибозу, фосфат и одно из четырех азотистых оснований - аденин, гуанин, урацил или цитозин. РНК синтезируется на молекулах ДНК при помощи ферментов РНК-полимераз с соблюдением принципа комплементарности и антипараллельности, причем аденину ДНК в РНК комплементарен урацил. Все многообразие РНК, действующих в клетке, можно разделить на три основных вида: мРНК, тРНК, рРНК.
Функция информационной РНК, синтезируемая на ДНК (точнее синтезируется ее предшественник), после своего созревания переносит информацию о структуре белка с ДНК на рибосомы. Это функция иРНК. В большинстве случаев каждый белок кодируется одной отдельной молекулой иРНК. Таким образом, разновидностей иРНК существует множество.
Основная функция рибосомальной РНК – структурная. Рибосомальная РНК вместе с рядом белков формирует субъединицы рибосом. Рибосомальная РНК не только формирует субъединицы рибосом, но и часть ее молекул непосредственно участвуют в синтезе белка на рибосомах.
Функция транспортной РНК — перенос аминокислот к месту синтеза полипептидной цепочки, т. е. на рибосомы. Данная функция реализуется благодаря тому, что у тРНК есть антикодон, комплиментарный определенному участку иРНК. В зависимости от своего антикодона каждая тРНК способна связываться только со «своей» аминокислотой.
Кроме перечисленных основных функций РНК, соответствующих основным типам РНК, существуют и другие типы РНК, выполняющие «дополнительные» функции.
Так малая ядерная РНК принимает участие в процессе созревания иРНК, предшественником которой является гетерогенная ядерная РНК (именно она синтезируется на ДНК, после чего превращается в иРНК).
РНК-праймер представляет собой маленькую молекулу РНК, которая принимает участие в реализации процесса репликации (удвоения) ДНК.
Некоторые молекулы РНК в клетках выполняют каталитическую функцию.
Генетический код
Генетический код - это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности нуклеотидов в ДНК. Поскольку ДНК непосредственного участия в синтезе белка не принимает, то код записывается на языке РНК. В РНК вместо тимина входит урацил.
Свойства генетического кода
1.Триплетность. Каждая аминокислота кодируется последовательностью из 3-х нуклеотидов.
2.Вырожденность. Все аминокислоты, за исключением метионина и триптофана, кодируются более чем одним триплетом. Всего 61 триплет кодирует 20 аминокислот.
3.Однозначность. Каждый триплет кодирует лишь одну аминокислоту или является терминатором трансляции.
4.Наличие межгенных знаков препинания. В конце каждого гена, кодирующего полипептид, находится, по меньшей мере, один из 3-х терминирующих кодонов, или стоп-сигналов: UAA, UAG, UGA. Они терминируют трансляцию. Условно к знакампрепинания относится инициирующий кодон AUG - первый после лидерной последовательности, кодирует формилметионин (у прокариот).
5.Компактность, или отсутствие внутригенных знаков препинания. Внутри гена каждый нуклеотид входит в состав значащего кодона.
6.Колинеарность – последовательность кодонов соответствует последовательности аминокислот в кодируемом белке.
7.Неперекрываемость – триплеты не накладываются друг на друга, располагаясь рядом.
8.Универсальность. Генетический код един для всех живущих на Земле существ. Объяснение:
Первично все многообразие жизни обусловливается разнообразием белковых молекул,
выполняющих в клетках различные биологические функции. Структура белков определяется набором и порядком расположения аминокислот в их пептидных цепях. Именно эта последовательность аминокислот в пептидах зашифрована в молекулах ДНК с помощью биологического (генетического) кода.
В многообразии белков, существующих в природе, было обнаружено около 20 различных аминокислот. Для шифровки такого их числа достаточное количество сочетаний нуклеотидов может обеспечить лишь триплетный код, в котором каждая аминокислота шифруется тремя стоящими рядом нуклеотидами. В этом случае из четырех нуклеотидов образуется 4 в третьей степени= 64 триплета. Код, состоящий из двух нуклеотидов, дал бы возможность зашифровать только 4 во второй степени= 16 различных аминокислот.
Из 64 возможных триплетов ДНК 61 кодирует различные аминокислоты;оставшиеся 3 получили название бессмысленных, или «нонсенс-триплетов». Они не шифруют аминокислот и выполняют функцию знаков препинания при считывании наследственной информации. К ним относятся АТТ, АЦТ, АТЦ.
Обращает на себя внимание явная избыточность кода, проявляющаяся в том, что многие аминокислоты шифруются несколькими триплетами. Это свойство триплетного кода, названное вырожденностью, имеет очень важное значение, так как возникновение в структуре молекулы ДНК изменений по типу замены одного нуклеотида в полинуклеотидной цепи может не изменить смысла триплета. Возникшее таким образом новое сочетание из трех нуклеотидов кодирует ту же самую аминокислоту.
В процессе изучения свойств генетического кода была обнаружена его специфичность. Каждый триплет способен кодировать только одну определенную аминокислоту.
Интересным фактом является полное соответствие кода у различных видов живых организмов. Такая универсальность генетического кода свидетельствует о единстве происхождения всего многообразия живых форм на Земле в процессе биологической эволюции.
Незначительные отличия генетического кода обнаружены в ДНК митохондрий некоторых видов. Это не противоречит в целом положению об универсальности кода, но свидетельствует в пользу определенной дивергентности в его эволюции на ранних этапах существования жизни.
Наряду с триплетностью, вырожденностью, специфичностью и универсальностью важнейшими характеристиками генетического кода являются его непрерывность и неперекрываемость кодонов при считывании. Это означает, что последовательность нуклеотидов считывается триплет за триплетом без пропусков,
При этом соседние триплеты не перекрывают друг друга, т.е. каждый отдельный нуклеотид входит в состав только одного триплета при заданной рамке считывания. Доказательством неперекрываемости генетического кода является замена только одной аминокислоты в пептиде при замене одного нуклеотида в ДНК. В случае включения нуклеотида в несколько перекрывающихся триплетов его замена влекла бы за собой замену 2—3 аминокислот в пептидной цепи.

7. Этапы реализации генетической информации: Роль транскрипционных, посттранскрипционных и посттрансляционных модификаций в регуляции клеточного обмена. Примеры.
Транскрипция – происходит синтез иРНК на основе молекулы ДНК. Ключевой фермент РНК-полимераза.
1.РНК-полмераза распознает промотер и взаимодействует с ним. Промотр – особый участок гена, располагается перед кодирующей частью гена. Ф-ция – распознование промотора необходимо для активации РНК-полимеразы
2.РНК-полимераза после активации обеспечивает разрушение водородных связей между цепями ДНК на определенном участке молекулы.ж РНК-полимераза на основе кодогенной цепи ДНК обеспечивает синтез молекулы иРНК в соотвествии с принципами комплиментарности и антипараллельности. Кодогенной цепью является та епь ДНК на которой промотер располагается ближе к 3- концу
3.Когда РНК-полимераза достигает стоп-триплета, поступает сигнал о прекращении синтеза РНК. РНК-полимераза отделяется от ДНК, освобождается молекула РНК и восстанавливается структура ДНК. Синтезированная молекула РНК является проРНК (первичный транскрипт)
Процессинг-происходит созревание первичного транскрипта и формируется зрелая молекула матричной РНК
Сплайсинг – происходит вырезание интронов и сшивание между собой экзонов. Процесс сложный. В нем участвуют разные молекулы.
+
Альтернативный сплайсинг – у человека имеется около 25 000 генов и 100 000 белков. В соматических клетках организма гены одинаковые. На них образуется одинакове проРНК. Но в клетках разных тканей по-разному определяются границы и количество экзонов и
интронов. Поэтому в разных типах клеток образуются разные иРНК и синтезирутся разные белки
Трансляция – происходит сборка полипептидной цепи на рибосомах в соотвествии со строением мРНК
1.Инициация – начало процесса. мРНК с помощью КЭПа распознает малую субъединицу рибосомы. С помощью лидера формирует связь между мРНК и малой субъединицей рибосомы. К стартовому кодону присоединяется аминоацилтРНК,
которая несет а\к метионин. тРНК может присоединяться к мРНК только тогда, когда антикодон соотвествует кодону иРНК К сформировавшемуся комплексу присоединяется большая субъединица рибосомы. В целой рибосоме формируется 2 активных центра: Аминоацильный (А) и пептидийный (П)
2.Элонгция – Ацентр поступает аминоацил тРНК – антикодон которой сотвествует кодону мРНК. Эта аминоацил тРНК несет определенную а\к. Специальный ф-т образует пептидную связь между а\к. Рибосома двигается на 1 кодон относительно мРНК. Первая тРНК освобождается и уходит в цитоплазму. Ацентр свободен. В Пцентре находится тРНК, связанная с 2 а\к. Цикл многократно повторяется и с каждым шагом пептидная цепь увеличивается на 1 а\к
3.Терминация – завершение сентеза белка. В Ацентр поступает стоп-кодон. Его распознает специальный белок. Это является сигналом для прекращения синтеза белка. Разделяются субъединицы рибосом, освобождается мРНК и освобождается вновь образованный полипептид
Посттрансляционные изменения – при трансляции формируется первичная структура белка. Таком виде белок не способен выполнять свои функции
1.+Формируется вторичная структура за счет водородных связей
2.Формируется третичная структура за счет дисульфидных связей
3.У некоторых белков формируется четвертичная структура (гемоглобин)
4.Отщепляется метионин
5.Гликозирование белков (присоединение остатков сахаровантитела)
6.Отщепление от пептида определенных фрагментов – инсулин
8.Реакции матричного синтеза. Принципы и этапы репликации ДНК. Последствия нарушения нормального хода репликации ДНК.
При реакциях матричного синтеза образуются полимеры, строение которых полностью определяется строением матрицы. В основе реакций матричного синтеза лежит комплементарное взаимодействие между нуклеотидами. Основное функциональное значение репликации – обеспечение потомка стабильной генетическойинформациейразвития, функционирования и поведения. Полуконсервативнойпуть репликации ДНК. Установлено (М. Мезельсон, Ф. Сталь), что в процессе репликации две нити ДНК разделяются, каждая из них является шаблоном (матрицей) для синтеза вдоль нее новойнити. Последовательность оснований, которые должны быть в новых нитях, можно легко предсказать, так как они комплементарны основам, присутствующие в старых нитях. Таким образом, образуются две дочерние
молекулы, идентичные материнской. Каждая дочерняя молекула состоит из однойстарой(материнской) нити и однойновойнити. Поскольку только одна материнская нить сохранена в каждойдочернеймолекуле, такойтип репликации называется полуконисервативным. Каждая из двух цепейматеринской молекулы ДНК используется как матрица для синтеза новых комплементарных цепей. Репликация ДНК — ключевое событие в ходе деления клетки. Принципиально, чтобы к моменту деления ДНК была реплицирована полностью и при этом только один раз. Это обеспечивается определёнными механизмами регуляции репликации ДНК. Репликация проходит в три этапа:
1)инициация репликации;
2)элонгация;
3)терминация.
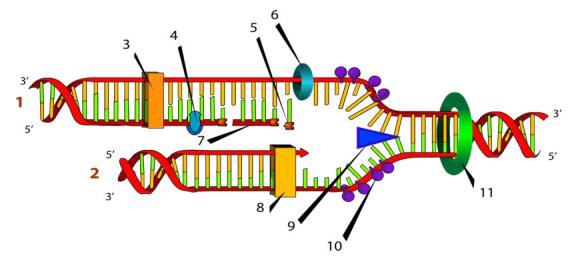
Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая нить, (3) ДНК-полимераза (Polα), (4) ДНК-лигаза, (5) РНК-праймер,
(6) праймаза, (7) фрагмент Оказаки, (8) ДНК-полимераза (Polδ), (9) хеликаза, (10) одиночная нить со связанными белками, (11) топоизомераза
Ферменты (хеликаза, топоизомераза) и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимеразы, способной распознать и исправить ошибку. Репликация у эукариот осуществляется несколькими разными ДНКполимеразами. ДНК-полимераза I действует на запаздывающей цепи для удаления РНК-праймеров и дорепликации очищенных мест ДНК. ДНК полимераза III — основной фермент репликации ДНК, осуществляющий синтез ведущей цепи ДНК и фрагментов Оказаки при синтезе запаздывающей цепи. Далее происходит закручивание синтезированных молекул по принципу суперспирализации и дальнейшей компактизации ДНК. Синтез энергозатратный. Цепи молекулы ДНК расходятся, образуют репликационную вилку, и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две новые двуспиральные молекулы ДНК, идентичные родительской молекуле.
В процессе репликации на каждой полинуклеотидной цепи материнской молекулы ДНК синтезируется комплементарная цепь. В итоге из одной двойной спирали ДНК образуются две идентичные двойные спирали. Такой способ удвоения молекул, при котором каждая дочерняя молекула содержит одну материнскую и одну вновь синтезированную цепь,