

Морфология растений / Шамров И.И. Семязачаток цветковых растений строение, функции, происхождение
.pdf70 |
Шамров И.И. Семязачаток цветковых растений: строение, функции, происхождение |
|
|
(Chenopodiaceae — Bruun, Olesen, 1989) клетки париетальной ткани также вовле- чены в процесс продвижения пыльцевой трубки к зародышевому мешку. Перед оплодотворением ее клеточные оболочки становятся интенсивно ШИК-положитель- ными, а в цитоплазме секретируются полисахариды, которые часто выходят в микропилярный канал.
Âслучае длительного существования париетальной ткани, она, возможно, уча- ствует в апикальном транспорте веществ из окружающих структур в развивающийся зародыш. В ее клетках обнаруживаются различные метаболиты в виде белков и декстринов. У Luzula pedemontana (Шамров, Анисимова, 1993в) на стадии трехклеточного проэмбрио в тангентальных стенках клеток периферических рядов париетальной ткани (контактирующих непосредственно с клетками латеральной области нуцеллуса) откладываются полисахариды, возможно, каллоза. Тем самым создается канализированное поступление веществ в зародыш из окружающих тканей через клетки центральной части париетальной ткани.
Основную часть нуцеллуса крассинуцеллятного семязачатка составляет паренхима нуцеллуса, которая располагается преимущественно в латеральной области. Число слоев паренхимы нуцеллуса варьирует от одного (некоторые виды Alliaceae — Stenar, 1933; Cave, 1948; Berg, 1978), 2 (Limnocharitaceae — Johri, 1938; Liliaceae — Петрова, 1977; Шамров, 1999а), 3–4 (Poaceae — Батыгина, 1974), 6–7 (Costaceae — Grootjen, Bouman, 1981) äî 13–15 (Ceratophyllaceae — Шамров, 1997и).
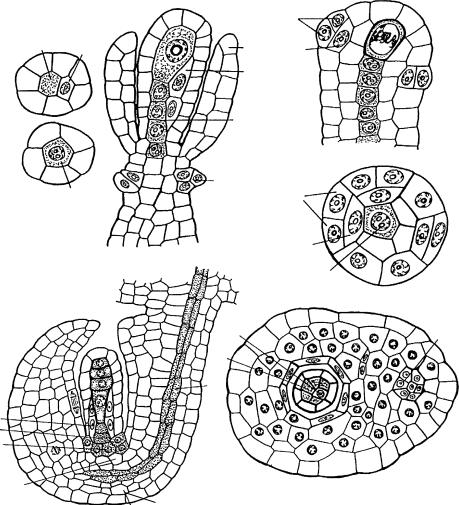
Âсемязачатках с однослойной латеральной областью нуцеллуса обнаружены различия в ее строении. У многих таксонов она представлена слоем, имеющим на поперечном срезе вид кольца. У представителей ряда семейств слой становится асимметричным (число клеток, образующих слой, на поперечном срезе может быть различным), что вызывает смещение осевого ряда клеток под археспориальной клеткой из центрального положения в латеральное (Listera ovata, Orchidaceae — Шамров, Никитичева, 1992; Шамров, 2001; Azorina vidalii, Campanulaceae — Шамров, Жинкина, 1994; Cytinus ruber, Rafflesiaceae — по препаратам, любезно предоставленным Э.С. Терехиным и Г.М. Анисимовой) (рис. 22). Подобное, вероятно, имеет место в семействах Dipsacaceae (Камелина, 1980), Scrophulariaceae (Никитичева, 1987б) и Araceae (Коробова, Жинкина, 1990).
При исследовании Ceratophyllum demersum нами была установлена гетерогенность клеток и наличие структурной дифференциации на внутренние и наружные слои в латеральной области нуцеллуса. Эта дифференциация наблюдается с самого раннего развития и обусловлена тем, что каждая часть латеральной области нуцеллуса образуется за счет самостоятельных инициалей. В сформированном семяза- чатке 5–6-слойная наружная часть образована изодиаметрическими или прямоугольными клетками. Внутренняя часть состоит из клеток, сильно вытянутых вдоль оси нуцеллуса. Такая же особенность, вероятно, характерна для Nelumbo nucifera (Батыгина и др., 1983). У ряда таксонов неодинаковое строение латеральной области нуцеллуса выявляется только в сформированном семязачатке (Gossypium
Глава 2. Стр т рная дифференциация семязачат а |
71 |
|
|
hirsutum, Malvaceae — Jensen, 1965; Alcea rosea, Malvaceae — Плиско, 1980; Oenothera biennis, Onagraceae — Noher de Halac, 1979; Agave parryi, Agavaceae
— Tilton, Mogensen, 1979; Spinacia oleracea, Chenopodiaceae — Wilms, 1980).
á î í ë î í
1
2
á î í
è â è
â è è ã
ë î í |
|
á î í |
|
|
4 |
í è |
|
è â è |
|
3 |
|
è ã |
|
î ð ê |
5 |
á î í |
ïð ê |
ë î í |
|
ïð ê
ëî í
áî í
ã
â î õ |
6 |
7 |
|
||
|
|
|
|
í î õ |
|
Рис. 22. Асимметричное строение субэпидермального слоя нуцеллуса.
1–3 — Cytinus ruber (по: Шамров, 1995); 4, 5 — Listera ovata (по: Шамров, 2001); 6, 7 — Azorina vidalii (по: Шамров, Жинкина, 1994); 1, 2, 5, 7 — поперечные и 3, 4, 6 — продольные срезы; á î í — базальная область нуцеллуса, â è — внутренний интегумент, â î õ — внутренняя область халазы, è â è — инициали внутреннего интегумента, è ã — инициали гипостазы, ã — гипостаза, ë î í — латеральная область нуцеллуса, í è — наружный интегумент, í î õ — наружняя область халазы, î ð ê — осевой ряд клеток, ïð ê — прокамбиальные клетки.
72 |
Шамров И.И. Семязачаток цветковых растений: строение, функции, происхождение |
|
|
Клетки паренхимы нуцеллуса в начальный период развития семязачатка имеют правильную форму; вакуоли мелкие, цитоплазма насыщена свободными рибосомами, хорошо развит ЭР и аппарат Гольджи. Стенки клеток пронизаны много- численными плазмодесмами (Noher de Halac, 1979; Tilton, Mogensen, 1979; Плиско, 1980; Wilms, 1980; Belyaeva, 1983; Fougиre-Rifot, 1988; Полюшкина, 1993). В процессе развития происходит специализация клеток и в них накапливаются различные вещества.
Еще до оплодотворения клетки паренхимы нуцеллуса могут подвергаться дегенерации. Этот процесс охватывает сначала внутренние клетки, примыкающие к зародышевому мешку. Как показали ультраструктурные исследования, дегенерация клеток нуцеллуса происходит по типу локального автолиза, о чем свидетельствует наличие в них большого количества хорошо развитых лизосом и аккумуляция кислой фосфатазы на плазмалемме (Schulz, Jensen, 1966, 1971; D’AlascioDeschamps, 1973; Norstog, 1974; Matile, 1975; Fougиre-Rifot, 1988). На примере Aquilegia vulgaris показано, что лизис клеток нуцеллуса происходит в два этапа: сначала дегенерирует цитоплазма под действием внутривакуолярных гидролаз, а затем клеточные стенки — под действием экзогидролаз (Fougиre-Rifot, 1988). Некоторые авторы (Чочиа, 1989) предполагают, что лизис нуцеллуса может происходить иначе — путем коагуляционного некроза. Содержимое клеток сжимается, разрушается пространственная организация органелл, возрастает плотность гиалоплазмы и нуклеоплазмы, вакуоль распадается на мелкие пузырьки, в пластидах появляются пластоглобулы. Клетки уплощаются и постепенно приобретают вид гомогенной пленки, наслаивающейся на оболочку зародышевого мешка. Существует также точка зрения, что дегенерация нуцеллуса представляет собой процесс запрограммированной гибели клеток: сначала наблюдается фрагментация ДНК в ядрах, затем происходит деструкция ядерной оболочки, органелл цитозоля и плазмалеммы, при этом дольше всего сохраняются интактными и возможно функциональными элементы аппарата Гольджи (Dominguez et al., 2001).
Нуцеллус в крассинуцеллятном семязачатке может разрушаться в процессе развития (Annonaceae, Asparagaceae, Magnoliaceae, Myristicaceae, Trilliaceae,
Winteraceae) либо полностью или частично сохраняться, трансформируясь в перисперм (ãðå÷. peri — вокруг, около, sperma — семя) — запасную ткань семени. Термин был введен Juel (1907), хотя сама ткань была описана первоначально Brown (1826) под названием «первичный белок» (лат. albumen primarium). Нали- чие перисперма в семени большинство авторов рассматривает как примитивный признак.
Генезис нуцеллуса, преобразующегося после оплодотворения в перисперм, изучен недостаточно. Обычно констатируют лишь факт наличия в семени перисперма, возникающего из клеток так называемого «центрального тела» амфитропного и кампилотропного семязачатков — остатка нуцеллуса со стороны халазы, над которым располагается изогнутый в виде дуги или подковы зародыш (Gibbs, 1907; Kajale, 1954; Bocquet, 1959; Турсунов, 1986) (рис. 23, 1). Исследование нуцеллуса в
Глава 2. Стр т рная дифференциация семязачат а |
73 |
|
|
динамике показало, что у Nuphar lutea (Nymphaeaceae) не во всех сохраняющихся клетках откладывается запасной крахмал, что предшествует образованию перисперма. Нуцеллус является гетерогенным по особенностям развития и строения отдельных зон. В нуцеллусе дифференцируются нуцеллярный колпачок, париетальная ткань, постамент и подиум. В процессе развития семени постамент и клетки микропилярной зоны нуцеллуса вокруг развивающегося эндосперма, в том числе и клетки париетальной ткани, разрушаются (Шамров, 1997в, 1998а). Свободными от крахмала остаются подиум и идущие от него до зародыша 2–3 слоя узких субэпидермальных клеток (своеобразная «периферическая проводящая система») (Shamrov, 2000a) (рис. 23, 2–4). Ó Hablitzia tamnoides (Chenopodiaceae) в крассинуцеллятном семязачатке дифференцируются двухслойный нуцеллярный колпачок и 4–6-слойная париетальная ткань, клетки которых в процессе развития постепенно разрушаются. Асимметричная латеральная область нуцеллуса, более массивная с абаксиальной стороны семязачатка, сохраняется в центре семени. Она трансформируется в перисперм, в клетках которого накапливается крахмал (Камелина, 2001).
При анализе данных о наличии перисперма в семени выявляются определенные корреляции между степенью развития перисперма, с одной стороны, и эндосперма и зародыша, с другой стороны. Исходя из этого, выделены 4 группы семян с периспермом (Шамров, 1997в). Первая группа — перисперм составляет основную часть зрелого семени, эндосперма сравнительно мало, зародыш небольших размеров (Barclayaceae, Cabombaceae, Cannaceae, Costaceae, Marantaceae, Nymphaeaceae, Zingiberaceae, некоторые Saururaceae, Piperaceae). Вторая группа — перисперм составляет половину объема зрелого семени и окружает крупный изогнутый зародыш только со стороны халазы, эндосперма относительно мало (Amaranthaceae, Caryophyllaceae, Chenopodiaceae (подсем. Chenopodioideae), Basellaceae, Nyctaginaceae, Phytolaccaceae, Portulacaceae, Stegnospermataceae).
Третья группа — перисперм в виде тонкого слоя, зародыш небольших размеров, эндосперм занимает основную часть зрелого семени (Heliconiaceae, Musaceae). Четвертая группа — перисперм имеется только вокруг верхушек семядолей, зародыш крупный, занимает почти весь объем зрелого семени, и в нем накапливаются запасные вещества, эндосперм в виде тонкого слоя вокруг зародышевого корня (Ñhenopodiaceae — подсем. Salsoloideae).
Своеобразное преобразование халазальной зоны нуцеллуса еще во время развития семязачатка описано в сем. Podostemaceae. Уже на стадии двуядерного зародышевого мешка происходит дезинтеграция клеток этой зоны и постепенно возникает многоядерный протопласт. Предполагают, что он выполняет питательную функцию, замещая эндосперм. Это образование предложили называть «нуцеллярным плазмодием» (Mukkada, 1962; Jдger-Zьrn, 1967).
Мы уже отмечали, что нуцеллус, особенно в крассинуцеллятном семязачатке, имеет дифференцированное строение. В его микропилярной зоне образуются нуцеллярный колпачок, археспорий, париетальная ткань. Еще более структурированными являются средняя и халазальная зоны нуцеллуса, в которых формируются

74 |
Шамров И.И. Семязачаток цветковых растений: строение, функции, происхождение |
|
|
çàð |
ñ ê |
|
ïåð |
||
|
1
ýíä
ýíä
ó ê í
ïåð
4 |
ã ïä |
Рис. 23. Формирование перисперма.
ïñ
ïä
ïð ï
2
ïñ
ïä
ã
3
1 — зрелое семя Stellaria media (ïî: Gibbs, 1907); 2–4 — преобразование нуцеллуса в перисперм у Nuphar lutea (по: Шамров, 1998а); ã — гипостаза, çàð — зародыш, ïä — подиум, ïåð — перисперм, ïð ï — проводящий пучок, ïñ — постамент, ñ ê — семенная кожура, ó ê í — узкие клетки нуцеллуса, ýíä — эндосперм.
Глава 2. Стр т рная дифференциация семязачат а |
75 |
|
|
постамент и подиум. Эти структуры, а также гипостаза, по мнению ряда исследователей, принципиально не различаются, а представляют лишь модификации одной и той же структуры (Johansen, 1928; Dahlgren, 1940; Maheshwari, 1950; Савченко, 1973; Поддубная-Арнольди, 1976; Szujkу-Lacza, 1978; Tilton, Mogensen, 1979; Rudall, 1997).
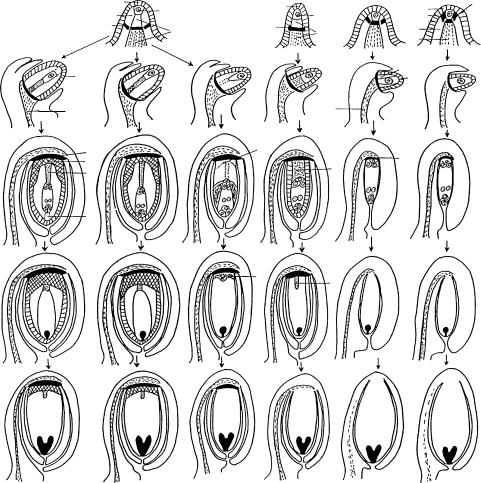
Изучение рассматриваемых структур с самых ранних стадий показало, что они действительно развиваются как единое образование. Их дифференциация связана с активностью клеток, прежде всего базальной, латеральной и переходной зон примордия семязачатка. Как уже было указано выше, переходная зона примордия семязачатка образует собственно гипостазу. Имеющиеся данные свидетельствуют о том, что базальная зона, представленная продольными рядами таблитчатых или удлиненных клеток, связана с проводящей системой плаценты и завязи. В зависимости от типа формирующегося семязачатка и характера трансформации этой зоны могут возникать постамент, подиум и проводящий пучок (рис. 24). В крассинуцеллятных семязачатках постамент и подиум (центральная часть) возникают из клеток проксимального (по отношению к археспориальной клетке) района, а проводящий пучок — из клеток дистального района меристемы базальной зоны, при этом подиум дифференцируется непосредственно под постаментом (Ceratophyllum demersum, Ceratophyllaceae; Ribes aureum, Grossulariaceae; Nuphar lutea, Nymphaeaceae — Шамров, 1997и, 1998а; Ungnadia speciosa, Sapindaceae — Anisimova, Shamrov, 1997). Латеральные части подиума образуются из клеток меристемы латеральной зоны примордия семязачатка. В тенуинуцеллятных семязачатках (Gentianaceae — Шамров, 1990б) формируется только проводящий пучок.
Пути дифференциации нуцеллуса в медионуцеллятных семязачатках оказались более разнообразными (Шамров, 2002б), чем считалось ранее (Батыгина, Шамров, 1994а). Если семязачатки довольно массивные и близки по строению к крассинуцеллятным семязачаткам (как правило, синдермальная вариация), в них могут возникать подиум и постамент (Zea mays, Poaceae — Воронова и др., 2002; возможно, Muilla maritima — Berg, Maze, 1966 è âèäû Allium, Alliaceae — Ashurmetov, Yengalycheva, 1997; Ashurmetov et al., 2001; ряд представителей сем. Ranunculaceae — Bouman, Calis, 1977; Бутузова, 1999). Если организация медионуцеллятных семязачатков сходна с таковой тенуинуцеллятных, то в них дифференцируется только постамент (Gymnadenia conopsea, Orchidaceae — Шамров, Никитичева, 1992) либо вся халазальная зона нуцеллуса трансформируется в поста- менто-подиум (Azorina vidalii, Campanulaceae — Шамров, Жинкина, 1994; Gagea stipitata, Liliaceae; Capsella bursa-pastoris, Brassicaceae — Шамров, 1999а, 2002а). Время инициации структур, продолжительность существования и особенности их строения варьируют у разных таксонов. Эти параметры тесно связаны с особенностями генезиса других сопряженно развивающихся структур семязачатка и семени, что в совокупности определяет специфику мегаспорогенеза, развития зародышевого мешка, зародыша и эндосперма.
76 |
Шамров И.И. Семязачаток цветковых растений: строение, функции, происхождение |
|
|
ï ç |
à ç |
ï ç |
|
ï ç |
ïðõ ç |
|
|
|
|||||
ïðõëç |
ç |
ïðõ ç |
à ç |
ëàçç |
|
|
á ç |
|
|||||
|
|
|
á ç |
á ç |
|
|
ý í |
|
|
|
|
ë î í |
|
|
|
|
|
ý í |
|
|
ïð ï |
|
|
|
ïð ï |
|
|
ã |
|
|
ïïä |
|
ã |
|
ïä |
|
|
|
ïñ |
|
|
ïñ |
|
|
|
|
|
|
ï ò
ïïä
ïñ
Рис. 24. Схема, иллюстрирующая преобразование базальной и латеральной зон примордия семязачатка в структуры халазальной части семязачатка (по: Шамров, 2002б).
Крайний левый столбец — крассинуцеллятный семязачаток, крайний правый столбец — тенуинуцеллятный семязачаток, четыре средних столбца — медионуцеллятные семязачатки; a ç — апикальная зона, á ç — базальная зона, ã — гипостаза, ë ç — латеральная зона, ë î í — латеральная область нуцеллуса, ïä — подиум, ï ç — периферическая зона, ïïä — постаменто-по- диум, ïð ï — проводящий пучок, ïðõ ç — переходная зона, ïñ — постамент, ï ò — париетальная ткань, ý í — эпидерма нуцелуса.
Глава 2. Стр т рная дифференциация семязачат а |
77 |
|
|
Таким образом, гипостаза, постамент и подиум различаются особенностями генезиса, хотя могут выполнять, как будет показано в дальнейшем, сходные функции (Шамров, Винтер, 1991; Shamrov, 1992; Батыгина, Шамров, 1994а, б; Шамров, 1995, 1999б, 2002б; Batygina, Shamrov, 1999, 2002a, b).
Осевая часть халазальной зоны нуцеллуса семязачатка представлена постаментом (ëàò. postament — колонка). Синонимы: колонка, пьедестал, подставка для антипод. Постамент располагается в виде колонки под спорогенными или гаметофитными структурами.
Колонка клеток в основании развивающегося зародышевого мешка была впервые обнаружена у Butomus umbellatus (Ward, 1880). Однако термин «постамент» первым употребил Westermaier (1890). Под постаментом понималась колонка из удлиненных клеток под антиподальным концом зародышевого мешка, при- чем они отличались от окружающих клеток более густой цитоплазмой и более интенсивно окрашивающимися оболочками. Считалось, что основная функция постамента — трофическая.
K. Dahlgren (1939, 1940) расширил объем термина и полагал, что постамент присутствует как на стадии зародышевого мешка, так и на различных стадиях развития семени. Им было высказано предположение о двух способах возникновения постамента: 1) «врастание» ткани под антиподами в полость зародышевого мешка, 2) преобразование гипостазы, располагающейся в виде колонки под зародышевым мешком и представленной клетками с утолщенными оболочками. Различные авторы поддерживают либо первое (Комар, 1965; Савченко, 1973), либо второе (Поддубная-Арнольди, 1976; Фрейберг, Камелина, 1981) положение.
Согласно нашим представлениям, в постамент трансформируются клетки продольных рядов меристемы базальной зоны примордия семязачатка, располагающиеся под археспориальной клеткой. Постамент представляет собой осевую часть халазальной зоны нуцеллуса семязачатка (Shamrov, 1992, 2002k; Шамров, 1994л). Сходная точка зрения на формирование постамента высказывалась и ранее (Mьcke, 1908). Как «своеобразная гипостаза» он был описан для злаков (Батыгина, 1974, 1987).
Постамент образован удлиненными или таблитчатыми клетками в виде рядов, доходящих до гипостазы, часто напоминающих по строению прокамбиальные клетки (Mьcke, 1908; Joshi, Venkateswarlu, 1935; Смирнов, 1982; Камелина, Проскурина, 1987; Алимова, 1990б; Шамров, Никитичева, 1992; Шамров, 1995, 1997з, и, 1998; Shamrov, 1998) (рис. 25, 2, 3, 5, 7; 26, 4). Клетки постамента, например, у Quercus gambelii (Fagaceae), связаны между собой немногочисленными плазмодесмами и содержат ядра удлиненной формы, а также большое количество митохондрий, свободных рибосом и хорошо развитый ЭР. По периферии этих клеток плазмалемма формирует инвагинации различных размеров (Mogensen, 1973). Оболочки клеток постамента могут иметь различное строение. Так, у Glycine max (Fabaceae) продольные стенки ровные, уплощенные, с плазмодесмами, а поперечные — сильно извилистые, с множеством протуберанцев (Полюшкина, 1993).
78 |
Шамров И.И. Семязачаток цветковых растений: строение, функции, происхождение |
|
|
Продолжительность существования постамента варьирует у разных растений и скоррелирована с особенностями развития и функционирования других структур нуцеллуса. Чаще всего он полностью разрушается вскоре после оплодотворения. У некоторых таксонов (Taccaceae — Håkansson, 1921; Poaceae — Батыгина, 1974; Asparagaceae — Сатарова, 1990) разрушение клеток колонки происходит еще до оплодотворения и сопровождается постепенным их замещением растущим зародышевым мешком, в результате чего обеспечивается более тесный контакт с проводящей системой семязачатка и завязи. У представителей сем. Calycanthaceae (Камелина, 1981) на месте разрушающихся клеток осевой части халазальной зоны нуцеллуса формируется лизигенная полость над гипостазой. При длительном существовании постамента происходит постепенное разрушение латеральных слоев, а затем и эпидермального слоя в халазальной зоне нуцеллуса. У одних таксонов (Orchidaceae, Fagaceae, Nymphaeaceae, Ranunculaceae) постамент, представленный тонкостенными клетками, разрушается на средних стадиях эмбриогенеза, у других (Araceae, Costaceae, Geissolomataceae, Grossulariaceae, Zingiberaceae) он состоит из одревесневших клеток и сохраняется в зрелом семени (рис. 25, 4).
Исходя из строения клеток и расположения их в виде рядов, а также накопления в них белков, крахмала, декстринов и других веществ, можно полагать, что основная функция постамента связана с транспортом метаболитов, поступающих из гипостазы в спорогенные, а позже — в гаметофитные структуры. По мнению Eames (1961), в семязачатках с массивным нуцеллусом формируется «центральный пучок» (постамент, согласно нашему представлению), который может служить доказательством существования сосудистого снабжения нуцеллуса. Если постамент сохраняется в зрелом семени, он выполняет и защитную функцию.
Другой специализированной структурой халазальной зоны нуцеллуса является подиум (ãðå÷. podion — основание). Термин ввел K. Dahlgren (1939, 1940) для обозначения долго сохраняющегося в этой зоне остатка нуцеллуса чашевидной формы. Согласно этому автору, у Zostera marina халазальная зона нуцеллуса в сформированном семязачатке имеет вид колонки. После оплодотворения происходит постепенное ее разрушение, при этом верхняя часть (более узкая) становится, по мнению автора, постаментом, а более массивная нижняя часть — подиумом. Клетки последнего приобретают утолщенные оболочки и сохраняются в зрелом семени, что дало основание ряду последующих авторов (Maheshwari, 1950; Савич, 1968; Савченко, 1973) трактовать эту структуру как гипостазу.
Согласно нашим представлениям, подиум, как и постамент, — это не возрастные состояния нуцеллуса в халальной зоне, а специализированные структуры, которые создаются в нуцеллусе для выполнения самых необходимых функций, связанных с переносом метаболитов в зародышевый мешок. При трактовке подиума, которую предложил Dahlgren (1939, 1940), упрощается целесообразность возникновения этой структуры в нуцеллусе и семязачатке в целом. По нашему мнению, подиум формируется в красси- и медионуцеллятных семязачатках с многослойной латеральной областью нуцеллуса, которая продолжительное время функционирует
Глава 2. Стр т рная дифференциация семязачат а |
79 |
|
|
после оплодотворения (Shamrov, 1992, 2002m; Шамров, 1994к). Подиум дифференцируется в форме бокала в халазальной зоне нуцеллуса на ранних стадиях развития семязачатка за счет периклинальных делений клеток, как правило, субэпидермы, реже и эпидермы (рис. 25, 1) (Trapaceae — Titova et al., 1997; Zea mays, Poaceae
—Воронова и др., 2002) латеральной области нуцеллуса (латеральная часть подиума) и антиклинальных делений клеток нижних слоев базальной области нуцеллуса (центральная часть подиума). В сформированном состоянии его верхняя граница находится на уровне антипод зародышевого мешка (Shamrov, 1992, 1998; Шамров, Анисимова, 1993а–в, Шамров, 1995, 1997з, и, 1998а, 1999б).
Как показал анализ литературных данных, в семязачатке ряда растений дифференцируется структура, которую также можно рассматривать как подиум. Подобная структура в виде радиально расположенных тонкостенных клеток с плотной цитоплазмой в нижней части халазальной зоны нуцеллуса сформированного семязачатка была описана в сем. Euphorbiaceae (Euphorbia milii — Bor, Bouman, 1974; E. geniculata — Bor, Kapil, 1975). Несмотря на отсутствие данных по генезису подиума, его наличие можно предположить и для некоторых других таксонов (Ribes missouriense, Grossulariaceae — Mauritzon, 1933; Cabomba caroliniana, Cabombaceae — Ramji, Padmanabhan, 1965; Hydrostemma (= Barclaya) longifolia,
Barclayaceae — Schneider, 1978).
Вариации в строении и времени дифференциации подиума скоррелированы со степенью массивности нуцеллуса семязачатка. Так, в очень массивном нуцеллусе крассинуцеллятного семязачатка Ceratophyllum demersum (Ceratophyllaceae) подиум начинает формироваться на стадии спорогенной клетки и состоит из 8–12 слоев клеток (Шамров, 1997и). В нуцеллусе из меньшего числа слоев инициация подиума происходит одновременно с началом мегаспорогенеза (Luzula pedemontana, Juncaceae
—Шамров, Анисимова, 1993а), на начальных стадиях (Ungnadia speciosa, Sapindaceae — Anisimova, Shamrov, 1997) либо на поздних стадиях (Juncus filiformis, Juncaceae — Шамров, Анисимова, 1993б) развития зародышевого мешка. В последнем случае подиум представлен 3–5 слоями (рис. 25, 1–9).
Обычно подиум состоит из нескольких слоев таблитчатых клеток с плотной цитоплазмой. Клетки сначала тонкостенные и могут частично дегенерировать в процессе развития семени. Чаще всего они становятся толстостенными. Полностью или частично подиум сохраняется в виде чаши в зрелом семени (Ceratophyllaceae, Euphorbiaceae, Grossulariaceae, Juncaceae, Nymphaeaceae, вероятно, Cyperaceae, Degeneriaceae) (ðèñ. 25, 4, 6, 8, 9).
Âклетках подиума в процессе развития обнаружены белки, крахмал, декстрины, танины, при этом выявляется определенная пространственно-временная связь между накоплением различных веществ в цитоплазме и характером строения клеточных стенок. У Nuphar lutea (Шамров, 1998а) в клетках подиума первыми аккумулируются крахмальные зерна. В период зиготогенеза они появляются снача- ла в центральной части, а к стадии глобулярного зародыша и в остальных клетках. По мере созревания семени из большой части клеток подиума исчезает крах-