

Морфология растений / Шамров И.И. Семязачаток цветковых растений строение, функции, происхождение
.pdf130 |
Шамров И.И. Семязачаток цветковых растений: строение, функции, происхождение |
|
|
семязачатков с перихалазой характерно ответвление постхалазального пучка в интегумент от фуникулярного проводящего пучка (Bouman, 1984) (рис. 41; 43, 3–6). Однако такая особенность строения проводящей системы присуща и ряду таксонов с мезохалазой (Campanulaceae, Cucurbitaceae, Paeoniaceae). Хотя степень участия перихалазы в образовании семенной кожуры увеличивается по сравнению с мезохалазой (главным образом в области ребра семени), все же на большом своем протяжении семенная кожура формируется за счет клеток интегумента.
Лептохалаза характерна для тенуинуцеллятных семязачатков ряда таксонов верхнего уровня системы цветковых растений (Begoniaceae, Gesneriaceae, Orchidaceae и др.). Она представлена небольшой группой клеток (рис. 41; 43, 2), разрушающихся в процессе развития семени. Производные лептохалазы практи- чески не участвуют в построении семенной кожуры. Прокамбиальные клетки, обычно формирующиеся в фуникулусе, в халазе отсутствуют.
Первые стадии развития халазы изучены еще недостаточно. По нашим данным, инициальные слои, участвующие в образовании ее областей, можно выявить на самых ранних стадиях развития семязачатка. В субэпидермальном слое семязачатка с мезохалазой под формирующимся интегументом (наружным интегументом в битегмальных семязачатках) происходят периклинальные деления клеток, за счет которых образуется наружная область халазы. В анатропных семязачатках (Campanulaceae, Gentianaceae, Nymphaeaceae) периклинальные деления обнаруживаются только с дорсальной стороны, а в ортотропных (Barclayaceae — Винтер, 1987) — с обеих сторон семязачатка. Клетки осевого ряда примордия семязачатка ниже формирующейся гипостазы трансформируются во внутреннюю область халазы, в которой дифференцируется проводящий пучок.
Положение халазы в семязачатке и одна из характерных особенностей ее строения (в ней обычно оканчивается проводящий пучок) позволяют предположить, что эта ткань является своеобразным «депо» метаболитов, поступающих как из проводящего пучка, так и из смежных областей семязачатка. Другая ее функция — защитная — наиболее характерна для пахихалазальных семян, у которых семенная кожура образована преимущественно производными халазы.
Фуникулус
Семязачаток снабжен специализированной структурой, с помощью которой осуществляется связь с плацентой. Эта структура получила название фуникулус (ëàò. funiculus — канатик). Термин был предложен Mirbel (1829). Синоним: семяножка.
В фуникулусе дифференцируется проводящая система, обеспечивающая поступление органических и минеральных веществ из завязи в ткани семязачатка и развивающегося семени. На фуникулусе могут развиваться различного рода волоски. Из его тканей формируются обтуратор и ариллус.
Глава 2. Стр т рная дифференциация семязачат а |
131 |
|
|
â è
íè
íê
1
õ
3
4
ô
ç ì
í
ï ï |
ã |
|
õ |
2 |
5
õ
6
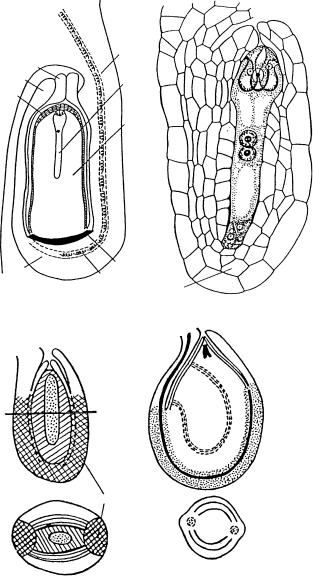
Рис. 43. Строение мезо-, лепто- и перихалазальных семязачатков.
1 — мезохалазальный семязачаток у Nuphar lutea (по: Винтер, Шамров, 1991а); 2 — лептохалахальный семязачаток у Orobanche cumana (по: Анисимова, 1983); 3–6 — перихалахальные семязачатки: 3, 4, 6 — на схемах (3, 4 — ïî: Periasamy, 1962b; 6 — ïî: Boesewinkel, Bouman, 1984), 5 — ó Cissus witiginea (ïî: Periasamy, 1962a); 4, 6 — строение семязачатков на попереч- ных срезах; â è — внутренний интегумент, ã — гипостаза, ç ì — зародышевый мешок, í — нуцеллус, í è — наружный интегумент, í ê — нуцеллярный колпачок, ï ï — проводящий пу- чок, ô — фуникулус, õ — халаза.
132 |
Шамров И.И. Семязачаток цветковых растений: строение, функции, происхождение |
|
|
Âпроцессе созревания между собственно семенем и фуникулусом или рафе
èплацентой образуется отделительный слой, а в дальнейшем возникает след, который получил название хилум, или рубчик. Размер, форма (точковидный, линейный, овальный, округлый и т.д.) и цвет хилума варьируют у разных растений (Данилова, Кирпичников, 1985; Werker, 1997).
Классификация семязачатков в зависимости от наличия или отсутствия семяножки была предложена Корчагиной (1969). Она выделила два типа семязачатков: семяножковый и бессемяножковый, или сидячий (семязачаток имеет широкое основание, ткань которого непосредственно переходит в ткань плаценты, образуя плацентохалазу). Семяножковые семязачатки могут быть прямосемяножковыми и согнутосемяножковыми.
Нами была предложена оригинальная классификация семязачатков, основанная на одном из ведущих принципов типизации — специфике развития и строения, позволяющем дать иную трактовку сидячих семязачатков (Шамров, 1994и; Shamrov, 2002j). Поскольку в литературе наибольшее распространение получил термин «фуникулус», считаем необходимым включить его в названия типов семязачатков. В зависимости от особенностей строения и степени развития фуникулуса, семязачатки можно подразделить на три типа: фуникулярные (семязачаток с фуникулусом), афуникулярные (фуникулус как структура отсутствует) и сессильные (лат. sessilis — cидячий, или прикрепленный основанием к чему-либо; фуникулус морфологически не выражен и семязачаток прикрепляется к плаценте базальной частью рафе) (Шамров, 1994и; Shamrov, 2002j).
Âфуникулярных семязачатках фуникулус может быть длинным или коротким, прямым или изогнутым, закрученным или складчатым (рис. 44, 1–3, 5). Эти особенности строения фуникулуса не приводят к принципиально различным морфологическим типам семязачатка, но в значительной степени определяют его положение в завязи (Maheshwari, 1950; Корчагина, 1969, 1994в; Савченко, 1973; Данилова, Кирпичников, 1985; Werker, 1997). Фуникулус хорошо выражен в семязачатках, занимающих боковое положение в завязи (например, в сем. Nymphaeaceae — Батыгина, 1981; Винтер, Шамров, 1991а, б; особенно у Nuphar lutea, где семязачаток является гипертропным — Винтер, 1987; Винтер, Шамров, 1991а). Длинным фуникулусом часто обладают висячие семязачатки (Amaranthaceae — Kajale, 1937; Brassicaceae — Родионова, 1972, 1983; Шамров, 2002а). В цирцинотропных семязачатках длинный фуникулус сильно закручен (Cactaceae — Archibald, 1939; Plumbaginaceae — Коробова, 1983) (рис. 44, 5).
Âсессильных семязачатках, как правило, обращенных (анатропных, кампилотропных и амфитропных), в процессе развития происходит полное конгенитальное срастание фуникулуса с интегументом и образуется рафе (рис. 44, 6). В этом случае собственно фуникулус морфологически не выражен и семязачатки прикрепляются базальной частью рафе к сильно разросшемуся плацентарному выросту (Campanulaceae, Juncaceae, Paeoniaceae). У некоторых таксонов (Cactaceae) описано разветвление фуникулуса (Buxbaum, 1953; Kapil, Vasil, 1963), что, веро-
Глава 2. Стр т рная дифференциация семязачат а |
133 |
|
|
ô
ð 
1
í
ç
ã
ïëõ
4
ô
2
|
|
3 |
|
ô |
|
|
|
âíèè |
ç ì |
ïë â |
|
í |
|||
|
|
5 |
6 |
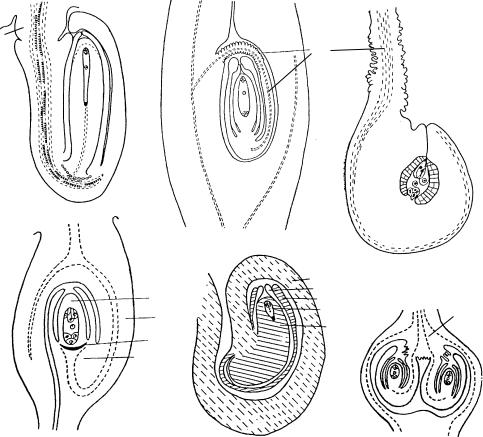
Рис. 44. Типы семязачатков по строению фуникулуса.
1 — анатропный семязачаток с расширенным в основании фуникулусом у Elaeagnus argentea (по: Камелина, Проскурина, 1987); 2 — гипертропный семязачаток с длинным закрученным фуникулусом у Sparganium neglectum (по: Савченко, Комар, 1965); 3 — анатропный семязача- ток с длинным складчатым фуникулусом у Centranthus ruber (по: Камелина, Жукова, 1987); 4 — афуникулярный семязачаток с плацентохалазой у Ceratophyllum demersum (по: Шамров, 1997и); 5 — цирцинотропный семязачаток с многократно закрученным фуникулусом у Opuntia elata (по: Наумова, 1978); 6 — сессильные семязачатки на плацентарных выростах у Luzula pedemontana (по: Шамров, Анисимова, 1993а); â è — внутренний интегумент, ã — гипостаза, ç — завязь, ç ì — зародышевый мешок, í — нуцеллус, í è — наружный интегумент, ïë â — плацентарный вырост, ïëõ — плацентохалаза, ð — ðàôå, ô — фуникулус.
134 |
Шамров И.И. Семязачаток цветковых растений: строение, функции, происхождение |
|
|
ятно, является своеобразным плацентарным выростом, а не базальной ветвящейся частью фуникулуса.
Âафуникулярных семязачатках фуникулус редуцирован полностью, а плацента
ñхалазой формируют единую структуру — плацентохалазу (Poaceae — Александров и др., 1944; Батыгина, 1974; Воронова и др., 2002; Melanthesa rhamnoides,
Euphorbiaceae — Singh, 1968; Ceratophyllaceae — Шамров, 1997и; Santalum album,
Santalaceae — Шамров и др., 2001) (рис. 44, 4).
Обтуратор
В ходе развития семязачатка и завязи для обеспечения прохождения пыльцевых трубок в зародышевый мешок возникают специализированные ткани секреторного типа. Они локализованы в различных частях завязи и семязачатка и представлены эпидермальными удлиненными клетками, которые растут в направлении микропиле и часто закрывают вход в него. Им было дано название обтураторы (ëàò. obturator
— затычка, пробка). Термин предложил Baillon (1858), но описание обтуратора под названием «проводниковая ткань» было дано ранее (Brongiart, 1827).
Первая классификация типов обтуратора была предложена Савченко (1973). Автор выделила несколько типов в зависимости от положения обтуратора в семязачатке и плодолистике: фуникулярный, интегументальный, плацентарный и плодолистиковый. Затем было предложено различать еще нуцеллярный и септальный обтураторы (Веселова, 1989, 1991). По нашему мнению, термин «плодолистиковый обтуратор» не совсем точно отражает его происхождение. Поскольку в этом случае обтуратор возникает из эпидермальных клеток стенки завязи, то его корректнее называть париетальным (лат. parietis — стенка).
Анализ имеющихся данных позволил нам прийти к заключению, что в зависимости от происхождения (из клеток семязачатка или завязи) следует различать два основных типа обтураторов: 1) овулярные (лат. ovulum — семязачаток) — обтураторы, формирующиеся из клеток семязачатка; 2) овариумные (лат. ovarium — завязь) — формирующиеся из клеток завязи. Учитывая положение обтураторов в семязачатке или завязи, в каждом типе можно выделить варианты: в овулярном типе — интегументальный, фуникулярный и нуцеллярный, в овариумном типе — плацентарный, париетальный и септальный обтураторы (рис. 45, 1–6). Существуют также обтураторы промежуточного, овулярно-овариумного типа, которые формируются из клеток как семязачатка, так и завязи (рис. 45, 4, 6) (Шамров,1994ж, 1999б; Shamrov, 2002h).
Èç овулярных обтураторов наиболее распространен фуникулярный обтуратор (рис. 6, 1, 2), который формируется в основании фуникулуса преимущественно анатропных семязачатков (Liliaceae — Савченко, Комар, 1965; Sapindaceae — Anisimova, Shamrov, 1997; Grossulariaceae — Шамров, 1998а). Интегументальный обтуратор (рис. 45, 2, 5) образуется в результате разрастания верхнего края
Глава 2. Стр т рная дифференциация семязачат а |
135 |
|
|
обычно внутреннего интегумента, который может врастать в канал столбика (Urticaceae — Fagerlind, 1944; Poaceae — Савченко, Петрова, 1963), реже — наружного интегумента (Nymphaeaceae — Винтер, 1987; Шамров, Винтер, 1991; Sapindaceae — Anisimova, Shamrov, 1997). Нуцеллярный обтуратор (рис. 45, 4) формируется из апикальных клеток эпидермы нуцеллуса и в виде многослойной клювовидной структуры может выходить за пределы микропиле, контактируя с обтураторами овариумного типа (Nyctaginaceae — Bhargava, 1932; Rosaceae — Савченко, 1959; Euphorbiaceae — Bor, Kapil, 1976; Polygonaceae — Солнцева, 1983; Trapaceae — Батыгина, Колесова, 1985; Caryophyllaceae — Веселова, 1989).
Овариумные обтураторы обнаружены у сравнительно небольшого числа таксонов. Париетальный обтуратор (рис. 45, 6) образуется чаще всего у основания столбика (Rosaceae — Arbeola, Herrero, 1987; такой обтуратор у Daphne laureola èç ñåì. Thymelaeaceae предложено назвать «столбиковым» обтуратором — Терехин, 1996), но иногда и в базальной части завязи (Passerina pectinata, Thymelaeaceae — Fuchs, 1938). Плацентарный обтуратор (рис. 45, 4, 6) возникает из клеток плаценты, граничащих с клетками семязачатка (Liliaceae — Berg, 1962; Alliaceae — Шамров, 1999а). Септальный обтуратор (рис. 6, 3) формируется из клеток перегородок завязи синкарпного гинецея (Caryophyllaceae — Веселова, 1991).
Обтураторы имеют эпидермальное происхождение и состоят из сравнительно крупных клеток с утолщенными наружными стенками, плотной цитоплазмой и большим ядром с несколькими ядрышками. Они имеют признаки передаточных клеток (Johnson et al., 1975; Peterson et al., 1979; Singh, Walles, 1992; Walles, 1992) и являются, подобно клеткам рыльца и канала столбика, разновидностью проводниковой ткани (Tilton, Horner, 1980; Tilton et al., 1984; Clifford, Owens, 1990). В них синтезируются и могут выделяться на поверхность в виде капель жиры, сахара и слизистые вещества (Schweiger, 1905; Савченко, 1973; Arbeola, Herrero, 1987; Шамров, Анисимова, 1993а; Шамров, 1997з).
Интегументальные, нуцеллярные, фуникулярные и париетальные обтураторы обычно возникают в период формирования зародышевого мешка и достигают полного развития к моменту проникновения пыльцевой трубки в полость завязи (Bhargava, 1932; Савченко, Комар, 1965; Савченко, 1973; Винтер, 1987; Шамров, Винтер, 1991). Время дифференциации плацентарных обтураторов варьирует. У одних таксонов (Liliaceae — Шамров, 1999а) они начинают выделяться на стадии примордия семязачатка, у других (Euphorbiaceae — Maheshwari, Johri, 1941; Juncaceae — Шамров, Анисимова, 1993а–в; Alliaceae — Шамров, 1999а) — на стадии двуядерного зародышевого мешка. Считается, что рост пыльцевой трубки происходит по поверхности обтуратора, клетки которого выделяют секрет, однако пусковые механизмы этого процесса мало изучены. Для Prunus persica èç ñåì. Rosaceae установлено, что на продуцирование секрета не влияет опыление, а рост пыльцевой трубки начинается после исчезновения из клеток париетального обтуратора запасного крахмала (Arbeola, Herrero, 1987). Накопление различных плас-
136 |
Шамров И.И. Семязачаток цветковых растений: строение, функции, происхождение |
|
|
ô
1
ñ î
3
è î
5
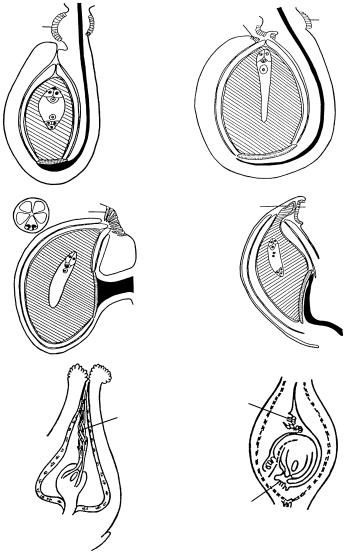
Рис. 45. Типы обтуратора.
è î |
ô î |
2
í î |
ïë î |
4
ïî
ôî 
ïë î |
6 |
|
1 — фуникулярный обтуратор у Ribes aureum (по: Шамров, 1998а, с изменениями); 2 — фуникулярный и интегументальный обтураторы у Victoria amazonica (по: Шамров, Винтер, 1991, с изменениями); 3 — септальный обтуратор у Lychnis coronaria (по: Веселова, 1991, с изменениями); 4 — нуцеллярный и плацентарный обтураторы у Melanthesa rhamnoides (по: Singh, 1968, с изменениями), 5 — интегументальный обтуратор у Leucosyke capitellata (ïî: Fagerlind, 1944), 6 — париетальный, плацентарный и фуникулярный обтураторы у Ruscus hypophyllum (по: Савченко, Комар, 1965); è î — интегументальный обтуратор, í î — нуцеллярный обтуратор, ïë î — плацентарный обтуратор, ï î — париетальный обтуратор, ñ î — септальный обтуратор, ô î — фуникулярный обтуратор.
Глава 2. Стр т рная дифференциация семязачат а |
137 |
|
|
тических веществ в клетках обтуратора, характеризующихся признаками передаточных клеток, а также их своеобразный рост в направлении микропиле свидетельствуют о функции обтуратора как секреторного образования, обеспечивающего питание, рост и проникновение пыльцевой трубки в зародышевый мешок (Maheshwari, 1950; Савченко, 1973; Johnson et al., 1975; Tilton, Horner, 1980; Tilton et al., 1984).
Эволюционные аспекты развития халазы, фуникулуса и обтуратора
Как уже было отмечено, вопросы эволюции и морфологической природы семязачатка и его структур до настоящего времени составляют предмет широкой дискуссии (подробнее об этом см. главу 6). Можно лишь предположить, что у анцестральных семязачатков собственно нуцеллус соединялся с интегументом (интегументами) с помощью своеобразной ножки, а халаза была неразвитой и состояла лишь из производных интегумента (Shamrov, 1998; Шамров, 1999б). Подобное строение имеют ортотропные семязачатки некоторых современных цветковых растений: Juglandaceae, Myricaceae (Kershaw, 1909; Boesewinkel, Bouman, 1967; Van Heel, 1970), Betulaceae, Fagaceae (Корчагина, 1981, 1994).
Постепенное объединение оснований нуцеллуса и интегумента приводило к возникновению мезохалазальных семязачатков. Исходя из представлений Тахтаджяна (1964, 1980) и Терехина (1977, 1991) о модусах эволюционных преобразований органов и тканей, возникновение лептохалазы можно рассматривать в связи с общей редукцией семязачатка как отражение одного из основных направлений эволюции структур — от крупных массивных до мелких немногослойных семязачатков. Однако у ряда высокоспециализированных таксонов (Apocynaceae,
Asclepiadaceae, Rubiaceae, Santalaceae, Theligonaceae) редукция семязачатка, обусловленная главным образом уменьшением размеров нуцеллуса и интегумента, приводила к возникновению не лепто-, а пахихалазальных семязачатков. Появление последних, отмеченное как среди примитивных, так и среди продвинутых таксонов, происходило, вероятно, многократно различными путями. Оно сопровождалось не только уменьшением размеров нуцеллуса и интегумента и, соответственно, возрастанием доли халазы в семязачатке, но и маргинальным срастанием интегументов между собой (возникновение экзопахихалазы), интегументов и нуцеллуса (возникновение перихалазы) и постепенным объединением не только краев, но и оснований нуцеллуса и внутреннего интегумента (возникновение эндопахихалазы) либо обоих интегументов (возникновение экзо-эндопахихалазы). Становление различных типов и вариаций халазы явилось результатом адаптивной эволюции, связанной с особенностями диссеминации (специфика развития и строения семенной кожуры) и прорастания.
138 |
Шамров И.И. Семязачаток цветковых растений: строение, функции, происхождение |
|
|
|
Процесс эволюции семязачатка сопровождался появлением ряда специфиче- |
ских структур. Как считает большинство исследователей, формирование и морфологическое разнообразие фуникулуса и обтуратора обусловлено, вероятно, особенностями экологии опыления. Следует отметить, что, предназначенные для выполнения определенной функции в завязи и семязачатке, они могли в случае сохранения выполнять иные функции в зрелом плоде и семени. Так, в фуникулярных семязачатках разнообразие строения фуникулуса связано с положением семязачатка в завязи (особенно если в завязи развивается более одного семязачатка). Таким образом фуникулус обеспечивает ориентацию микропиле относительно растущих пыльцевых трубок. У некоторых растений, характеризующихся цирцинотропными семязачатками, фуникулус играет активную роль при диссеминации, способствуя, подобно пружине, выбрасыванию семян из плода (Fabaceae — La Rue, 1954; Magnoliaceae — Earle, 1938).
Подобные примеры можно привести и для обтуратора. Эволюционно обтуратор возник как структура, адаптированная к выполнению одной функции — обеспечению роста пыльцевых трубок в направлении микропиле. Обычно обтураторы после оплодотворения разрушаются, но иногда сохраняются в зрелом семени (плацентарный обтуратор у Luzula pedemontana, Juncaceae — Шамров, Анисимова, 1993а, в). В этом случае функция обтуратора в семени неясна. Возможно, он выполняет роль элайосом во время диссеминации.
Важно подчеркнуть, что возникновение морфологического разнообразия фуникулуса и обтуратора, как результат полифилетической эволюции, следует рассматривать сопряженно с другими развивающимися структурами и с учетом общих направлений эволюции семязачатка и плодолистика в целом. Так, выявляется связь между типом обтуратора и особенностями строения фуникулуса. В фуникулярных семязачатках обтуратор обычно развивается из клеток фуникулуса, реже фуникулуса и интегумента (Victoria amazonica, Nymphaeaceae — Винтер, 1987; Шамров, Винтер, 1991). В афуникулярных, сессильных и фуникулярных семяза- чатках со слабо выраженным фуникулусом обтураторы являются производными плаценты (Juncaceae, Paeoniaceae — Шамров, Анисимова, 1993а–в; Шамров, 1997з) либо плаценты и фуникулуса (Gagea stipitata, Liliaceae — Шамров, 1999а).
В заключение хотелось бы отметить, что анализ критериев, положенных в основу существующих классификаций, позволяет типизировать семязачатки, исходя в основном из признаков, присущих какой-либо одной структуре (например, нуцеллусу — крассинуцеллятный, халазе — пахихалазальный, фуникулусу — фуникулярный, интегументу — битегмальный). Создание единой морфогенетиче- ской классификации семязачатка, в которой нашли бы отражение признаки всех или хотя бы нескольких структур, остается на сегодняшний день лишь пожеланием для будущих исследователей.
Глава 3. Критичес ие стадии в развитии семязачат а и семени |
139 |
|
|
Глава 3. КРИТИЧЕСКИЕ СТАДИИ В РАЗВИТИИ СЕМЯЗАЧАТКА И СЕМЕНИ
Периодизация развития семязачатка и семени
Âнастоящее время возникновение и поддержание целостности организма можно считать не только общебиологической, но и наиболее важной комплексной проблемой эволюционной морфологии и эмбриологии. Центральной проблемой современной эмбриологии растений остается изучение закономерностей дифференциации структур, обеспечивающих нормальное развитие зародыша. Одним из наименее исследованных аспектов этой проблемы является выявление морфогенетических
èморфофизиологических корреляций. Сложная система связей, как было установлено Шмальгаузеном (1982), обусловливает развитие организма как целого. При этом прогрессивное усложнение (как в фило-, так и в онтогенезе) означает дифференциацию целого и специфику частей, приобретающих самостоятельные функции.
Как было отмечено ранее, семязачаток и семя являются интегрированными динамическими системами, основные элементы которых определяют специфику развития зародыша и, в конечном счете, семенную репродукцию. Кроме того, характер взаимодействий между ними не только имеет большое значение для познания процессов развития семязачатка и семени, но, несомненно, должен учитываться при решении вопросов классификации эмбриональных структур и эволюции онтогенеза цветковых растений.
Используя системный подход, попытаемся выявить некоторые структурно-фун- кциональные взаимосвязи между нуцеллусом, интегументами, халазой и фуникулусом, с одной стороны, и археспориальной и спорогенной клетками, мегаспороцитом, тетрадой мегаспор, зародышевым мешком, зародышем и эндоспермом, с другой.
Âботанической литературе для характеристики процессов индивидуального развития и оценки их длительности используются различные понятия: этапы, фазы, периоды, стадии, содержание которых и их взаимосвязи в иерархической системе изучены до сих пор недостаточно (Исаева, 1974; Куперман, 1977; Скрипчинский, 1977). Развитие семязачатка представляет собой непрерывный процесс. Однако, чтобы показать специфику строения семязачатка в отдельные моменты, условно выделяют стадии (фазы или этапы) развития, соответствующие образованию той или иной эмбриональной структуры (археспориальная клетка, мегаспороцит, зародышевый мешок и т.д.).