

SRJ10262Volume501
.pdfSee discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/231520522
Priestorová a časová diverzita potravy sovy obyčajnej (Strix aluco)
Article in Slovak Raptor Journal · January 2011
DOI: 10.2478/v10262-012-0057-8
CITATIONS |
READS |
23 |
841 |
1 author:
Ján Obuch
Comenius University Bratislava
81 PUBLICATIONS 1,123 CITATIONS SEE PROFILE
Some of the authors of this publication are also working on these related projects:
Diet of the Eagle-Owl View project
Estimating density, population size and dynamics of Common Buzzard (Buteo buteo) in the West Carpathian region by a new method ,On foraging ecology of the Scops owl Otus scops on the northern limit of its distribution View project
All content following this page was uploaded by Ján Obuch on 26 May 2014.
The user has requested enhancement of the downloaded file.

Slovak Raptor Journal 2011, 5: 1–120. DOI: 10.2478/v10262-012-0057-8.
© Raptor Protection of Slovakia (RPS)
Spatial and temporal diversity of the diet of the tawny owl (Strix aluco)
Priestorová a časová diverzita potravy sovy obyčajnej (Strix aluco)
Ján OBUCH
Abstract:The author compared diet of eight owl species in Slovakia, out of which four species prey in the non-forest environment (Bubo bubo, Tyto alba, Asio otus and Athene noctua), while the other four species prey mostly in the forest (Glaucidium passerinum, Aegolius funereus, Strix uralensis and Strix aluco). Tawny owl (Strix aluco) has shown the highest degree of adaptability when it comes to various types of environment and broad diversity of prey.Appendix 1 presents material of total 225,441 pieces of diet, which contains 69 species of mammals and at least 147 species of birds. While B. bubo diet is typical especially for high presence of bigger prey species within mammals and birds (66 diagnostic taxa), diet of S. aluco is characterized by as many as 89 taxa with higher share than the average of all eight compared owls (taxa with values 1+ and 2+ in blocks as marked by full line). Based on the analysis of 68,070 pieces of S.aluco diet collected in Slovakia, it is possible to differentiate seven basic diet types: A– from lower mountains, B – from middle montane locations, C – from colder and more humid parts of mountains, D – characterized by high share of bats (Chiroptera), E – characterized by high share of slugs (Limacidae), F – from floodplain forests, and finally G – from environment strongly influenced by humans. Diagnostic species for one or several diet types are characterized by markedly higher share than Slovak average. By the large amount of analyzed samples of S. aluco diet it is possible to gain the knowledge about structure of several animal groups from relatively little disrupted forest ecosystems and those from environment to some degree influenced by humans. Diet types represent simplified models, which are understood in a different sense than plant communities. Individual disposition for specific food preference (e. g. Chiroptera) also plays an important role by some tawny owl’s specimens. Some types of prey can be further divided into undertypes (e. g. undertypes G1 and G2 in Table 1). Diet of S. aluco has been examined more in detail and over a longer period of time especially in following national parks: Slovenský kras, Muránska planina and Veľká Fatra Mts. Samples from other parts of Slovakia are also presented here according to their relevance to particular diet types. Pellets of S. aluco were collected over a longer period of time (up to 30 years) in several sites and it is chronological periodicity in presentence of diagnostic species, which stands in focus. Long-term changes in S. aluco diet during Holocene were examined in two parts of Veľká Fatra Mts, which are influenced by climatic changes as well as human activity (deforestation and pasture). The author has occasionally collected pellets of S. aluco in several mountains of Europe between WesternAlps and Caucasus and from northern border in S. aluco areal, from Trondheim area in Norway. Furthermore, the author presents hereby his own material from pellets of S. aluco from the Middle East to Himalaya (in particular countries of Jordan, Israel, Lebanon, Syria, Turkey, Iran, Kyrgyzstan and Nepal).
Abstrakt:ZúzemiaSlovenskaautorporovnávapotravuôsmichdruhovsov,zktorýchštyridruhyloviavnelesnomprostredí(Bubo bubo,Tytoalba,AsiootusaAthenenoctua)aštyridruhyprevažnevlese(Glaucidiumpasserinum,Aegoliusfunereus,Strixuralensis a Strix aluco). Sova obyčajná (Strix aluco) je najprispôsobivejšia na rôzne typy prostredia a na široké spektrum druhov koristi. Prezentujesamateriál225441kusovanalyzovanejpotravy,vktorombolozistených69druhovcicavcov(Mammalia)aminimálne 147 druhov vtákov (Aves). Kým B. bubo sa vyznačuje najmä početnejším zastúpením väčších druhov koristi (Mammalia aAves; 66 diagnostických taxónov), v potrave S. aluco je až 89 taxónov s výrazne vyšším podielom v potrave, ako je priemer od porovnávaných osem druhov sov. Na základe analýzy 68 070 kusov potravy Strix aluco z územia Slovenska možno diferencovať sedem základných potravných typov: A – z nižších pohorí, B – zo stredných horských polôh, C – z chladnejších a vlhších častí pohorí, D–svysokýmpodielomnetopierov(Chiroptera),E–svysokýmpodielomslizniakov(Limacidae),F–znížinnýchlužnýchlesov, G–zantropickysilneovplyvnenéhoprostredia.Určitývplyvpriniektorýchtypochmáindividuálnadispozícianauprednostňovanie špecifických druhov koristi (napr. Chiroptera). Niektoré typy potravy sa delia na podtypy (napr. podtypy G1 a G2.). Z dvoch častí Veľkej Fatry sa vyhodnocujú dlhodobé zmeny v zložení potravy S. aluco počas holocénu, ktoré sú ovplyvnené klimatickými zmenami, ale tiež antropickou činnosťou (odlesňovanie a pastva). V práci sa analyzujú aj vzorky potravy S. aluco z niektorých pohorí Európy (od Západných Álp až po Kaukaz), zo severnej hranice areálu druhu v Nórsku (Trondheim) ako aj materiál z ázijských pohorí (od Blízkeho Východu po Himaláje: Jordánsko, Izrael, Libanon, Sýria, Turecko, Irán, Kirgizstán a Nepál).
Key words: Tawny owl, Strix aluco, diet, Slovakia, Europe,Asia.
Ján Obuch, Botanical Garden of Comenius University, SK–038 15 Blatnica, Slovakia. E-mail: obuch@rec.uniba.sk.
1

Obuch J: Spatial and chronological diversity of tawny owl (Strix aluco) diet
Acknowledgement: Karel Čapek wrote in his novel Krakatit: “Whatever a man encounters, comes from within!”. Throughout my life I have been accompanied by a need to understand nature’s secrets. I have encountered precious personalities, who have led me to the path of studying the owl’s diet. By chance I found owl pellets and brought them to J. Sládek, who was a professor in zoology at the forestry college in Zvolen. After my university studies I met H. Schaefer, who was a follower of the interwar German “pellet school”.At my first workplace at the Považské Museum in Žilina I received into my custody the rich local zoological collections fromA. Štollman and undisturbed I could learn to identify animal species from their bone samples. In the 1980’s I started to cooperate on the research of owl diets in the Czech Republic together with B. Kloubec, J. Zima and J. Červený, with whom I experienced my first zoological expeditions to southern Kyrgyzstan, being invited by the local zoologist S. N. Rybin. In the 1990’s thanks to Z. Hodková I could join the research team of the Faculty of Natural History at Charles University in Prague. With this team I participated in ten expeditions to the Near East countries. I owe my gratitude to my wife Dana, who had great understandingformynightlysittingoverbonefragments.MydaughterZoranegotiatedmycooperationwiththeRoyalSocietyfor the Conservation of Nature (RSCN) on the research of Jordanian nature reserves. My daughter Barbora spurred my interest for the research of owl diets in Norway, and helps me in my projects with local ornithologists, especially G. Bangjord from Trondheim. I am thankful to my colleagues at the Botanical Gardens of Comenius University in Blatnica for creating a very stimulating environment.Especiallythedirectorofthisinstitution,D.BernátováandthedirectoroftheBotanicalGardensofComeniusUniversity in Bratislava, J. Bella have provided me with great help in negotiating foreign country visits. I owe my gratitude to M. Uhrin and M. Dravecký for proofreading this manuscript.
Introduction
The tawny owl Strix aluco is an inhabitant of autochthonous deciduous and mixed forests. It is a typical ‘generalist’ species, very adaptable to different food sources andenvironmentalconditions.Jirsík(1949)quitesharply characterizes the hunting abilities of the tawny owl: “We canbutsaythatnootherraptorhuntsitspreyinsuchversatilewaysandwithsomanytalents.”Thankstoitslarge distribution range and rather high density, S. aluco is an optimalmodelspeciesforthedeterminationofspatialand temporal diversity of species that comprise its prey.
Owl pellets often contain rather well preserved undigestedremainsofprey.Herewecanfindmostfrequently bones of vertebrate species and sclerotized body parts of invertebrates. Pukinskij (1977) put forward: “Without exaggeration we can claim that nine tenths of our knowledge about owl dietary relations comes from the identification of owl pellet contents.”
The identification procedures for mammals based on jawboneswaselaboratedinthe19thcentury.Theanalysis of owl pellets as a method of owl diet research was first usedbytheGermannaturalistAltum(1863).Thismethod was further systematically developed by Uttendörfer (1939, 1952). He analyzed mostly owl pellets of A. otus, T. alba and S. aluco. He trained several students who werededicatedtoamorethoroughdeterminationofprey species not only of the class Mammalia, but also Aves, Anura and Evertebrata. März (1969) wrote a methodical handbook about the collection and identification of owl pellets and food remains. The works of Uttendörfer also includeddataonthedietofT.albaandB.bubofromSlo-
vakia, which were collected and identified by his student Schaefer (1933, 1938)
In the post-war period, owl pellets from the southern partofSlovakiawereanalyzedbytheCzechauthorsBalát (1956)andFolk(1956)andSlovakauthorRandík(1956). In the 1960s the stomach contents of deceased and shot owlswereanalyzedbySládek(1962).Thesummaryofall dataonthedietofraptorsandowlsusingthismethodwas presented in the work of Kropil & Sládek (1990). In the 1970s other Slovak authors started collecting owl pellets includingSchmidt&Štollman(1972),Ginter(1971)and Darolová (1976). The pre-war efforts in the Belaer Tatra Mts. were followed by Schaefer (1972, 1974). Jánossy &Schmidt(1970)intheircompoundwork aboutthediet of B. bubo from the whole Palearctic region presented further records of about 100 pieces of food from the Slovenský kras Mts. (Rožňava) and Malá Fatra Mts. (Terchová). Owl pellets were collected by Štollman and identified by Schmidt. Until the second half of the 1970s we have had no specific data on the diet of S. aluco from owl pellets. Summary data from 32 locations in eastern Slovakia was later published by Danko (1989).
The greatest collection of material on the diet of S. aluco, from 261 locations, was published by Uttendörfer (1939) based on material collected between 1896 and 1938 (Jirsík, 1949). In his later work, this data was updatedwithmaterialfrom1939to1949andsummarized about 58,500 pieces of animal food items (Uttendörfer 1952). This material contained about 30,000 mammal individualsof45species,7,500birdsof100species,7,300 amphibians of eleven species, 400 fish of ten species and
2
Slovak Raptor Journal 2011, 5: 1–120. DOI: 10.2478/v10262-012-0057-8. © Raptor Protection of Slovakia (RPS)
almost 2,000 invertebrate individuals. This author also recorded 41 slugs of the genus Limax. According to the data summary the vertebrate prey items were dominated by voles of the genus Microtus and birds of the species Passer domesticus. Therefore we can conclude that Uttendörfermainlyanalyzedowlpelletsfromindividuals huntinginstronglyhumaninfluencedenvironments.Most European authors analyzed pellets of S. aluco coming from such environments: e. g. Delmee et al. (1979) in Belgium; Smeenk (1972) in the Netherlands; Southern (1954) in England; several studies from Germany (e. g. März 1954, Snurre & März 1970, Wendland 1980) and Poland (e. g. Goszczynski et al. 1993, Zalewski 1994, Bogucki 1967). In Austria, S. aluco diet was studied by Steiner (1961), in Czech Republic by Plesník & Dusík (1988), in Bulgaria by Simeonov (1985), in Slovenia by Lipej (1988), in Italy Contoli & Sammuri (1978), in Russia Bozsko (1967), in Estonia Balčianskione et al. (2000) and in Hungary Schmidt et al. (1971). Several of the presented works were devoted only to Mammalia and rodents such as Microtus and Apodemus sp. were determined only to genus level.
The analysis of a large number of collections from different locations shows that the increased frequency of some taxons occurs in specific environments. This has leadtotheclassificationofdietarytypeswhichbecamean original addition to our knowledge about the fauna of the forestecosystems(Obuch1992a),furthersupplementedby thedescriptionofdietarytypefromthelowlandfloodplain forests (Obuch 2003), as well as typical aspects related to human impact on the environment (Obuch 2004a).All seven types were first presented at the international conferenceonowlsintheGermanHarz(Obuch2000a).Long term temporal changes were analyzed at four locations on the Muránska planina Mts. (Obuch 1997). The much scarcer material of this author from some European and Asian mountain ranges indicates the presence of several endemicspecies,characteristicofisolatedforestenclaves inthelargerangefromtheWesternAlpstotheHimalayas. Data on the collection of S. aluco pellets from 20 Palaearcticmountainrangeswerepresentedattheinternational conference on birds of prey in Mikulov (Obuch 1999). LargesetsofsamplesfromtheCzechRepublicwereanalyzed (Obuch 1994a, Zvářal & Obuch 1996, Reiter et al. 1997, Zima et al 1998) and results from Bulgaria (Obuch & Benda, 1996) and Turkey (Obuch, 1994b) were also published. Currently a research project on the diet of S. aluco from nest box beddings in the central Norwegian region of Sør Trondelag is approaching completion and was referred to at its outset (Obuch & Bangjord 2008).
Themainaimsofthisworkwere:(1)todeterminediagnosticspeciesinthedietofS.alucoincomparisonwith thedietofotherowlspeciesinSlovakia;(2)toclassifyS. aluco diet in Slovakia into the main dietary types; (3) to identify spatial and temporal diversity in the individual types of S. aluco diet and (4) to present the results of the study on S. aluco diet from the European andAsian parts of its distribution range.
Material and methods
Most of the author’s data on the owls’ diet is based on the analysis of owl pellets. In case of S.aluco this means especially pellets collected at sites of the owls’ diurnal shelters in the rocks and buildings. Part of the material maycomefromthenestboxbedding,whereitaccummulates during young feeding. Bedding may contain parts of unconsumed stored prey and deceased or consumed young.As we compared the three different types of food item identification (Zvářal & Obuch 1996), we found certain differences. In case of A. otus most owl pellets arecollectedinthewinterperiodandshowlowerspecies diversity than samples from the nesting period (Šotnár & Obuch 1996). In B. bubo the most common material comes from nest bedding amongst rocks, which could be accummulated over decades. Older strata cannot be separated, since during digging of the nesting hole, the nestmaterialismixedup,underminingtheintegrityofthe samples.InAegoliusfunereus,themostcommonmaterial comesfromnestbeddinginnestboxesornaturaltreecavities hollowed out by the black woodpecker.The material fromtheGlaucidiumpasserinuminSlovakiawasmainly collected from the winter storage in the nestboxes.
The owl pellets or nest beddings from each sample weresoakedforonehourinhot5%NaOHsolution.After dissolving hair and feather debris, the sample was rinsed on a dense sieve under running water and in a container withstillwater.Floatingbodypartsofinsectsandhollow bird bones were collected and using careful circular washing motionalldebriswerewashed outuntilonlybones and snail shells remained.These were further cleaned by washing the sieve with circular motion to remove stones with different weight. After drying the washed sample, we sorted out the jaw bones (maxilla and mandible, as well as some teeth) of mammals (Mammalia), beaks, tarsometatarsi, humeral and metacarpal bones (rostrum, tarsometatarsus, humerus and metacarpus) of the birds (Aves), iliac bones (os ilium) of the frogs (Anura), jaws of reptiles and salmonid fish (Reptilia, Salmonidae), pharyngeal teeth (os pharyngeum inferior) of carp fish (Cypriniformes), heads (caput) or jaws of different in-
3

Obuch J: Spatial and chronological diversity of tawny owl (Strix aluco) diet
sect orders, dorsal shell lenses of slugs (Limacidae) and pincers of crayfish and crabs (Decapoda).
Thenumberofindividualsofeachtaxonwascalculated as the number of the most frequent body parts in the sample(e.g.leftorrightmandibleormaxillainmammals or reptiles, one out of four types of bones in birds, left or right iliac bone in frogs; Fig. 1, 2).
Theidentificationofboneswascarriedoutaccording to the reference collections of vertebrate skeletons from captured or deceased animals and according to the published identification characteristics (Anděra & Horáček 1982,Gromov&Erbajeva1995,Mendellsohn&Yom-Tov 1987, Harrison & Bates 1991, Lay 1967, DeBlase 1980, Kryštufek &Vohralík 2001, 2005, 2009, Mitchell 1975). Individuals from the Pannonian populaton of Apodemus uralensis (Pallas, 1811) were referred to as Apodemus microps Kratochvíl & Rosický 1952. These individuals couldbedistinguishedfromtheA.uralensisfromeastern Europe and Asia on the basis of smaller jaw bones. Due to extensive bone fragmentation, I could not always differentiate C. corone/cornix and Corvus frugilegus, therefore I refer to them using a shared name C. corone +frugilegus.Somebirdtaxonsweredeterminedtogenus level whilst lower vertebrates and invertebrates were determinedtothefamilyororderlevelandarereferredtoas „sp.“ (e. g. Apodemus sp. or Coleoptera sp.). In cases of dubious identification with a certain level of probability, the taxon is referred to as cf. (e. g. Mus cf. musculus).
Data on the owl diet in this work are evaluated using the calculations of marked differences from the mean (MDFM, Obuch 1991, 2001a). Samples in the adjusted
results tables are sorted according to the similarity and the ordering is adjusted such as the determining species with positive MDFM values arranged in columns and blocks. These blocks are enclosed in continuous line borders. Species without MDFM are arranged under a dashed line and ranked down according to total abundance. Diversity indices H’ (Shannon & Weaver 1949) are given in the bottom row of the table. Less abundant and partially determined taxa are given under the table. Calculations of the MDFM and contingency tables were carried out in the Zber software application (Šipöcz 2004).
Names and boundaries of the geomorpological units ofSlovakiawereclassifiedaccordingtoMazúr&Lukniš (1980). The overview of locations where food items of S. aluco were found is presented in the Appendix 41. Bone samples found in sites from the Pleistocene period or earlier are referred to as fossil whilst samples found in sites from the earlier Holocene period are referred to as sub-fossil. ‘Recent‘ finding places are from current period, whereas sub-recent are close to recent but with certain environmental differences. Ložek (1973) in his geological classification of the Holocene period referred tothelast500yearsastherecentperiodandtotheMiddle Ages (500–1400) as the subrecent period. In the bone finding places of S. aluco we can distinguish preserved pellets (less than five years old), recent detritus from the disintegrated pellets in the 2 cm surface layer (5–50, 200 years at maximum) and bones from the layers at 2–5cm,maximum10cm(50–200,atmaximum500years ago) from the sub-recent period.
J. Obuch
Fig. 1. Sorting bones after pellet processing with NaOH. Obr. 1. Triedenie kostí po spracovaní vývržkov v NaOH.
J. Obuch
Fig. 2. Determination of bird bones from the pellets. Obr. 2. Určovanie kostí vtákov z vývržkov.
4
Slovak Raptor Journal 2011, 5: 1–120. DOI: 10.2478/v10262-012-0057-8. © Raptor Protection of Slovakia (RPS)
Results and discussion
Comparison of Strix aluco diet with dietary ranges of other owl species in Slovakia
In order to describe differences in the hunting strategy of eight owl species in Slovakia, we have summarized mainly the data of this work’s author, supplemented by partially published data on Tyto alba from Borská nížina Lowlands(Noga2004).Fromthetotalnumberof225,441 food items with at least 250 taxa we have included in our analysis those diagnostic taxons, which are recorded in one or several owl species (Appendix 1). As first we list the most frequently occurring species with positive MDFM in T. alba,Asio otus,Athene noctua, Glaucidium passerinum,AegoliusfunereusandStrixuralensis.Atotal of89diagnostictaxonswerefoundinS.aluco.Thewater pippit (Motacilla alba) is equally distributed in the diet ofalleightowlspecies.Other,lessabundantpreyspecies
(∑n<20)aregivenunderthetable.ThedietofBubobubo asthelargestowlspeciesistypicallycomposedofgreater proportionoflargermammalandbirdspecies(especially non-passerine birds and corvids). Among the owls, this species hunts the most other predators, i. e. carnivores, raptorsandowls(Obuch2000b).Themostfrequentlower vertebrate species in the food items of B. bubo is the Rana temporaria, Lacerta sp. and Cypriniformes. As in S. aluco, its food also comprises Salmo trutta and some medium sized passerine birds, e. g. Turdus sp.
The diet of T. alba is distinguished by its high proportion of synanthropic and non-forest species of mammals and passerine birds. In three cases we found a high proportion of bats in its diet (Obuch 1998a). The most prominent hunting specialization in agricultural land can be seen in A. otus. Winter pellets of this species are dominated by 89% of M. arvalis remains. A. noctua also forages exclusively in non-forest environments, but
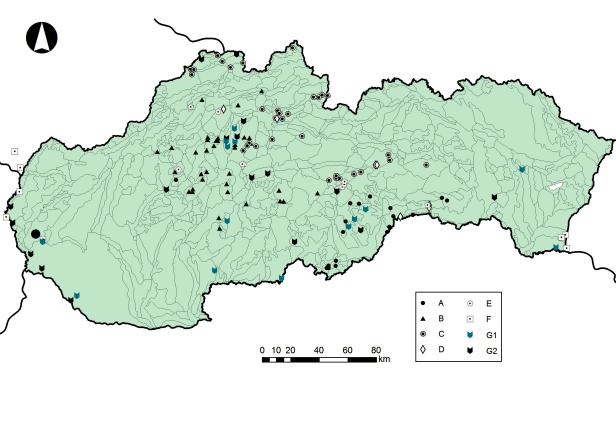
Fig. 3. Geographical distribution of the food samples of S. aluco and their classification into dietary types in Slovakia. For dietary types description see text. Author: F. Tulis.
Obr. 3. Poloha vzoriek potravy S. aluco s ich zaradením do potravných typov na Slovensku. Opis potravných typov je uvedený v texte. Autor: F. Tulis.
5
Obuch J: Spatial and chronological diversity of tawny owl (Strix aluco) diet
its diet is characteristized by a higher proportion of invertebrates (52%). This proportion is even higher in arid countries (Obuch & Krištín 2004).
The four other owl species hunt mainly in forest habitats. G. passerinum is typical for its high proportion of passerine birds in its diet (48%).Ahigh proportion of insectivore mammals and rodents can be found in the diet of A. funereus (95%). The diet of the S. uralensis in Slovakia has been little studied, but besides continuous forestcover,thisspeciesalsoentersforestpatchesenclosedbyfields(Košicebasin,Mihókinverb.).Thisspecies can winter in urban areas (e. g. Košice), hunting mainly doves and pigeons (Dravecký & Obuch 2009).
ThedietofS.aluco appearstobethemostvariablein thecontextoftheeightowlspeciesinSlovakia(H’=3.05) with the highest number of differential species from the classesMammalia,AvesandAmphibia.Aratherhighproportion of Limacidae (11%) in its diet is quite unique.
Dietary types of Strix aluco in Slovakia
Theanalysisofagreatnumberoflargesamplesusingthe MDFM method, has allowed us to find several species with positive MDFM appearing in many samples. These diagnostic species characterize a certain habitat or hunting behaviour.Thus we can describe a group of samples with common diagnostic features as a specific dietary type. A single sample can represent pellet collection from a single site, or several collections from the same site or several collections from several sites with similar prey species composition (e. g. from the same mountain range).Bysummarizingseveralcollectionsfromthesame site we can equalize the fluctuations in prey population cycles and the collective sample can better characterize the food niche at the sampling location. Food samples not large enough for their respective regional units were omitted from the analysis.
ThefirstdifferentiationoffoodsamplesfromS.aluco into dietary types was based on a set of 38 samples from mountain regions of Slovakia. This data set was evaluated by the MDFM method (Obuch 1992a). Dendrogram of similarity classified the diagnostic species into five groups, which were described as dietary types A, B, C, D, E. Three of these groups reflected altitudinal zones: A – lower mountain ranges, B – middle mountain zone, C – cold and wet mountains. The other two types were determined by the local specialization of S. aluco due to habitat conditions: D – Bat hunting specialization (Chi- roptera),E–frequentconsumptionofslugs(Limacidae). After analysing additional samples from the plains and
basins of Slovakia, we could differentiate two other die- tarytypes:F–floodplains(Obuch2003)andG–strongly human-influenced habitats (Obuch 2004b).
IntotalwecoulddifferentiatesevendietarytypesofS. aluco in Slovakia (Tab. 1, Fig. 3), categorized according tosimilarityindistributionofdiagnosticspecies.E–type was characteristic for a higher proportion of Limacidae in comparison with other dietary types. Some diagnostic specieswerecommonwithtypesCandB.Samplesofthis typecamemainlyfrombeechforestsofthe4thvegetation zone on calcareous ground (forest communities such as Fagetumdealpinum,Hančinský1972)onthesouthernside ofMuránskaplaninaMts.andinthevicinityofHarmanec in Veľká Fatra Mts. Samples from the warmer southern slopes with oak and hornbeam trees on the Muránska planina Mts. fall into the B type (Obuch 2004a). Samples fromthecoldandwetupperpartsoftheplainandfromthe Horehronieregionwithprevailingconiferforestscomprise the type C. Some locations with the E type include food pellets since the year 1978. In four of these locations we could analyze a 20-year time period (Obuch 1997).
The C type from the cold and wet mountain areas is characterized by increased occurrence of mountain species of small ground mammals such as: Sorex alpinus,
Sicista betulina,Microtus tatricus andMicrotus agrestis. This dietary type is dominated by Muscardinus avellanarius (10.2%), whereas the abundance of Apodemus flavicollis(10.1%)isbelowaverageandtheabundanceof C. glareolus 8.7% is close to the Slovak average. Higher humidity is reflected in the presence of Neomys fodiens,
Arvicola amphibius and Rana temporaria (24.2%). The B type from the middle montane zone shows no
characteristic diagnostic species. Some species are commonwithtypesCandE.HighdominanceofA.flavicollis iscommonwithtypeA.However,typeBischaracteristic for its lower proportion ofAves (5.4%).
The D type with markedly high proportion of 13 bat species was identified in 14 sites from carstic regions of Slovakia (Obuch 1998a). This type includes several sub-fossil sites including the finding of Schaefer under the Muráň Mountain in the Belaer Tatras, which was described as a sample from the eagle owl from the 18th century (Schaefer 1974). The explanation of this finding can be found in the paper of Obuch (1992b). Chiroptera comprise 9–90% of the type D samples. Other characteristic prey species include Eliomys quercinus, which is especially abundant in earlier samples but rarely found in the fresh pellets since 1970’.
Besides the most frequently occurring A. flavicollis, thetypeAfromthelowermountainrangesalsoshowsalso
6
Slovak Raptor Journal 2011, 5: 1–120. DOI: 10.2478/v10262-012-0057-8. © Raptor Protection of Slovakia (RPS)
a greater proportion ofAves (8.4%). Several passeriform bird species are quite frequent also in the type F (Coccothraustes coccothraustes and Fringilla coelebs). The dormousespeciesGlisglis(3.3%)ismorecommoninthe lower mountains than in colder regions (Obuch 1998b). The Dryomys nitedula inhabits two different areas: conifer forests in the type C and dry scrubland in the typeA. Recently this species did not appear in several mountain ranges of Central Slovakia, although it could be found as S. aluco prey in the fossil samples from e. g. Veľká Fatra Mts.(Obuch&Darola1980).Crociduraleucodon(1.4%) is the most commonly occurring insectivore.
ThetypeFfromthelowlandfloodplainforests(Obuch 2003)ischaracterizedespeciallybythehighproportionof several frog species (Anura 40%), excluding the species R.temporaria.QuitehighistheproportionofPasseriform birds (12.2%) and lower proportion of mammals (Mammalia, 40.5%), comprising mainly non-forest species such as Microtus arvalis, Apodemus agrarius, Micromys minutus. Floodplain forests of both soft and hard woods are to a great extent altered to monotypic poplar stands, forming mainly forest fragments adjacent to agricultural land. S. aluco hunts frogs in moorlands, especially in bayouswithstillwater.Thistypesharesseveraldiagnostic species of birds and mammals with type G.
TheGtypesamplescomefromdifferentpartsofSlovakia from the plains to the mountain valleys. We found them mainly on the lofts of different buildings and park trees. The first G subtype G1 is distinct for the dominant occurrence of M. arvalis (45.2%), whereas the subtype G2 was characterized by high species diversity of prey withahighproportionofsynanthropicspecies,including among mammals Mus musculus and among birds Passer domesticus.
Spatial and temporal diversity of the diet of Strix aluco in different dietary types in Slovakia
Type A from the lower mountain ranges. In this type (Tab. 2) Mammalia are the dominant food components of S. aluco (77.5%). The most dominant prey species was A. flavicollis (37.5%). The non-forest species M. arvalis was also quite abundant (11.7%). Aves (8.4%) also comprised a common component of the S. aluco diet with a very broad range of passeriform species. The most frequent species included F. coelebs (0.7%), C. coccothraustes (0.7%), Turdus philomelos (0.8%) and
T. merula (0.7%). Amphibia (4.2%) were less frequent than at higher altitudes, including mainly R. temporaria (2.0%) and some other frog species. The most abundant
groupsofEvertebrata(9.9%)includedColeoptera(6.3%), with Limacidae (3.0%) comprising a less important part of the S. aluco diet.
Most ofthematerialofthedietarytypeAwas recorded in the Slovenský kras Mts. and collected from owls which use cave entrances as their diurnal roost place. MostofthemhuntbatsonlyoccasionalywithChiroptera comprising<5%oftheirdiet.Incomparisonwithsamples from other lower mountain ranges in Slovakia, there are four bat species and the forest dormouse (D. nitedula) and non-forest species of small ground mammals (C. leucodon,ApodemusmicropsandM.arvalis).Evertebrata are more frequent.
A rather detailed screening of the S. aluco diet was conducted in the neighbouring Revúcka vrchovina Mts. Someowlsrestclosetotheentrancesofabandonedmines, inhabitedbysummercoloniesofbatspeciesMiniopterus scheibersi and Rhinolophus euryale. Other individuals hunt frogs in shallow marshes (seven species) or some marshlandmammalspecies(MicromysminutusandApodemusagrarius).A.flavicollisisquitedominantsimilarly to Cerová vrchovina Mts. where the dormouse G. glis is also quite abundant. The massif of Drieňová Mt. (616 m a.s.l.)belongstothewarmerpartoftheStrážovskévrchy Mts.withforestsofthe2ndvegetationzoneKropil(1987) found higher proportion of some forest and non-forest species of passeriform birds. Increased abundance could be seen in the species M. arvalis, C. glareolus and Neomys anomalus. Data sample from the Malé Karpaty Mts. comprises a sum of eleven smaller pellet samples.
Slovenský kras Mts. (Table 3). In the Slovenský kras Mts., S. aluco used large entrances to caves and gaps as rest places. Where bats were present in higher numbers, some individuals specialized in hunting them and could beclassifiedastheDdietarytype.Thuswehaveobserved the hunting of Nyctalus noctula in Zbojnícka jaskyňa cave for long periods of time. The sub-recent samples from the caves Ohnište and Erňa at the entrance to the canyon Zádielská dolina valley showed that local owls havehuntedthePipistrelluspipistrellus.InthecaveErňa there is a large winter colony of this bat species (Matis et al. 2002) and in the recent period, there used to be an owl in the cave during the summer, not hunting bats.The dietary type D from the Slovenský kras Mts. comprises five dominant bat species (P. pipistrellus, N. noctula,
Vespertilio murinus, Myotis myotis and M. blythiii). Further seven bat species show greater frequency at some locationswiththedietarytypeA.Thetotalnumberofbat speciesrecordedinthedietofS.alucofromtheSlovenský kras Mts. reached 21.
7
Obuch J: Spatial and chronological diversity of tawny owl (Strix aluco) diet
Tab. 1. Distribution of diagnostic species in the dietary types of Strix aluco in Slovakia
Tab. 1. Zastúpenie diagnostických druhov v typoch potravy Strix aluco na Slovensku
species / dietary type |
|
C |
|
B |
|
E |
|
A |
|
D |
|
G2 |
|
G1 |
|
F |
∑ |
% |
druhy / typ potravy |
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Rana temporaria |
2+ |
2658 |
|
577 |
2- |
439 |
2- |
320 |
1- |
181 |
2- |
100 |
1- |
132 |
6- |
0 |
4407 |
6.47 |
Sicista betulina |
3+ |
170 |
2- |
4 |
2- |
8 |
4- |
0 |
1- |
3 |
1- |
4 |
2- |
0 |
2- |
0 |
189 |
0.28 |
Microtus agrestis |
2+ |
312 |
1- |
34 |
1- |
52 |
2- |
20 |
2- |
5 |
2- |
7 |
1- |
11 |
3- |
0 |
441 |
0.65 |
Microtus tatricus |
1+ |
10 |
|
6 |
|
6 |
1- |
0 |
|
2 |
|
|
|
|
|
|
24 |
0.04 |
Turdus torquatus |
2+ |
36 |
|
7 |
|
13 |
2- |
0 |
|
1 |
|
2 |
|
|
|
|
59 |
0.09 |
Sylvia communis |
1+ |
7 |
|
|
|
1 |
|
3 |
|
1 |
|
1 |
|
2 |
|
|
15 |
0.02 |
Hymenoptera sp. |
1+ |
12 |
|
4 |
|
5 |
|
5 |
|
|
|
3 |
|
3 |
|
|
32 |
0.05 |
Neomys fodiens |
2+ |
77 |
|
12 |
1- |
22 |
1- |
25 |
1- |
5 |
1- |
4 |
1+ |
16 |
|
4 |
165 |
0.24 |
Neomys anomalus |
1+ |
26 |
|
10 |
2- |
8 |
1+ |
46 |
|
7 |
|
7 |
|
9 |
1- |
0 |
113 |
0.17 |
Dryomys nitedula |
1+ |
49 |
1- |
4 |
1- |
16 |
1+ |
46 |
|
10 |
1- |
0 |
1- |
0 |
1- |
0 |
125 |
0.18 |
Arvicola amphibius |
2+ |
263 |
1- |
37 |
1- |
65 |
1- |
47 |
2- |
12 |
1+ |
50 |
|
31 |
1- |
8 |
513 |
0.75 |
Delichon urbicum |
1+ |
67 |
1- |
8 |
1- |
22 |
1- |
17 |
1- |
2 |
1+ |
25 |
|
11 |
|
11 |
163 |
0.24 |
Turdus philomelos |
1+ |
138 |
1+ |
94 |
1- |
80 |
|
118 |
1- |
23 |
1+ |
54 |
|
27 |
|
20 |
554 |
0.81 |
Sorex alpinus |
1+ |
34 |
|
11 |
1+ |
49 |
3- |
0 |
|
5 |
1- |
0 |
|
1 |
1- |
0 |
100 |
0.15 |
Sorex araneus |
1+ |
580 |
1+ |
510 |
1+ |
854 |
1- |
399 |
1- |
106 |
2- |
38 |
2- |
41 |
1- |
58 |
2586 |
3.80 |
Microtus subterraneus |
2+ |
588 |
1+ |
195 |
1- |
135 |
1- |
109 |
|
83 |
2- |
25 |
3- |
7 |
1- |
23 |
1165 |
1.71 |
Talpa europaea |
1+ |
190 |
1+ |
138 |
|
141 |
1- |
84 |
1- |
22 |
1- |
18 |
1- |
13 |
1- |
11 |
617 |
0.91 |
Muscardinus |
1+ |
1126 |
1+ |
529 |
|
1256 |
|
775 |
1- |
153 |
2- |
66 |
2- |
52 |
3- |
18 |
3975 |
5.84 |
avellanarius |
|
|
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Salmo trutta |
1+ |
50 |
1+ |
25 |
|
23 |
2- |
6 |
|
7 |
1- |
1 |
|
4 |
1- |
0 |
116 |
0.17 |
Apodemus flavicollis |
1- |
1113 |
1+ 2252 |
|
4860 |
1+ |
5884 |
1- |
991 |
1- |
510 |
1- |
451 |
2- |
156 |
16217 |
23.82 |
|
Clethrionomys |
|
952 |
1+ |
1055 |
1+ |
2774 |
1- |
1153 |
1- |
362 |
1- |
212 |
2- |
122 |
1- |
127 |
6757 |
9.93 |
glareolus |
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Limacidae sp. |
1- |
582 |
1- |
412 |
2+ |
5668 |
2- |
477 |
2- |
228 |
5- |
11 |
4- |
32 |
6- |
2 |
7412 |
10.89 |
Dendrocopos major |
|
11 |
|
5 |
1- |
4 |
1+ |
19 |
|
3 |
|
4 |
|
1 |
|
5 |
52 |
0.08 |
Lanius collurio |
|
1 |
|
2 |
1- |
0 |
1+ |
10 |
|
|
|
3 |
|
1 |
|
1 |
18 |
0.03 |
Sitta europaea |
1- |
5 |
|
7 |
1- |
12 |
1+ |
46 |
|
7 |
|
6 |
|
4 |
|
8 |
95 |
0.14 |
Emberiza citrinella |
1- |
3 |
|
5 |
2- |
2 |
1+ |
27 |
|
6 |
|
8 |
|
2 |
|
5 |
58 |
0.09 |
Garrulus glandarius |
|
12 |
|
6 |
1- |
6 |
1+ |
42 |
|
3 |
|
5 |
|
2 |
|
|
76 |
0.11 |
Lepus europaeus |
|
1 |
|
2 |
|
1 |
1+ |
9 |
|
1 |
|
|
|
1 |
|
2 |
17 |
0.02 |
Miniopterus |
|
|
|
|
1- |
0 |
1+ |
13 |
|
2 |
|
|
|
|
|
|
15 |
0.02 |
schreibersii |
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Rhinolophus euryale |
1- |
0 |
|
|
1- |
1 |
2+ |
31 |
|
|
|
|
|
|
|
|
32 |
0.05 |
Coleoptera sp. |
|
459 |
|
284 |
2- |
243 |
1+ |
989 |
2- |
56 |
1- |
94 |
1- |
58 |
1+ |
199 |
2382 |
3.50 |
Fringilla coelebs |
|
63 |
1- |
37 |
2- |
46 |
1+ |
150 |
|
27 |
1+ |
56 |
|
29 |
1+ |
52 |
460 |
0.68 |
Coccothraustes |
2- |
7 |
1- |
7 |
3- |
8 |
1+ |
114 |
|
11 |
1+ |
26 |
1- |
3 |
1+ |
27 |
203 |
0.30 |
coccothraustes |
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Parus major |
2- |
16 |
2- |
8 |
2- |
24 |
1+ |
101 |
|
16 |
2+ |
99 |
|
15 |
|
14 |
293 |
0.43 |
Erithacus rubecula |
1- |
33 |
|
25 |
1- |
40 |
1+ |
91 |
1- |
10 |
1+ |
38 |
|
15 |
|
18 |
270 |
0.40 |
Orthoptera sp. |
1- |
6 |
|
14 |
1- |
17 |
1+ |
72 |
1- |
1 |
1+ |
18 |
1- |
1 |
|
3 |
132 |
0.19 |
Crocidura leucodon |
4- |
1 |
2- |
6 |
3- |
10 |
2+ |
219 |
|
36 |
2+ |
67 |
|
12 |
1- |
5 |
356 |
0.52 |
Glis glis |
1- |
158 |
|
182 |
|
325 |
1+ |
519 |
1+ |
147 |
3- |
13 |
1- |
39 |
4- |
0 |
1383 |
2.03 |
Myotis mystacinus |
1- |
6 |
1- |
1 |
1- |
8 |
1+ |
35 |
2+ |
29 |
|
4 |
|
4 |
|
1 |
88 |
0.13 |
Pipistrellus pipistrellus |
6- |
0 |
4- |
3 |
2- |
60 |
4- |
12 |
4+ |
916 |
4- |
0 |
4- |
0 |
3- |
2 |
993 |
1.46 |
Barbastella |
2- |
3 |
2- |
2 |
|
16 |
1- |
17 |
3+ |
121 |
1- |
1 |
1- |
0 |
1- |
0 |
160 |
0.24 |
barbastellus |
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Vespertilio murinus |
1- |
23 |
1- |
10 |
2- |
17 |
3- |
5 |
3+ |
208 |
1- |
4 |
2- |
0 |
2- |
0 |
267 |
0.39 |
Myotis myotis |
2- |
25 |
1- |
17 |
1- |
52 |
1- |
39 |
3+ |
277 |
|
25 |
2- |
2 |
3- |
0 |
437 |
0.64 |
8
Slovak Raptor Journal 2011, 5: 1–120. DOI: 10.2478/v10262-012-0057-8.
© Raptor Protection of Slovakia (RPS)
Tab. 1. continuation / pokračovanie
species / dietary type |
|
C |
|
B |
|
E |
|
A |
|
D |
|
G2 |
|
G1 |
|
F |
∑ |
% |
druhy / typ potravy |
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Myotis brandtii |
|
3 |
|
1 |
2- |
0 |
|
8 |
2+ |
24 |
|
|
|
|
|
|
36 |
0.05 |
Eptesicus serotinus |
1- |
9 |
|
14 |
1- |
17 |
|
28 |
2+ |
28 |
|
2 |
|
1 |
1- |
0 |
99 |
0.15 |
Plecotus auritus |
|
4 |
|
2 |
|
17 |
|
6 |
1+ |
13 |
|
1 |
|
1 |
|
|
44 |
0.06 |
Myotis blythii |
|
2 |
|
|
|
8 |
|
8 |
1+ |
12 |
|
4 |
|
|
|
|
34 |
0.05 |
Myotis bechsteini |
1- |
1 |
|
4 |
|
12 |
|
14 |
1+ |
14 |
|
|
|
|
|
|
45 |
0.07 |
Rhinolophus |
|
|
|
|
|
|
|
7 |
1+ |
7 |
|
|
|
|
|
|
14 |
0.02 |
ferrumequinum |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Eliomys quercinus |
|
11 |
|
4 |
|
6 |
|
9 |
1+ |
10 |
|
|
|
|
|
|
40 |
0.06 |
Lacerta muralis |
|
|
|
|
|
1 |
|
3 |
1+ |
5 |
|
|
|
|
|
|
9 |
0.01 |
Motacilla cinerea |
|
3 |
|
3 |
|
1 |
|
1 |
1+ |
9 |
|
1 |
|
|
|
|
18 |
0.03 |
Nyctalus noctula |
3- |
3 |
2- |
5 |
2- |
31 |
4- |
4 |
3+ |
246 |
1+ |
44 |
2- |
3 |
1- |
9 |
345 |
0.51 |
Rhinolophus |
1- |
3 |
1- |
1 |
|
10 |
|
9 |
1+ |
9 |
2+ |
15 |
|
1 |
|
|
48 |
0.07 |
hipposideros |
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Myotis emarginatus |
1- |
0 |
|
|
2- |
0 |
|
10 |
|
3 |
2+ |
16 |
|
1 |
|
1 |
31 |
0.05 |
Columba livia |
|
3 |
|
2 |
|
4 |
1- |
0 |
|
|
2+ |
15 |
|
|
|
|
24 |
0.04 |
domestica |
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Streptopelia decaocto |
|
1 |
|
2 |
1- |
0 |
1- |
0 |
|
|
2+ |
14 |
|
2 |
|
|
19 |
0.03 |
Apus apus |
|
1 |
|
1 |
|
1 |
|
|
|
|
2+ |
11 |
|
|
|
|
14 |
0.02 |
Phoenicurus ochruros |
|
3 |
|
1 |
1- |
0 |
|
2 |
|
2 |
2+ |
13 |
|
3 |
|
1 |
25 |
0.04 |
Turdus merula |
|
74 |
|
39 |
1- |
45 |
|
112 |
|
24 |
2+ |
75 |
|
21 |
|
13 |
403 |
0.59 |
Muscicapa striata |
|
1 |
|
|
|
6 |
|
1 |
|
|
1+ |
6 |
|
|
|
|
14 |
0.02 |
Carduelis spinus |
|
4 |
|
|
1- |
3 |
|
7 |
|
2 |
1+ |
7 |
|
1 |
|
5 |
29 |
0.04 |
Phylloscopus collybita |
|
|
|
|
|
1 |
|
1 |
|
2 |
1+ |
7 |
|
2 |
|
3 |
16 |
0.02 |
Phylloscopus trochilus |
|
|
|
|
|
|
|
2 |
|
1 |
1+ |
6 |
|
2 |
|
|
11 |
0.02 |
Motacilla alba |
|
|
|
|
|
1 |
|
3 |
|
|
1+ |
9 |
|
4 |
|
|
17 |
0.02 |
Carduelis chloris |
1- |
4 |
1- |
0 |
3- |
0 |
1- |
6 |
|
1 |
3+ |
43 |
|
3 |
1+ |
8 |
65 |
0.10 |
Sylvia atricapilla |
1- |
4 |
1- |
4 |
1- |
7 |
|
20 |
1- |
1 |
2+ |
29 |
|
7 |
1+ |
14 |
86 |
0.13 |
Passer montanus |
2- |
4 |
2- |
1 |
3- |
0 |
|
26 |
1- |
2 |
2+ |
45 |
|
7 |
2+ |
34 |
119 |
0.17 |
Hirundo rustica |
|
5 |
|
7 |
1- |
7 |
|
7 |
|
2 |
1+ |
8 |
|
4 |
1+ |
7 |
47 |
0.07 |
Carduelis cannabina |
|
3 |
1- |
0 |
1- |
2 |
|
10 |
|
1 |
1+ |
8 |
|
6 |
1+ |
9 |
39 |
0.06 |
Serinus serinus |
1- |
0 |
|
1 |
2- |
0 |
|
3 |
|
1 |
2+ |
14 |
1+ |
6 |
1+ |
7 |
32 |
0.05 |
Carduelis carduelis |
1- |
4 |
2- |
0 |
2- |
2 |
1- |
11 |
1- |
0 |
2+ |
30 |
1+ |
11 |
2+ |
19 |
77 |
0.11 |
Rattus norvegicus |
1- |
8 |
1- |
3 |
3- |
2 |
1- |
19 |
1- |
2 |
3+ |
64 |
1+ |
19 |
1+ |
15 |
132 |
0.19 |
Mus cf. musculus |
1- |
32 |
2- |
10 |
6- |
0 |
2- |
41 |
1- |
28 |
3+ |
308 |
1+ |
40 |
1+ |
43 |
502 |
0.74 |
Micromys minutus |
2- |
18 |
2- |
5 |
4- |
3 |
|
75 |
1- |
12 |
2+ |
97 |
2+ |
51 |
3+ |
99 |
360 |
0.53 |
Apodemus microps |
2- |
1 |
2- |
0 |
3- |
0 |
|
21 |
|
5 |
2+ |
33 |
2+ |
28 |
2+ |
22 |
110 |
0.16 |
Apodemus agrarius |
3- |
13 |
4- |
0 |
6- |
0 |
1+ |
221 |
|
33 |
1+ |
69 |
1+ |
62 |
3+ |
160 |
558 |
0.82 |
Microtus arvalis |
1- |
512 |
1- |
527 |
3- |
305 |
1+ |
1841 |
|
487 |
1+ |
684 |
3+ |
1466 |
1+ |
402 |
6224 |
9.14 |
Cyanistes caeruleus |
1- |
9 |
2- |
1 |
1+ |
19 |
1+ |
50 |
|
6 |
2+ |
28 |
1+ |
12 |
|
8 |
133 |
0.20 |
Crocidura suaveolens |
4- |
1 |
3- |
3 |
4- |
2 |
|
86 |
1+ |
42 |
3+ |
144 |
1+ |
31 |
1- |
5 |
314 |
0.46 |
Apodemus sylvaticus |
2- |
15 |
1- |
13 |
4- |
2 |
|
75 |
|
26 |
3+ |
130 |
1+ |
44 |
|
10 |
315 |
0.46 |
Turdus pilaris |
|
35 |
1- |
6 |
3- |
3 |
2- |
12 |
1- |
2 |
3+ |
72 |
2+ |
31 |
1- |
2 |
163 |
0.24 |
Passer domesticus |
3- |
4 |
4- |
1 |
5- |
2 |
1- |
57 |
3- |
0 |
3+ |
237 |
2+ |
77 |
|
19 |
397 |
0.58 |
Sturnus vulgaris |
|
5 |
|
4 |
2- |
0 |
|
11 |
|
2 |
2+ |
24 |
1+ |
9 |
|
3 |
58 |
0.09 |
Sorex minutus |
|
122 |
1+ |
98 |
1- |
126 |
|
157 |
1- |
33 |
1- |
20 |
1+ |
44 |
1+ |
51 |
651 |
0.96 |
Rana cf. esculenta |
5- |
0 |
4- |
2 |
6- |
0 |
1+ |
255 |
2- |
13 |
1- |
21 |
2- |
4 |
3+ |
232 |
527 |
0.77 |
Rana arvalis |
4- |
0 |
3- |
0 |
5- |
0 |
3- |
4 |
3- |
1 |
2- |
1 |
1- |
6 |
4+ |
268 |
280 |
0.41 |
Rana ridibunda |
3- |
0 |
3- |
0 |
4- |
0 |
2- |
5 |
2- |
0 |
2- |
0 |
1- |
1 |
4+ |
154 |
160 |
0.24 |
Pelobates fuscus |
5- |
0 |
4- |
2 |
6- |
0 |
1- |
55 |
3- |
3 |
1- |
10 |
|
19 |
4+ |
435 |
524 |
0.77 |
9