

Вольвокс одноклеточный или многоклеточный
Организм относится к одноклеточным простейшим, которые способны создавать колонии. Внутри вольвокс соединены цитоплазматическими мостиками. Размер одной колонии достигает 2-3 миллиметров.
Строение вольвокс
Колония шаровидной формы имеет внутри от 70 тыс. до 75 тыс. клеток. Они расположены в один слой по периферии, а полость заполнена слизью студневидного характера. Каждая клетка обладает двумя жгутиками, а сзади расположен пристеночный хроматофор.
Соединение клеток происходит благодаря протопластам, которые связываются тонкими нитями. Клетки похожи на хламидомонады, имеющие округлые формы. Большинство клеток внутри колонии вольвокса являются вегетативными и питающимися. И только 7-10 клеток имеют способность к размножению.
Размножение и жизненный цикл вольвокса
Для колонии вольвокс характерно два вида размножения:
Половой
Бесполый.
Для первого способа – полового – характерно следующее: Происходит образование оогониев и антеридиев, которые могут развиваться в нескольких шарах или в одном из них. Оогонии состоят из 8-10 клеток, которые отличаются большими размерами, способствуют образованию одной неподвижной яйцеклетки.
Антеридии формируют множество двужгутиковых сперматозоидов. Когда происходит разрыв оболочки антеридия, то сперматозоиды направляются к оогонию, проникают туда, и порисходит оплодотворение яйцеклетки. Зигота находится некоторое время в состоянии покоя, покрываясь постепенно толстой оболочкой и заполняясь необходимыми питательными элементами.
После зимы начинается деление зиготы. Клетки, которые образуются в результате этого процесса, создают шар полого типа, а потом постепенно выходят наружу. В бесполом размножении принимают участие клетки, которые еще сохраняют способность делиться. Они создают пластинки клеток, которые при достижении нужных размеров, загибаются в чашеобразную форму.
Потом происходит выворачивание формы наизнанку, смыкаясь в полый шар. Он проваливается в центральную часть материнской колонии. В результате этого процесса происходит создание от 8 до 15 новых колоний. На периферических клетках формируются жгутики. Выход наружу происходит, когда молодые колонии достигают полного созревания. При этом материнская колония гибнет.
Интересные факты
Жгутики клеток ищут минеральные вещества в окружающей среде
Важное значение для развития и размножения играют нитраты и фосфаты, которые присутствуют в озерах и реках
Вольвокс потребляют в пищу бактерии, органику и растения, из которых они «готовят» пищу в процессе фотосинтеза. Колонии вольвокс перемещаются, за этот процесс ответственными являются периферийные клетки.
Движение происходит туда, где колония видит свет.
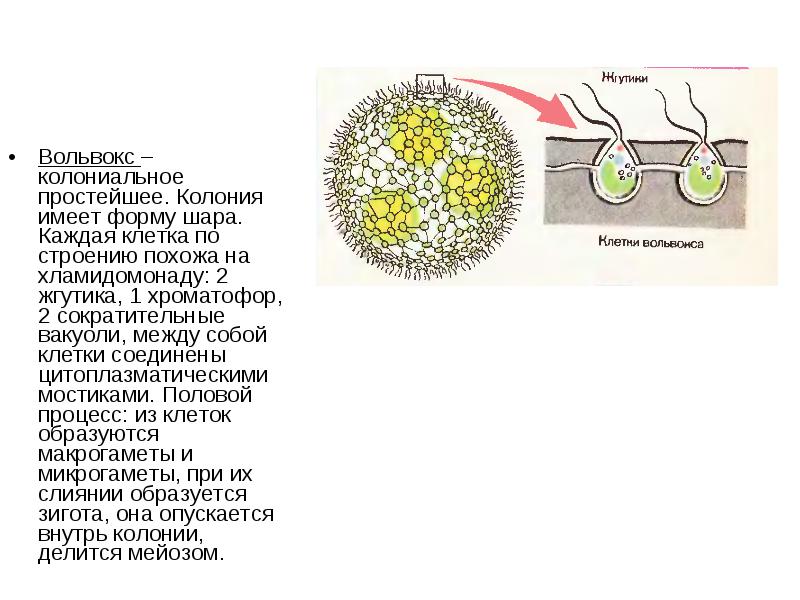
Вольвокс – род колониальных растений из отдела зеленых водорослей. В настоящее время изучено около 20 видов данного рода. Эти подвижные колониальные организмы распространены в пресных водоемах со стоячей водой. В период массового размножения окрашивают воду в зеленый цвет. Появляется так называемое «цветение» воды.
Строение. Колония вольвокса выглядит как небольшой подвижный зеленый шар (до 2-3 мм в диаметре). Каждая колония объединяет от сотен до десятков тысяч клеток вольвокса, расположенных на поверхности шара. Между собой клетки соединены особыми протоплазматическими нитями. Внутренняя полость сферы заполнена жидким слизистым веществом. Клетки вольвокса по строению сходны с хламидомонадой. Парные жгутики каждой клетки обращены кнаружи.
Размножение вольвокса осуществляется как бесполым, так и половым путем. На уровне колонии вольвокса отмечается специализация клеток. Основная масса клеток – вегетативные. Между ними находятся крупные генеративные клетки, участвующие в размножении. В оогониях образуются яйцеклетки, в антеридиях – сперматозоиды. После слияния гамет формируется зигота – ооспора. Прорастание ооспоры наблюдается в весенний период. В жизненном цикле вольвокса только у зиготы имеется двойной набор хромосом, вегетативные клетки гаплоидны.
В бесполом размножении участвуют особые клетки партеногонидии. Деление данных клеток осуществляется перпендикулярно к поверхности шара, в результате чего появляется дочерний шар.
Изучение вольвокса позволяет сделать научное предположение о том, что в процессе эволюции развитие живых организмов от одноклеточных к многоклеточным происходило через формирование колониальных форм. Эти растения вызывают большой интерес ученых как типичный пример колониальных форм среди водорослей.
2. Видоизменения листьев.
Колючки
Это видоизменение характерно для растений, обитающих в сухом и жарком климате, хотя нередко они возникают и у растений других климатических зон. Колючки уменьшают транспирацию и защищают растения от поедания животными.
Метаморфоз всего листа в колючку характерен для кактусов. У многих астрагалов, эспарцетов в колючку превращается рахис сложного листа, у белой акации — прилистники.
Есть растения, у которых листья видоизменены в усики. Этими усиками растение цепляется за опору и удерживается в вертикальном положении. Примером растениями с усиками являются горох, чина, вика и другие бобовые. У гороха в усики превращены верхние части листьев.
Это нитевидные образования, чувствительные к прикосновению и приспособленные для лазания. У вики, чечевицы, гороха в усик преобразуются верхняя часть рахиса и несколько верхних листочков. У чины безлистной все листочки редуцируются, а рахис представляет собой единственный усик (функцию фотосинтеза берут на себя прилистники.
Ловчие аппараты
Встречаются у растений, произрастающих на болотистых, торфяных, бедных минеральными веществами почвах. При помощи ловчих аппаратов росянка в средней полосе, венерина мухоловка в Северной Америке, непентес в тропической Азии используют богатую азотом и фосфором органическую пищу, переваривая животных. Строение ловчих аппаратов различно (кувшинчики непентесов, ловушки венериной мухоловки, листья росянки), но все они способны улавливать и переваривать насекомых и других мелких животных с помощью ферментов, выделяемых особыми пищеварительными железками.
Главными функциями листьев можно назвать фотосинтез и испарение воды. Чтобы выполнять эти функции наиболее эффективно, лист должен иметь форму пластинки, т. е. обладать большой поверхностью и быть тонким. Таковыми и являются листья большинства растений. Однако у некоторых растений листья в процессе эволюции видоизменились (претерпели изменения) и стали непохожими на обычные листья. Причина данного явления заключается в том, что листья начали выполнять другие функции, не связанные с фотосинтезом и испарением.
Листья ряда растений засушливых мест обитания видоизменились в колючки. Такие листья, с одной стороны, почти не испаряют воду, с другой стороны, защищают растения от поедания животными. Примером растений пустынных мест с колючками являются различные кактусы. Фотосинтез у них протекает в расположенных ближе к поверхности клетках толстого стебля. Также в стебле запасается вода. Таким образом, видоизмененные в колючки листья утратили обе свои главные функции (фотосинтез и испарение), но в замен стали выполнять защитную функцию.
Листья-колючки можно наблюдать не только у растений пустынь и полупустынь. Колючки есть у барбариса, кустов розы, шиповника и др. Однако у этих растений не все листья видоизменены в колючки, а только некоторые. При этом функция колючек здесь такая же как у кактусов — защита от поедания животными.
Листья других растений засушливых мест обитания приспособились к недостатку влаги иным способом. Так у алоэ и агавы листья видоизменились в толстые и сочные образования, в которых запасается вода. А чтобы уменьшить испарение такие листья покрыты восковым налетом, волосками, имеют меньше устьиц. В данном случае листья видоизменились так, что не утратили свои основные функции, при этом приобрели дополнительную — запасание воды.
В природе существуют насекомоядные растения. Их листья видоизменены в своеобразные ловчие аппараты. Когда насекомое садится на лист растения росянки, то приклеивается к нему, т. к. лист покрыт волосками, выделяющими вязкую массу. После этого волоски и сам лист сворачиваются. Внутри образовавшейся полости насекомое переваривается за счет выделяемых листом ферментов. Из насекомого росянка усваивает богатые азотом органические вещества. Именно из-за нехватки азота и других микроэлементов в местах своего обитания листья росянки приобрели столь специфическую функцию.
Еще одним видоизменением листьев являются чешуи. При этом чешуи бывают разными, т. к. выполняют различные функции. Например, у лука в луковице листья превращены в сочные чешуи, в которых откладывается запас питательных веществ. Другие чешуи покрывают почки. В этом случае они выполняют защитную функцию.
1. Строение и цикл развития представителей бурых водорослей на примере ламинарии.
У бурых водорослей встречаются все формы размножения: вегетативное,
бесполое и половое. Большинство бурых водорослей
встречается в виде двух самостоятельных форм развития - спорофита
и гаметофита, которые у разных видов бывают как сходного размера
и строения, так и различного. Иными словами, существуют изоморфная
и гетероморфная смены форм развития. При гетероморфной смене
форм развития одна из них бывает микроскопической, чаще всего
это гаметофит. Так происходит, в частности, именно у ламинарии,
где мы имеем дело с крупным многоклеточным спорофитом и невидимым
невооруженным глазом гаметофитом.
Размножение осуществляется спорами. Спорофиты всех ламинариевых
образуют одноклеточные одногнездные спорангии. Число
зооспор в них колеблется в зависимости от вида и от внешних условий
от 16 до 128. Образование зооспор сопровождается мейозом.
После созревания и выхода зооспор спороносные участки пластины
разрушаются. Массовое развитие спорангиев происходит в конце
лета - начале осени. Цикл годовой активности бурых водорослей
обычно таков: весна - начало роста и развития; лето - «расцвет»;
осень - замедление развития и отмирание листовидной пластинки;
зима - покой.

Рис. 14. Цикл развития ламинарии [15].
1 - зрелый спорофит с пятном спорангиев;
2 - зооспоры;
3 - эмбриоспоры;
4 - эмбриоспоры с проростковой трубкой;
5 - гаметофиты: справа - мужской, с вышедшим из антеридия антерозоидом;
слева - одноклеточный женский после выхода яйцеклетки из оогония;
6-7 - развивающееся слоевище спорофита.
1, 6, 7 - макростадии; 2-5 ~ микростадии
Число спор у ламинарии огромно. Одно слоевище длиной 250
см и шириной 20 см может дать до 36 млн спор. Зооспоры грушевидные,
длиной 5-10 мкм, с двумя жгутиками, отходящими сбоку, и
одним хлоропластом внутри. Строение их типично для подвижных
клеток бурых водорослей. Споры рассеиваются из зрелых талломов
в окружающую воду, разносятся течениями и оседают на субстрат.
При подходящих температурных условиях зооспоры способны к
активному движению в течение 2 суток. Не найдя места для прикрепления,
они погибают. Встретив твердую поверхность, зооспора
прикрепляется к ней передним жгутиком, который сокращается и
подтягивает тело зооспоры к субстрату. Затем зооспора теряет жгутики
и округляется, у нее появляется хорошо заметная оболочка, и
она превращается в эмбриоспору [31].
В течение 1-2 суток эмбриоспора дает вырост - проростковую
трубку, куда перетекает протоплазма, отделяющаяся затем перегородкой
от опустевшей эмбриоспоры. Так образуется первая клетка
гаметофита. Затем на гаметофитах будут созревать гаплоидные половые
клетки, которые сольются в диплоидную зиготу, и уже из зиготы
вырастет зрелое растение - спорофит (рис. 14). Оболочка эмбриоспоры
еще долго сохраняется у гаметофита, даже тогда, когда
на нем вырастает спорофит. Именно спора является той ключевой
стадией, от которой в конечном счете зависит урожай водорослей,
или многоклеточных спорофитов, ибо массовый выход спор крайне
важен для их успешного прикрепления (в соответствующее время)
и всего последующего развития культивируемых водорослей.
Микроскопические гаметофиты ламинариевых обычно представляют
собой однорядные разветвленные стелющиеся нити.
Мужские гаметофиты - многоклеточные, женские могут быть одноклеточными
и многоклеточными, но и многоклеточные не превышают
в диаметре 300 мкм [49]. Обычно они существуют не более
1-4 месяцев до созревания гаметангиев.
Половой процесс у ламинариевых - оогамия. Антеридри в виде
мелких бесцветных клеток группами или по одной располагаются
на поверхности вегетативных клеток. В антеридиях образуется по
одному антерозоиду, после выхода которых мужские гаметофиты
погибают. В случае многоклеточного женского гаметофита в оого-
нии обычно превращаются конечные клетки ветвей, в случае одноклеточного
- единственные клетки [49]. На севере гаметофиты ламинарий
зимуют на одноклеточной стадии и не растут из-за нехватки
света, в Северном море созревают в феврале, причем женские
гаметофиты остаются одноклеточными [49]. В каждом оогонии образуется
по одной яйцеклетке, которая выходит через отверстие на
вершине оогония, но не отделяется, а остается прикрепленной к
стенкам отверстия. В таком положении происходит ее оплодотворение
и рост спорофита. После выхода всех яйцеклеток гаметофит
отмирает, и на месте одного многоклеточного женского гаметофита
развивается несколько спорофитов. Женские гаметофиты не пре49
доставляют развивающимся спорофитам никаких питательных веществ,
но они обеспечивают им место на грунте. Это важно, так как
основная масса зачатков морских донных водорослей, спор и гамет
погибает из-за того, что не попадает на твердую поверхность [39].
Оплодотворенная яйцеклетка вырабатывает оболочку и начинает
расти, образуя вначале однорядную нить из 7 клеток. Затем,
наряду с поперечными перегородками, появляются продольные и
образуется однослойная пластинка спорофита. Из нижних клеток
пластинки вырастают тонкие ризоиды. По мере роста пластинки
увеличивается число слоев клеток, происходит их специализация,
появляются ствол и многорядные ризоиды. На этом этапе спорофит
представляет собой хорошо знакомое нам растение. Таким образом,
важно помнить, что известные нам водоросли - это зрелые спорофиты,
макроскопическая стадия жизненного цикла ламинариевых.
Споры, гаметофиты и ювенильные спорофиты относятся к микроскопическим
стадиям, не видным невооруженным глазом.
Ламинариевым - крупным водорослям - для роста нужны условия,
при которых обеспечивалось бы поступление питательных
веществ к слоевищам. Поэтому они обычно растут в местах с сильным
течением или прибоем, в водах, обогащенных питательными
веществами.
2. Растения-паразиты.
презентация
1. Внешнее и внутреннее строение мохообразных.
Завоевание растениями суши произошло 420 миллионов лет назад. По-видимому, первыми организмами, заселившими сушу, стали потомки зелёных водорослей. Выход на сушу – это качественный скачок в эволюции, типичный пример аромогенеза (последовательности ароморфозов), потребовавший от растений преодолеть следующие трудности:
- Интенсивные потери воды требуют приспособления для её добычи и запасания. У наземных растений развивается кутикула – защитный восковой слой, уменьшающий испарение воды.
- Половые клетки водорослей могли оплодотворяться только в воде. В ходе эволюции у наземных растений образовались различные приспособления, помогающие решить эту проблему.
- Воздух в отличие от воды не может служить опорой растениям. Тело наземных растений должно быть сравнительно жёстким.
- Растениям для роста и размножения требуется свет и минеральные вещества. Часть растения должна остаться над землёй и осуществлять фотосинтез, а другая часть – уйти под землю, снабжая растение водой и минеральными веществами и удерживая его в почве.
- Газообмен CO2 и O2 должен происходить не в воде, а в воздухе.
- Колебания условий окружающей среды (температура, влажность, концентрация различных веществ) более не сглаживаются водной средой.
Все эти приспособления мы можем наблюдать у современных мхов – наиболее примитивных организмов из группы высших растений. Моховидные (Bryophyta) – это отдел царства растений, объединяющий многоклеточные, просто устроенные растения, у которых отсутствует проводящая ткань (флоэма либо ксилема). Именно с последним фактом связаны небольшие размеры этих растений. Тело представителей этого отдела невелико (1–50 см) и слегка дифференцировано на условные «стебель» и «листья», хотя настоящих листьев и стеблей у них нет. Нет у мхов и корней, «стебель» прикрепляется к земле ризоидами – нитевидными выростами стебля, состоящими из одной или нескольких клеток (настоящие корни, в отличие ризоидов, содержат клетки проводящей ткани).
|
строение мхов |

Мхи достаточно плохо приспособлены к жизни на суше. Они растут, в основном, в сырых и затенённых местах, нередко на гниющей древесине или под водой. Мхи образуют основной растительный покров тундры и торфяных болот. Из-за почти полного отсутствия кутикулы поверхность таллома интенсивно испаряет воду; её недостаток восполняется за счёт поглощения всей поверхностью тела. Некоторые моховидные, однако, могут выдерживать продолжительные периоды засухи, используя какие-то не совсем понятные механизмы. Другие могут расти на открытых скалах.
Как и у всех высших растений, у мхов наблюдается чередование поколений. В течение одного жизненного цикла гаплоидный гаметофит в антеридиях образует сперматозоиды, которые с током воды по таллому достигают архегония и сливаются с яйцеклеткой в диплоидную зиготу. Зигота вырастает в диплоидный спорофит – поколение, размножающееся бесполым путём. В результате мейоза на спорофите появляются гаплоидные споры, из которых прорастают гаплоидные гаметофиты, и всё повторяется вновь. У мхов гаметофит является доминирующим поколением, а спорофит, развиваясь и питаясь за счёт гаметофита, полностью зависит от него. Споры образуются в споровой коробочке, возвышающейся над гаметофитом на тонкой ножке.
|
Чередование поколений у высших споровых растений |

Как мы убедимся в дальнейшем, «листья» и «стебли» мхов не являются настоящими стеблями и листьями не только из-за отсутствия сложного тканевого строения, но и потому, что развиваются на другой стадии жизненного цикла – на гаметофите, преобладающей стадии жизненного цикла мхов, а не на спорофитной стадии, как у сосудистых растений.
|
Так спорофит прикрепляется к гаметофиту |
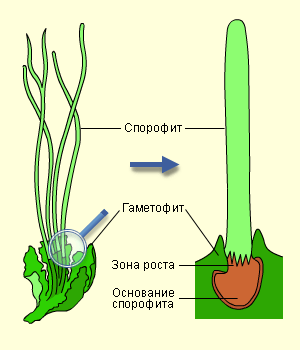
У мхов также хорошо развито вегетативное размножение при помощи специализированных почек и клубеньков. Почти каждая вегетативная клетка, если её изолировать от прочих, способна вырасти в самостоятельное растение.
2. Типы опыления покрытосеменных растений. Итак, опыление — процесс, без которого невозможно половое размножение растений. В ходе опыления пыльца переносится с тычинок на рыльце пестика, который ее захватывает. Существует два вида опыления: самоопыление и перекрестное.
1. Самоопыление — перенос пыльцы с тычинок на пестики одного цветка. Это крайне невыгодный для растений вид опыления, при котором гены не обновляются, сорта вырождаются. Но вот для селекционеров самоопыление может быть значимым — выводятся чистые сорта. Распространено самоопыление у таких растений, как фиалка, ячмень, соя, помидор, нектарин. Необходимое условие для самоопыления — одновременное созревание на цветках одного растения и тычинок с пыльцой и рылец пестиков. У некоторых самоопыляющихся растений этого не происходит, и тут уж начинается суета! Нужно, чтоб ветер или насекомые перенесли пыльцу с одних цветков, на которых вызрели тычинки, на другие, где вызрели рыльца. Впрочем, есть растения, которые могут размножаться и путем самоопыления, и с помощью перекрестного опыления, например, подсолнечник или крыжовник.
2. Перекрестное опыление — перенос пыльцы с одного цветка на другой, где она попадает на рыльце. Транспортировать пыльцу могут ветер, насекомые или, например, вода. Такой вид опыления преобладает в природе, и недаром: он наиболее эволюционно выгоден. Резко возрастает возможность для рекомбинации генетического материала, а это в свою очередь приводит к расширению внутривидового разнообразия, появлению крепких, живучих растений.
Признаки самоопыляющихся растений
1. Цветки не выделяют нектар и не имеют запаха, — действительно, для кого стараться? Насекомые все равно им для опыления не нужны.
2. Тычинки как правило находятся выше, чем пестики, — так пыльца точно попадет на рыльца.
3. У некоторых растений пыльца созревает еще в бутоне, и самоопыление идет в нераспустившемся цветке (гусмания, горох, арахис). У фиалки удивительной, недаром так названной, весной появляются красивые некрупные цветы, в которых нет особого смысла — они не дают семян. А вот летом у фиалки семена образуются в нераскрывающихся цветках. Различают два типа опыления – это перекрестное (естественное и искусственное) опыление и самоопыление. Рисунок 1. Самоопыление Самоопыление свойственно растениям с двуполыми цветками. Большинства растений при опылении дают семена. К растениям самоопылителям можно отнести такие растения как ячмень, овес, горох, просо. Самоопыление характерно для цветков, которые совсем не раскрываются, т.е. там, где невозможно перекрестное опыление. Таким образом, благодаря самоопылению, мелки невзрачные цветки дают семена. Потомство, полученное от самоопыления, является мало прогрессивным. Таким растениям непосредственно угрожает вырождение. Поэтому у растений-самоопылителей мизерный процент цветков должен подвергнуться внутривидовому опылению. Полученные от него растения имеют несколько другие отцовские и материнские зачатки, их приспособительные способности больше, они выживают в процессе естественного отбора. Все это предусматривает сохранению вида.
. Перекрестное опыление Замечание 1 Перекрестное опыление зависит от внешних факторов: птиц, насекомых, ветра и воды. Анемофилия – ветроопыление. Данное опыление характерно для растений у которых мелкие цветки часто собранные в соцветия. Цветки имеют достаточно много пыльцы. Пыльца сухая и мелкая, выбрасывается она наружу при помощи пыльника. При чем пыльник находится на длинных тонких нитях. Рыльца пестика широкие и длинные, высовываются из цветков, что способствует лучшему попаданию на них пыльцы. Такое опыление характерно для злаковых, и для тех у кого соцветие сережка, например, ольха, береза, орех, хмель, тополь. Для крапивы и конопли, у которых цветок состоит из чашелистиков, простого околоцветника, что не привлекают внимание опылителей. Энтомофилия – опыление насекомыми. Растения, которые имеют нектар, аромат, цвет и размер цветков, липкую пыльцу с выростами опыляются насекомыми. Насекомые переносят пыльцу из одного цветка на рыльца другого цветка, обеспечивая, таким образом, опыление для двуполых растений. Например, маки, ромашки, калина, шалфей ,молочай, гречиха и др.
Орнитофилия – опыление птицами. Такое опыление характерно для тропических растений с яркими цветками птичкой колибри. Гидрофилия – опыление водой. Водные растения, имеющие пыльцу и рыльце нитеобразной формы переносятся водой, или иногда слизнями. К водным гидрофильным растениям относят резуху, взморника, роголистку, наяда, элодею, рунию. Искусственное опыление В плодовом и декоративном садоводстве, овощеводстве, лесном хозяйстве применяется искусственное оплодотворение. Оно заключается в искусственном перенесении пыльцы с тычинок на рыльце пестика. Другими словами искусственное опыление называется скрещивание. Селекционеры, применяя метод скрещивания, получают новые виды и сорта растений.
Опыление у покрытосеменных растений Покрытосеменные растения опыляются обоими способами, при которых пыльцевые зерна непосредственно попадают на рыльца пестиков. Рассмотрим опыление на примере винограда. Для этого растения присущи два способа опыления – перекрестное и самоопыление. При самоопылении у винограда наблюдается клейстогамия. Клейстогамия – опыление, с последующим оплодотворением. В основном для винограда характерно опыление ветром (анемофилия), так как строение цветка приспособлен к такому перекрестному опыления. Во время опыления выделяется секретная жидкость на рыльце, в то время когда оно готово к получению пыльцы. Таким образом рыльце способствует прилипанию пыльцы, защищает ее от инфекции, и задает благоприятные условия для ее прорастания. Для винограда характерен переход от перекрестного опыления к самоопылению. Это способность сохраняет вид на протяжение многих столетий. Растения имея такую способность, будет всегда развиваться, и давать хороший урожай. Селекционеры прибегают к искусственному оплодотворению. Это в своем роде напоминает перекрестное опыление, но уже не с помощью биотических и абиотических факторов, а с помощью антропогенных.
1. Жизненный цикл мохообразных на примере листостебельных мхов.
Гаметофит
Листостебельные мхи, представители которых изображены на фото, мы привыкли видеть как сплошной зеленый ковер. Это и есть половое поколение растений. Если рассмотреть его поближе, можно увидеть, что он состоит из небольших стебельков с мелкими сидячими листиками линейного типа. Как и все мхи, они прикрепляются к субстрату ризоидами. На побегах листостебельных мхов формируются гаметангии, в которых формируются половые клетки. При наличии воды они сливаются, образуя бесполое поколение мхов - спорофит.
Спорофит
Бесполое поколение мхов развивается на зеленом гаметофите. Оно имеет вид тонкой ножки, на которой расположена коробочка. Внутри нее развиваются споры - клетки бесполого размножения. Когда коробочка открывается, они попадают в почву, прорастают и вновь образуют гаметофит, который преобладает в жизненном цикле моховидных растений. Далее половое поколение формирует зиготу, которая делится путем мейоза и образует споры. И таким образом, жизненный цикл осуществляется снова. Спорофит неспособен к самостоятельному питанию, поскольку практически не содержит в своих клетках хлорофилла. Именно поэтому он прикреплен к листостебельному гаметофиту, за счет которого питается. Отличаются эти поколения и хромосомным набором. У спорофита он диплоидный. А вот половое поколение имеет одинарный набор, поскольку половые клетки при формировании зиготы сливаются. М. относятся к числу растений с чередованием поколений. В их жизненном цикле часто преобладает гаметофит (половое поколение, им является практически всё растение). Наряду с осн. предназначением (образованием половых клеток) гаметофит участвует также в выполнении вегетативных функций – в фотосинтезе, водоснабжении и минер. питании. Диплоидный спорофит (бесполое поколение) в течение всей своей жизни прикреплён к гаметофиту и получает от него воду и питат. вещества. У большинства М. спорофит значительно мельче гаметофита и выглядит как спорообразующий орган (отсюда второе название спорофита – спорогоний
У мн. листостебельных мхов быстро растущая ножка вызывает обрыв ткани в основании архегония, который остаётся в виде колпачка на верхушке ножки. Впоследствии под колпачком развивается коробочка. У андреевых и сфагновых мхов ножка остаётся короткой, под спорогонием развивается ложная ножка – продолжение стебля гаметофита; архегоний разрывается поздно из-за разрастания самой коробочки. У печёночников ножка спорофита остаётся короткой до созревания спор, а затем за неск. часов сильно удлиняется благодаря насасыванию воды и растяжению своих клеток, вследствие чего архегоний разрывается и коробочка оказывается во внешней среде. Длинные истинные или ложные ножки выносят коробочку из развивающихся вокруг архегониев защитных образований и тем самым обеспечивают беспрепятственное рассеивание спор.
Споры и спорообразование. Споры образуются в коробочке в результате мейоза и потому содержат гаплоидный набор хромосом в ядре. У антоцеротовидных и печёночников в коробочке наряду со спорами развиваются диплоидные нитевидные элатеры, гигроскопич. движения которых разрыхляют массу спор. У антоцеротовидных и листостебельных мхов по оси коробочки располагается тяж стерильной ткани – колонка. Коробочка со зрелыми спорами вскрывается продольными щелями (андреевые мхи), створками (мн. печёночники), кольцевой поперечной щелью, отделяющей крышечку (сфагновые и зелёные мхи), неправильно разрывается или сгнивает (некоторые печёночники). По краю устья вскрывшейся коробочки у зелёных мхов обычно хорошо заметны располагающиеся в 1 или 2 ряда зубцы, в совокупности составляющие перистом и участвующие в рассеивании спор. Обычно споры имеют сферич. форму, мелкие (5–50 мкм в диаметре), распространяющиеся воздушными потоками, у большинства М. одинаковые в пределах вида (изо- или гомоспоровые растения), но у некоторых видов в одной коробочке развиваются в равном числе крупные и мелкие споры (анизоспоровые растения).
2. Многообразие покрытосеменных растений на примере семейств Губоцветные и Березовые.