

Репарация без репарации - SOS-репарация –
индуцируемая репарация
Иногда в клетке активируются процессы, которые принято также называть репарацией, хотя на самом деле они являются средством осуществить
репликативный синтез ДНК, «не обращая внимания» на повреждение ее структуры.
Встречая повреждения в составе матрицы, реплисома обычно
останавливается. Если таких повреждений слишком много, и истинные
репарационные системы не успевают их исправить, переключение на неточный синтез ДНК дает клетке шанс на выживание.
Повреждения при этом остаются и, как следствие, дают большое количество мутаций. Все процессы такого типа обычно объединяют под названием SOS-репарации или (что точнее) - механизмы синтеза ДНК, толерантные к повреждениям (damage tolerance mechanisms).
Таким образом,
при SOS-репарации и
при пострепликативной (рекомбинационной) репарации,
повреждение остается
и может быть исправлено позже благодаря
эксцизионной репарации.
Репарация двунитевых разрывов
Двунитевые разрывы в ДНК возникают:
под действием ионизирующего излучения
под действием некоторых химических агентов, в частности, ингибиторов ДНК топоизомеразы II
Существует два основных пути репарации двунитевых разрывов: гомологичная рекомбинация
негомологичное соединение концов ДНК (NHEJ)
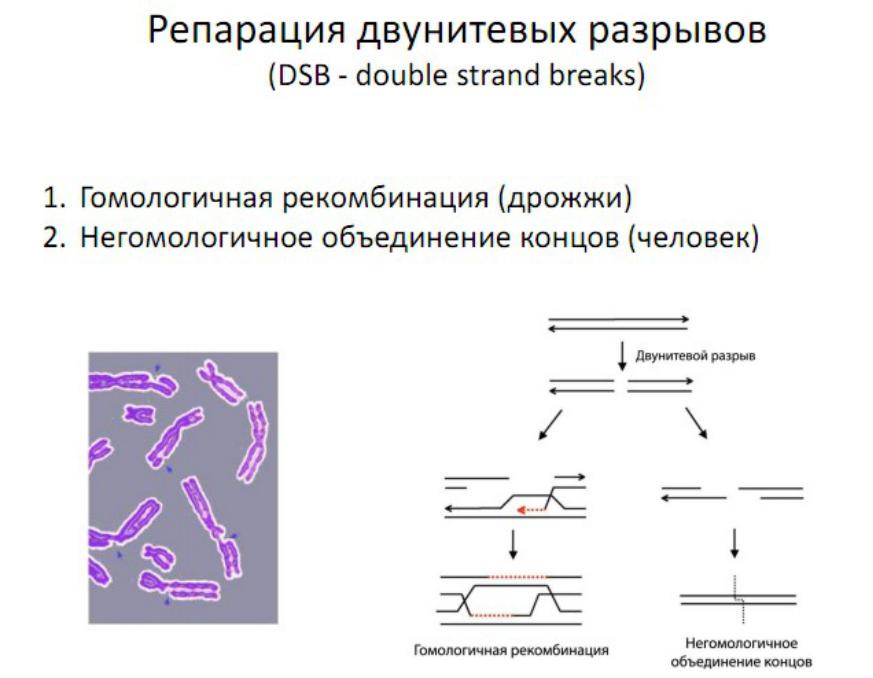

С рекомбинацией
Процесс репарации двунитевых разрывов разделяют на три этапа.
1. В первой, пресинаптической, фазе репарации происходит внесение двуцепочечного разрыва в ДНК или, при его наличии, сразу осуществляется
нуклеазное расщепление концов разрыва (процессирование концов экзонуклеазами).
В создании одноцепочечных 3'-OH-выступающих концов ДНК в месте разрыва принимает участие белок RecBCD, который обладает как хеликазной, так и экзонуклеазной активностями. RecBCD расплетает двухцепочечную молекулу ДНК в месте разрыва и гидролизует одну из цепей в направлении
5'->3', оставляя выступающий одноцепочечный участок.
2. Во второй фазе наблюдается синапсис гомологичных участков двух молекул ДНК с вхождением комплементарного одноцепочечного участка в ДНК-дуплекс (Инвазия 3’-конца первой цепи и Инвазия 3’-конца второй цепи) и последующим репаративным синтезом ДНК.
Далее происходит миграция ветвей с последующим образованием классической структуры Холлидея.
3. В третьей, постсинаптической, фазе репарации образовавшиеся структуры Холидея разделяются с помощью белков RuvA, B и C, RecG, а
также белков SOS-системы репарации (RecN, UvrD, RecF и RecJ).
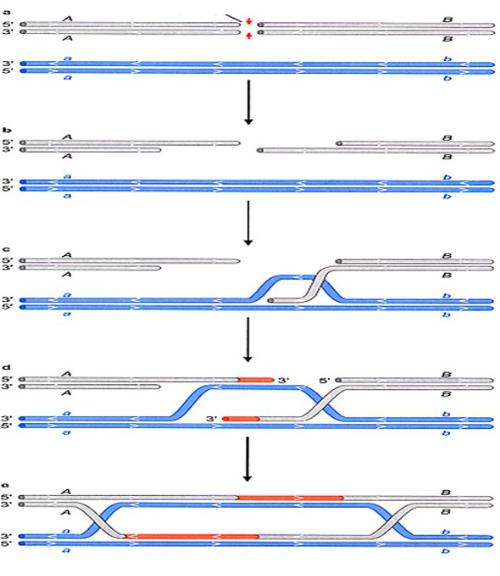
Репарация двунитевого разрыва посредством гомологичной рекомбинации
DSB
Процессирование концов экзонуклеазами; Создание выступающих 3’-концов
Инвазия 3’-конца первой цепи
Инвазия 3’-конца второй цепи и репаративный синтез
Миграция ветвей с последующим образованием классической структуры Холидея
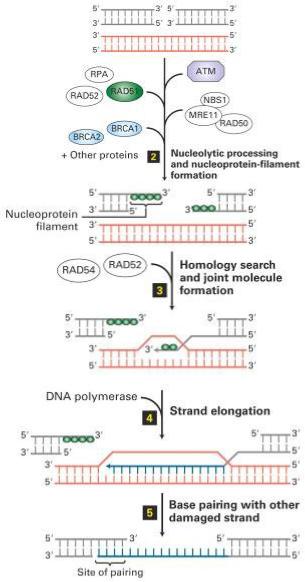
Homologous Recombination Repairs ds Breaks
В эукариотических клетках гомологом recA является
Rad51
Покрытая Rad51 однонитевая ДНК внедряется в гомологичный участок сестринской хроматиды с образованием D петли
3’-конец внедрившейся цепи достраивается ДНК полимеразой и отжигается с 3’-концом комплементарной цепи исходного дуплекса
Бреши застраиваются и однонитевые разрывы лигируются

Non-Homologous End Joining (Double Strand Breaks)
В других случаях целостность разорванных молекул ДНК восстанавливается в консервативном для всех организмов процессе негомологичного соединения концов (NHEJ - Non-Homologous End Joining). Соединяются при этом любые концы ДНК, что при наличии большого количества разрывов, приводит к объединению участков разных хромосом, транслокациям т.п.
Ключевую роль в процессе NHEJ играют белки Ku, которые
(в гетеродимерной форме) узнают концы ДНК, объединяют в нековалентный комплекс и рекрутируют к этому комплексу ряд других
белков.