

119
возможность образования кратных химических связей. Сера входит в состав белков, а фосфор – составная часть нуклеиновых кислот.
Первые живые организмы
Строение первых живых организмов хотя и было гораздо совершеннее, чем у коацерватных капелек, но все же оно было несравненно проще нынешних живых существ. Естественный отбор, начавшийся в коацерватных капельках, продолжался и с появлением жизни. В течение долгого времени строение живых существ все более улучшалось, приспособлялось к условиям существования.
Вначале пищей для живых существ были только органические вещества, возникшие из первичных углеводородов. Но с течением времени количество таких веществ уменьшилось. В этих условиях первичные живые организмы выработали в себе способность строить органические вещества из элементов неорганической природы – из углекислоты и воды. В процессе последовательного развития у них появилась способность поглощать энергию солнечного луча, разлагать за счет этой энергии углекислоту и строить в своем теле из ее углерода и воды органические вещества. Так возникли простейшие растения – сине-зеленые водоросли. Остатки сине-зеленых водорослей обнаруживаются в древнейших отложениях земной коры.
Другие живые существа сохранили прежний способ питания, но пищей им стали служить первичные растения. Так возникли в своем первоначальном виде животные.
На заре жизни и растения, и животные были мельчайшими одноклеточными существами, подобными живущим в наше время бактериям, сине-зеленым водорослям, амебам. Большим событием в истории последовательного развития живой природы стало возникновение многоклеточных организмов, т.е. живых существ, состоящих из многих клеток, объединенных в один организм. Постепенно, но значительно быстрее, чем раньше, живые организмы становились все сложнее и разнообразнее.
С образованием сложных ультра молекулярных систем (пробионтов) включающих нуклеиновые кислоты, белки ферменты и механизм генетического кода, появляется жизнь на Земле. Пробионты нуждались в различных химических соединениях – нуклеотидах, аминокислотах и др. Изза низкой степени генетической информации, пробионты обладали достаточно ограниченными возможностями. Дело в том, что они использовали для своего роста готовые органические соединения, синтезированные в ходе химической эволюции, и если бы жизнь на своем раннем этапе существовала только в форме одного вида организмов, то первичный бульон был бы достаточно быстро исчерпан.
Однако благодаря тенденции к приобретению большого разнообразия свойств, и в первую очередь, к возникновению способности синтезировать органические вещества из неорганических соединений с использованием солнечного света, этого не произошло.
В начале следующего этапа образуются биологические мембраныорганеллы, ответственные за форму, структуру и активность клетки. Биологические мембраны построены из агрегатов белков и липидов,
120
способных отграничить органическое вещество от среды и служить защитной молекулярной оболочкой. Предполагается, что образование мембран могло начаться еще в процессе формирования коацерватов. Но для перехода от коацерватов к живой материи были необходимы не только мембраны, но и катализаторы химических процессов – ферменты или энзимы. Отбор коацерватов усиливал накопление белково-подобных полимеров, ответственных за ускорение химических реакций. Результаты отбора фиксировались в строении нуклеиновых кислот. Система успешно работающих последовательностей нуклеотидов в ДНК усовершенствовалась именно путем отбора. Возникновение самоорганизации зависело как от исходных химических предпосылок, так и от конкретных условий земной среды. Самоорганизация возникла как реакция на определенные условия. При самоорганизации отсеивалось множество различных неудачных вариантов, до тех пор, пока основные черты строения нуклеиновых кислот и белков не достигли оптимального соотношения с точки зрения естественного отбора.
Благодаря предбиологическому отбору самих систем, а не только отдельных молекул, системы приобрели способность совершенствовать свою организацию. Это был уже следующий уровень биохимической эволюции, который обеспечивал возрастание их информационных возможностей. На последнем этапе эволюции обособленных органических систем сформировался генетический код. После образования генетического кода эволюция развивается вариациями. Чем дальше она продвигается во времени, тем многочисленнее и сложнёе были вариации.
Однажды возникнув, жизнь стала развиваться быстрыми темпами показывая ускорение эволюции во времени. Так, развитие от первичных пробионтов до аэробных форм потребовало около 3 млрд. лет, тогда как с момента возникновения наземных растений и животных прошло около 500 млн. лет; птицы и млекопитающие развились от первых наземных позвоночных за 100 млн. лет, приматы выделились за 12-15 млн. лет, для становления человека потребовалось около 3 млн. лет. (табл. 1 ) В таблице приводится в хронологическом порядке основные этапы развития животного и растительного мира за всю историю Земли.
Истинная основа жизни образовалась в результате появления клетки, в которой биологические мембраны объединили отдельные органеллы в единое целое.
Первые клетки были примитивны и не имели ядра. Но такие клетки существуют и в настоящее время. Удивительно, ведь они появились более 3 млрд. лет назад.
Первые клетки были прообразом всех живых организмов: растений, животных, бактерий. Позже, в процессе эволюции, под воздействием дарвиновских законов естественного отбора клетки совершенствовались и появились специализированные клетки высших многоклеточных, растений и животных – метафитов и метазоа.
В качестве объединяющей зависимости между химической эволюцией переходящей затем в биохимическую и биологическую эволюцию можно привести следующую: 1. атомы; 2. простые молекулы; 3. сложные
|
|
|
|
|
|
|
|
|
|
121 |
|
макромолекулы |
и |
ультра |
молекулярные |
|
системы |
(пробионты); |
|||||
4. одноклеточные организмы. |
|
|
|
|
|
|
|
|
|
||
На это потребовалось более 3 миллиардов лет, и это было самым |
|
||||||||||
трудным. Не поддается перечислению огромное количество вариантов |
|
||||||||||
развития исходных углеродных соединений. Однако самым важным был |
|
||||||||||
результат – возникновение жизни на Земле. |
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
Таблица 2. |
|
|
|
|
Хронологический порядок развития |
|
|
|
|
|||||
|
животного и растительного мира планеты Земля |
|
|
||||||||
|
|
|
|
||||||||
Эры, периоды |
|
Главные этапы развития |
|
||||||||
|
|
|
|
растительного и животного мира |
|
||||||
|
1 |
|
|
|
|
|
2 |
|
|
|
|
Архейская эра |
|
|
|
Появление |
|
первых |
простейших |
||||
Время от начала эры, периода более |
живых существ, водорослей и |
||||||||||
3 млрд. лет |
|
|
|
бактерий. |
|
Первые |
известковые |
||||
|
|
|
|
водорослевые |
|
постройки |
– |
||||
|
|
|
|
строматолиты |
|
|
|
|
|
||
Протерозойская эра |
|
|
Массовое развитие сине-зеленых |
||||||||
|
|
|
|
водорослей. Появление (около 1 млрд. |
|||||||
Время от начала эры, периода 2500 |
лет) |
первых |
животных |
- |
|||||||
млн. лет |
|
|
|
кишечнополостных, червей и др. |
|
||||||
Палеозойская эра (длительность 345 млн. лет) |
|
|
|
||||||||
Кембрийский (длительность 70 млн. |
В начале периода массовое появление |
||||||||||
лет) |
|
|
|
скелетов (внутренних и наружных - |
|||||||
|
|
|
|
раковин) |
|
у |
различных |
групп |
|||
Время от начала эры, периода 570 |
животных. |
|
Массовое |
|
развитие |
||||||
млн. лет |
|
|
|
известковых водорослей |
|
|
|
||||
Ордовикский (длительность 60 - 65 |
На значительной |
части |
Русской |
||||||||
млн. лет) |
|
|
|
платформы суша. В Сибири неглу- |
|||||||
Время от начала эры, периода 500 |
бокое открытое море. Распростра- |
||||||||||
млн. лет |
|
|
|
нение трилобитов и граптолитов. |
|||||||
|
|
|
|
Первые бесчелюстные позвоночные |
|
||||||
Силурийский (длительность 30 млн. |
Появляются |
|
наземные |
растения |
– |
||||||
лет) |
|
|
|
псилофиты. |
|
|
|
|
|
|
|
Время от начала эры, периода 435 - |
Распространяются |
панцирные |
и |
||||||||
440 млн. лет |
|
|
|
хрящевые рыбы, граптолиты и |
|||||||
|
|
|
|
брахиоподы |
|
|
|
|
|
|
|
Девонский (длительность 50 - 60 |
Широко |
распространена |
псилофи- |
||||||||
млн. лет) |
|
|
|
товая флора, появляются папорот- |
|||||||
Время от начала эры, периода 395 - |
ники. Широкое развитие кистеперых |
||||||||||
410 млн. лет |
|
|
|
и двоякодышащих рыб. Первые |
|||||||
|
|
|
|
земноводные – стегоцефалы |
|
|
|||||
Каменноугольный, |
или карбон |
Господство |
гигантских |
плауновых. |
|||||||
(длительность 65 млн. лет) |
|
Развитие земноводных, насекомых, |
|||||||||
Время от начала эры, периода 345 - |
возникновение пресмыкающихся |
|
|||||||||
350 млн. лет |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
122 |
|
|
|
Продолжение таблицы 2. |
|||||
1 |
|
|
|
2 |
|
|
|
|
Пермский (длительность 55 млн. |
Растут |
гигантские |
папоротники, |
|||||
лет) |
появляются |
|
первые |
голосемянные. |
||||
Время от начала эры, периода 280 - |
Развиваются |
пресмыкающиеся |
и |
|||||
285 млн. лет |
крупные земноводные. |
Вымирают |
||||||
|
табуляты, трилобиты и многие |
|||||||
|
брахиоподы |
|
|
|
|
|
|
|
Мезозойская эра (длительность 160 млн. лет) |
|
|
|
|||||
Триасовый (длительность 35 млн. |
Древовидные папоротники и голосе- |
|||||||
лет) |
мянные. |
Вымирают |
|
гигантские |
||||
|
земноводные. |
Развитие |
основных |
|||||
Время от начала эры, периода 225 - |
групп мезозойских пресмыкающихся. |
|||||||
230 млн. лет |
Появляются |
|
белемниты, |
костистые |
||||
|
рыбы, шестилучевые кораллы |
|
||||||
Юрский (длительность 54 - 58 млн. |
Распространяются |
|
голосемянные |
|||||
лет) |
растения. Разнообразие пресмыкаю- |
|||||||
|
щихся – летающие, плавающие, |
|||||||
Время от начала эры, периода 190 - |
гигантские |
формы. |
Распространение |
|||||
195 млн. лет |
аммонитов и белемнитов. Появляются |
|||||||
|
первые |
|
зубастые |
|
птицы |
и |
||
|
млекопитающие |
|
|
|
|
|||
Меловой (длительность 70 млн. лет) |
Появление и резкое увеличение в |
|||||||
|
конце |
периода |
покрытосемянных |
|||||
Время от начала эры, периода 136 - |
растений. Расцвет и вымирание |
|||||||
137 млн. лет |
крупных ящеров. Появление беззубых |
|||||||
|
птиц. Редкие примитивные млекопи- |
|||||||
|
тающие. Вымирают аммониты и |
|||||||
|
белемниты |
|
|
|
|
|
|
|
Третичный (длительность 64 - 66 млн. лет) |
|
|
|
|
||||
Палеогеновый (длительность 41-42 |
Распространение |
покрытосемянных |
||||||
млн. лет) |
растений. |
Развитие |
разнообразных |
|||||
|
групп |
млекопитающих, |
появляются |
|||||
Время от начала эры, периода 65 - 67 |
парнокопытные, хищники, китообраз- |
|||||||
млн. лет |
ные. |
Широко |
распространяются |
|||||
|
беззубые птицы |
|
|
|
|
|||
Неогеновый (длительность 24 - 25 |
Богатая |
|
|
и |
|
разнообразная |
||
млн. лет). |
растительность. Появляются лошади, |
|||||||
Время от начала эры, периода 25 - |
жирафы, саблезубые тигры |
|
||||||
26 млн. лет |
|
|
|
|
|
|
|
|
Кайнозойская эра (длительность 65 - 67 млн. лет) |
|
|
||||||
Четвертичный, или антропоген |
С начала периода животный и |
|||||||
|
растительный |
мир |
|
близки |
к |
|||
(длительность 1 млн. лет) |
современному. В Европе и Сибири |
|||||||
|
водились мамонты и носороги. |
|||||||
|
Появился человек |
|
|
|
|
|||
123
Многократные в течение истории земной коры и нередко могучие проявления тектогенеза и магматизма создавали не только все более сложное ее глубинное строение, но обусловливали постоянное изменение внешнего облика Земли, ее рельефа. В одни геологические эпохи преимущественно горный, возвышенный рельеф сочетался с господством суши над морем; в другие' эпохи выровненный невысокий пенепленизированный рельеф приводил к обширным, безбрежным степным просторам с подчиненными им по площади морскими пространствами; в иные же времена трансгрессии морей покрывали обширнейшие территории, резко изменяя отношение суши к морю. Все это создавало дифференциацию физико-географических условий, формировало микро- и мезоклиматические различия, которые тесно переплетались с эволюцией органического мира.
Определяющим фактором в развитии жизни является климат и солнечная радиация. Корни растительной жизни уходят в докембрий. Растения существуют на Земле уже более миллиарда лет. Наиболее древними являются морские известковые водоросли водоросли. Они широко распространены по всему миру, причем роль их в осаждении карбонатов особенно велика в теплых мелких морях. Мощное накопление строматолитов, имеющих, вероятнее всего, известково-водорослевое происхождение, на плитах северного полушария свидетельствует, что современные холодные и субарктические области в докембрий покрывались теплыми неглубокими морями.
Частая встреча водорослевых рифов среди осадков раннего палеозоя на континентах северного полушария может быть принята за указание на широкое вторжение мелких теплых морей по направлению к полюсу.
Особенную ценность для распознания климатических обстановок имеет наземная растительность, появление которой относится к позднему силуру и раннему девону. В истории флоры и фауны этот этап надо признать важнейшим. Выход растений из вод на сушу обусловил развитие сухопутных животных, особенно позвоночных (табл. 2).
Первая наземная растительность отличалась простотой строения и очень широким географическим распространением. Остатки девонской наземной флоры находят на Фалклендских островах, Шпицбергене, во внутренних областях Азии и Северной Америки. Слои с флорой девона, выходящие изпод тающих ледников Шпицбергена, наводят на мысль о более однородном и мягком климате на обширных площадях тогдашних материков и на отсутствие ледниковых условий в полярных областях.
К концу девона появились настоящие деревья, из которых состояли первобытные леса. Покрытие ранее голой земли растительностью, несомненно, должно было резко ослабить эрозионные процессы.
Чрезвычайный интерес приобретают пышная наземная флора карбона и мощное тогда угленакопление. Остатки раннекарбоновой растительности во множестве находят в соответствующего возраста пластах, в северном и южном полушариях, что показывает на слабую климатическую зональность, между высокими и низкими широтами. Угленакопление, шедшее главным образом в болотах, характеризует скорее большую влажность, а не высокие
124
среднегодовые температуры. У деревьев каменноугольных болот обычно отсутствие годичных колец.
Очевидно, в большей части периода не было зимних холодов и сезонности выпадения атмосферных осадков. Но в позднем палеозое, в слоях пермо-карбона в южном полушарии нередки куски дерева с ярко выраженными годичными кольцами, что надо связать с сезонной периодичностью и зимними холодами в современной тропической Африке и Индостане. Там, как нам известно, найдены тиллиты, т.е. древние морены континентального оледенения. Это показывает, что в позднем палеозое был резкий контраст климатов северного и южного полушарий.
Флора триаса говорит о возврате более теплого климата в умеренных шиторах. По-видимому, быстрое развитие хвойных и других простейших семенных растений указывает на сухой тогда климат в умеренных широтах.
Богатая флора юры, существовавшая севернее 80° с. ш. в Гренландии, на Земле Франца-Иосифа, а также в Антарктиде, говорит о неледниковом климате в полярных областях. Однако наличие в юре углеобразующих болот, иногда вблизи от южного полюса, свидетельствует о теплом и влажном климате. Поразительным надо признать факт развития растительности в обстановке полярной ночи.
Ныне вездесущая группа покрытосемянных цветковых растений, сформировавшаяся к верхнемеловым эпохам, была весьма разнообразна. Начиная с мела до эоцена, возможно до олигоцена, повсеместно развиваются лиственные леса с твердой древесиной, указывая на длительное господство теплого, влажного и умеренного климата даже в полярных областях. В раннем кайнозое были обильно покрыты средние широты растительностью, отвечающей теперь климату тропиков и субтропиков. Надо предполагать отсутствие зимнего замерзания в очень северных областях, что указывает на малое различие температур между высокими и низкими широтами, как это существует теперь.
Однако к середине третичного периода листопадная флора начинает опускаться в южные широты и вместе с тем развивается травяной покров. В позднем кайнозое быстрая эволюция трав сыграла важную роль в истории многих групп млекопитающих, включая человека. Похолодание климата продолжалось в позднетретичное время. Предполагают, что ледниковые условия в полярных областях появились в среднем плиоцене.
Изучение остатков животных также проливает свет на историю климата; особенно чувствительны к нему позвоночные, которые к тому же обитают в самых различных условиях: на любой глубине в океанах, на любой высоте на суше, живут и в воздухе.
История позвоночных восходит, как видим, к началу силура. Однако, их существование отмечено и в кембрии; остатков оттуда до нас не дошло, вследствие того, что тогда жили беспанцирные позвоночные. Все же удается находить разрозненные чешуйки. Однако основательные знания о позвоночных относятся к девону. Тогда распространеннейшими были рыбы, и девон нередко называют «эрой рыб». Наряду с панцирными рыбами были ранние предки современных рыб; в девоне обитали простейшие акулы,
125
первые двоякодышащие и кроссоптеригоидные рыбы; последние дали начало земноводным.
Условия находок остатков девонских позвоночных дают возможность предполагать, что они жили в пресных водах или в мелких устьях рек, впадавших в моря. Вероятно, что позвоночные зародились в пресной воде, и она долго была их местообитанием. Для познания климатических особенностей это исключительно важно, так как океанический животный мир менее подвержен влиянию климатических изменений, чем суша. Остатки девонских позвоночных часто находят в Северной Европе и Северной Америке. Древние красные песчаники Шотландии уже более ста лет являются источником богатейших находок ископаемых остатков позвоночных. Они же известны с Оркнейских островов, Прибалтики, Шпицбергена, Гренландии и прослежены в США на юг через Квебек до штатов Нью-Йорк и Огайо, а также на запад до Аризоны и Вайоминга. Встречены позвоночные в пластах девона в Новом Южном Уэльсе Австралии, Антарктике. Отсюда следует, что девонские позвоночные были распространены почти по всему земному шару. Будучи обитателями мелких наземных водоемов, рек, озер, иногда заливов, девонские рыбы могут дать указание, что в девоне был широко распространен однородный климат.
Отмечено, преобладание хоановых, дышащих атмосферным воздухом рыб, двоякодышащих и кроссоптеригий. Это прекрасный показатель того, что девонские водоемы часто периодически сокращались или даже совсем пересыхали. Ныне живущие в Австралии двоякодышащие при обмелении и загрязнении рек выходят на сушу и дышат атмосферным воздухом.
ВЮжной Америке и Африке они зарываются в ил и, используя атмосферный воздух, в течение нескольких месяцев выдерживают полное пересыхание водоемов. Приведенные факты наводят на мысль, что, в общем, климат девона не был засушливым, а характеризовался сухими сезонами, когда пресные водоемы сильно сокращались. Под влиянием периодических чередований сухих и влажных сезонов кроссоптеригий могли превращаться в земноводных.
Взасушливые сезоны эти рыбы принуждены были переползать из одной лужи в другую, что явилось первым выходом позвоночных на сушу. Остатки первых земноводных встречаются в верхнедевонских отложениях восточной Гренландии, где, следовательно, тогда был сравнительно теплый климат. От экватора к полюсам климат изменялся весьма постепенно, и температурные различия между сезонами не были очень большими, как теперь.
Вкарбоне полностью завершился выход позвоночных на сушу. На Земле тогда процветали обширные леса, давшие материал для каменного угля. Огромные тропические болота покрывали сушу, и в них кишели земноводные. Теплый и влажный климат благоприятствовал появлению пресмыкающихся.
Остатки земноводных и пресмыкающихся карбона найдены в Англии, Шотландии, Франции, Аллеганском районе Пенсильвании, Западной Виргинии, Иллинойсе, Нью-Брансуике, Новой Шотландии. Сходство находимых фаун исключительно велико, что указывает не только на однородный климат, но и на связь между материками, обеспечившую быстрое распространение наземных животных.
126
Несмотря на обширное оледенение в южном полушарии к концу карбона, оно оказало малое влияние на фауны, обитавшие в северном полушарии: в болотах и реках Северной Европы и Америки продолжали процветать земноводные и пресмыкающиеся.
Если карбон был временем господства земноводных, то в пермской фауне преобладали пресмыкающиеся. Соответствующие остатки найдены в пермских глинистых красных песчаниках Техаса, во Франции, ГДР, ФРГ, Шотландии, в Европейской части России на Северной Двине, в формации Карру в Южной Африке, в Азии, Австралии, Бразилии. Следовательно, пресмыкающиеся в пермское время распространились по всему свету. Смешанные скопления рыб, земноводных и пресмыкающихся в пермских слоях Техаса, Нью-Мексико и Оклахомы свидетельствуют о чередовании там засушливых и влажных периодов.
Вформации Карру находят изобилие травоядных и хищных пресмыкающихся, весьма сходных с нашей двинской фауной.
Сопоставляя приведенные данные, придем к заключению, что климат перми был разнообразным, возможно с заметным чередованием времен года. Температуры от современной Арктики до Антарктики в течение большей части перми мало отличались, были высокими и умеренными, но не низкими.
Подобные пермским, красноцветные породы, характерны и для триаса Северной Америки вдоль берега Атлантического океана, Шотландии, Центральной Европы, Южной Африки, районов Западного Китая, Индии, южной Бразилии. Это объясняют тем, что триас был периодом чередования влажных и засушливых сезонов, вызвавшего окисление железа в осаждавшихся породах и придание им красного цвета.
Из перми в триас перешли лабиринтодонтные амфибии. Их остатки свидетельствуют о влажном климате, множестве рек и водоемов: на севере России, в Южной Африке, Южной Аргентине, Восточной Австралии, до Восточной Гренландии и Шпицбергена. Открывается огромная территория от 80° с.ш. до 40° ю.ш. с бывшими на ней довольно однородными влажными условиями климата с умеренной температурой.
Вотложениях триаса севера России, Шотландии, юга Африки и южной Бразилии часты остатки крупных пресмыкающихся, говорящие об умеренной
итеплой температуре. Допускают распространение от экватора к полюсам тропических, субтропических и умеренно теплых климатов с чередованием влажных и сухих сезонов.
Существование огромных динозавров, появившихся в начале мезозоя в Южной Африке, Центральной Европе, Северной Америке, Западном Китае, служит признаком мягкого климата и обилия пищи.
Втриасе появились ихтиозаврьи (рыбоящеры); они должны быть тепловодными пресмыкающимися, и замечательно их нахождение в приэкваториальной полосе и на Шпицбергене.
Распространение в ряде мест, например, на юго-западе Соединенных Штатов, эоловых, дюнных песков, относимых к нижней юре, показывает, что в начале этого периода на обширных районах был аридный, сухой климат. Он вскоре сменился более влажным. Большая часть юры отличалась слабым
разнообразием климатических условий за всю историю Земли.
127
Господствовала низменная болотистая суша, на которую часто трансгрессировали морские воды. Разрасталась буйная растительность, среди многих наземных и морских пресмыкающихся наблюдалось стремление к гигантизму. Все это показывает на существование тогда весьма однородного и мягкого климата, вероятно большей частью тропического и субтропического. В Северной Америке и Восточной Африке обитало множество огромных травоядных динозавров, броненосцев – стегозавров, крокодилов и черепах.
Верхнеюрские отложения в Европе богаты остатками крупных динозавров, гигантских ихтиозавров и плезиозавров. Смешение остатков наземных и водных пресмыкающихся указывает на существование здесь низких островов с открытым между ними морем.
Вмеловых континентальных отложениях Северной Европы, востока Канады, Южной Африки, Австралии и Патагонии содержатся многочисленные остатки рептилий. Также широко распространены в морских осадках ихтиозавры, плезиозавры, мезозавры, свидетельствуя о проникновении теплых морей далеко в северные и южные широты Земли. Следовательно, на огромных пространствах были однообразные и благоприятные климатические условия для процветания крупных пресмыкающихся на большей части земной поверхности.
Однако ни один из меловых динозавров не достигал тех огромных размеров, какими отличались животные юры. Количественно их также стало значительно меньше. Это можно объяснить тем, что климаты мелового периода, по-видимому, не были такими однородно тропическими или субтропическими, как в юре; растительность также сильно обеднела.
Необходимо отметить, что как растительность, так и животные второй половины мела все больше и больше приобретали современный облик. Например, появившиеся в раннем мезозое крокодилы, черепахи, ящеры и змеи пережили весь меловой период и существуют в настоящее время. К мелу же относится и появление плацентарных млекопитающих.
Вобщем, характер позвоночных в меловых слоях свидетельствует о достаточно разнообразных климатах с хорошо выраженным чередованием времен года при отсутствии, однако, холодов.
Справедливо считают конец мела и начало кайнозоя временем огромных кризисов в истории~3емли и жизни. Мощный тектогенез альпийского цикла, создав грандиозные горные цепи, резко изменил рельеф нашей планеты, а вместе с тем климаты и общие физико-географические условия среды. Это вызвало колоссальное вымирание гигантов пресмыкающихся и внезапный бурный расцвет млекопитающих. Появились разнообразные, живущие и в настоящее время птицы. Но, став господствующими, птицы и млекопитающие не дают нам таких надежных показателей среды, какими были пресмыкающиеся. Климат сделался весьма разнообразным с отчетливым разграничением климатических поясов. В самом конце кайнозоя и установилось чередование сезонов: влажных и сухих в экваториальной зоне, теплых и холодных – в более высоких широтах. Климатическая дифференциация имела место во все времена, но наиболее ярка она проявилялась в периоды похолоданий и оледенений. Дифференциация
128
климата объясняется округлой поверхностью земного шара, большим поступлением солнечной энергии у экватора и меньшим у полюсов.
6.3.Биохимические основы наследственности
В1869г. швейцарский биохимик Ф. Мишер впервые описал вещество, содержащееся в ядрах клеток, и назвал его нуклеином, а позже оно было переименовано в нуклеиновые кислоты (от лат. nucleus – ядро). К ним относятся дезоксирибонуклеиновая кислота – ДНК (в её состав входит сахар дезоксирибоза) и рибонуклеиновая кислота – РНК (входит сахар рибоза).
Но уже в 1914 году доцент Петербургского университета А. Щепотьев предположил, что веществами, имеющими генетическое значение, являются прежде всего нуклеиновые кислоты. Однако мысль о генетическом значении нуклеиновых кислот в то время не получила распространения. Напротив, длительное время, вплоть до начала 40-х годов, считали, что генный материал организмов – это белки или нуклеопротеиды. Генетическая специфичность нуклеопротеидов связывалась с их белковой частью, но не нуклеиновокислотной.
В1928 г. бактериолог Ф. Гриффит изучал бескапсульные невирулентные пневмококки (не вызывающие заболевания) и вирулентные в полисахаридной капсуле (вызывающие воспаление легких) для получения вакцины против пневмококка. Он показал, что при инъекции мышам живых бескапсульных пневмококков мыши выживали, а при введении живых капсульных – погибали. При введении смеси убитых при нагревании капсульных и живых бескапсульных пневмококков мыши погибали, из них удалось выделить живых капсульных пневмококков. Таким образом, способность образовывать капсулу перешла от убитого капсульного пневмококка к живому бескапсульному.
В1944 г. О. Эвери с сотрудниками выяснили природу этого загадочного явления. Фактором, превращающим непатогенные (бескапсульные) в патогенные (капсульные) пневмококки, является ДНК, а само явление назвали трансформацией (от лат. transformatio – преобразование, превращение). Следовательно, трансформация – это преобразование признака у одного штамма бактерии в результате проникновения в нее ДНК другого штамма. Явление трансформации стало одним из основных доказательств того, что ДНК является носителем генетической (наследственной) информации.
Позже, в 1952 г. Дж. Ледербергом и Н. Циндером была выявлена передача генетического материала от одного штамма бактерий другому с помощью бактериофага, это было названо трансдукцией (от лат. transductio – перемещение, передача). U-образная трубка в нижней части разделена бактериальным фильтром. В одну половину были помещены штаммы сальмонеллы, не синтезирующие аминокислоту триптофан (Т-), а в другую – сальмонеллы, синтезирующие триптофан (Т+) и бактериофаги. После инкубации среди сальмонелл, не синтезирующих триптофан, были выделены бактерии Т+. Это объясняется тем, что бактериофаги проходили через бактериальный фильтр и переносили части ДНК от бактерии Т+ к бактериям Т-.
129
Решающим прямым доказательством в пользу генетической роли ДНК стала возможность конструирования рекомбинантных молекул ДНК методом генной инженерии.
Что касается косвенных доказательств, то их существует несколько. Одно из первых сводится к тому, что локализация ДНК в клетках специфична: ее обнаруживают в ядрах клеток, в частности в хромосомах, в митохондриях и хлоропластах. У многих микроорганизмов ДНК локализована только в ядерной области (нуклеоид) или в цитоплазме в виде плазмид. Для организмов каждого вида характерно определенное количество ДНК на клетку, а также то, что в диплоидных клетках ДНК содержится вдвое больше, чем в гаплоидных. Будучи постоянным, у организмов всех видов, содержание ДНК в клетке не подвержено влиянию со стороны физиологических факторов, включая пол и старение, а также неблагоприятных воздействий, например, голодания, повышенных или пониженных температур.
Далее, к косвенным доказательствам генетической специфичности ДНК могут быть отнесены также данные о способности искусственно синтезированных аналогов пуриновых и пиримидиновых оснований, вызывать наследственные изменения клеток вследствие прямого включения этих аналогов в ДНК клеток. Наконец, косвенным доказательством является и то, что мутагенный эффект УФ-излучения тесно связан со спектром поглощения его молекулами ДНК.
Исследование структуры молекулы ДНК проводилось многими учеными. И только в 1953 г., используя все накопленные биологические и физикохимические знания, Д. Уотсон и Ф. Крик открыли двухцепочечную спиральную (пространственную) структуру молекулы ДНК. Молекула ДНК не случайно имеет форму двойной спирали. Длина молекулы ДНК может достигать в длину 40м, в то же время молекула заключается в ядре, размеры которого составляют всего несколько микрон. Упаковка ДНК в ядре достигается ее плотным закручиванием. Уотсон и Крик показали, что уложенные стопками основания внутри спирали располагаются на расстоянии 0,34 нм друг от друга.
Каждая цепь – это полимер, мономерами которого являются нуклеотиды. Каждый нуклеотид состоит из сахара дезоксирибозы, остатка фосфорной кислоты и одного из четырех азотистых оснований (аденин, гуанин, тимин, цитозин). Две цепи ДНК соединяются слабыми водородными связями между азотистыми основаниями по принципу комплементарности: аденин дополняется тимином, гуанин – цитозином. Относительно состава ДНК открыт ряд общих закономерностей, которые известны как правила Чаргаффа:
1.Сумма нуклеотидов, содержащих пуриновые основания, равна сумме нуклеотидов, содержащих пиримидиновые основания (А + Г = Т + Ц, или
А+ Г / Т + Ц = 1).
2.Содержание аденина равно содержанию тимина, а гуанина – содержанию цитозина (А = Т, или А / Т = 1, или Г = Ц, или ГУЦ = 1).
3.Г + Т = А + Ц, или Г + Т / А + Ц = 1.
4.ДНК из разных источников могут иметь различия, обусловленные в одних случаях преобладанием аденина над гуанином и тимина над цитозином
130
(А + Т > Г + Ц), в других случаях – преобладанием гуанина и цитозина над аденином и тимином (Г + Ц > А + Т).
Молекулы ДНК и РНК имеют полярность (или направление), так как все фосфодиэфирные межнуклеотидные связи ориентированы вдоль цепи одинаковым образом. Из-за полярности каждая цепь полинуклеотидов имеет 5'-конец и 3'-конец.
В строении ДНК различают – первичную, вторичную и третичную структуру. Первичная структура ДНК (как и РНК) определяется последовательностьюнуклеотидов и характером их связей между остатком пентозы и фосфатом. Молекула ДНК состоит из нуклеотидных цепей, скелетную основу которых составляют чередующиеся сахарные и фосфатные группы, объединенные 3', 5'-фосфодиэфирньми связями, а боковые группы представлены тем или иным основанием (одним из четырех). Таким образом, последовательно располагающиеся нуклеотиды ковалентно связаны фосфодиэфирными связями между сахарным остатком и фосфатом и в результате этого объединены в полинуклеотидную цепь.
Представления о вторичной структуре ДНК были сформулирована Д. Уотсоном и Ф. Криком в 1953 году на основании биохимических и рентгеностуктурных данных о ДНК и сводятся к ряду принципов, а именно:
1. Молекула ДНК построена из двух скрученных слева направо спиралевидных полинуклеотидных цепей, причем каждый виток спирали соответствует 10 азотистым основаниям, или 3,4 нм по оси.
Сейчас молекулы ДНК, цепи которых скручены направо, называют В- формой.
Обе цепи объединены в результате закручивания одной вокруг другой по общей оси. Из-за противоположной последовательности атомов обе цепи инвертированы относительно одна другой, т.е. данное направление вдоль молекулы есть 3'-5' для одной цепи и 5'-3' для другой.
3.Сахарофосфатные группы располагаются на внешней стороне двойной спирали, тогда как основания находятся внутри спирали. Диаметр молекулы составляет 20 нм, расстояния между отдельными азотистыми основаниями в молекуле равны 0,34 нм.
4.Цепи в молекуле не идентичны, но комплементарны и удерживаются слабыми водородными связями между азотистыми основаниями, причем спаривание азотистых оснований для связывания цепей имеет специфический характер. Водородные связи возникают не между любыми азотистыми основаниями, а избирательно между пуриновым основанием одной цепи и пиримидиновым – другой. Так, при взаимодействии аденина и тимина образуется две водородные связи, а гуанина и цитозина – три.
Таким образом, в соответствии с моделью Д. Уотсона и Ф. Крика ДНК представляет собой скрученную в направлении слева направо двойную спираль, в которой пары азотистых оснований А – ТиГ – Цв комплементарных полинуклеотидных цепях подобны перекладинам лестницы, а сахарофосфатные цепи – её каркасу. Пуриновые и пиримидиновые основания комплементарных цепей соединены водородными связями в пары. Для цепей молекулы ДНК характерна противоположная полярность. Спираль ДНК образована двумя цепями, закрученными одна вокруг другой, причем на
131
каждый виток (шаг) спирали приходится 10 пар оснований, а расстояние между соседними витками составляет 3,4 нм. Одна из особенностей структуры ДНК заключается также в том, что двойная спираль имеет упорядоченный характер, поскольку каждая связь «основание – сахар» находится на одинаковом расстоянии от оси спирали и повернута на 36 градусов.
ДНК в клетках встречается в основном в двухцепочечной форме. Установлено, что в молекулах ДНК имеются районы, цепи в которых закручены налево. Эти районы получили название Z-форм.
Третичная структура ДНК связана с трехмерной пространственной конфигурацией ее молекул. К настоящему времени третичная структура ДНК еще изучена недостаточно, однако известно, что она зависит от внутримолекулярных условий.
О размерах молекул ДНК обычно судят по их молекулярной массе, выраженной в дальтонах, и длине – в количестве пар оснований.
Для характеристики строения ДНК используют также такие физические константы, как плотность ее при центрифугировании в градиенте тяжелых металлов, а также температура плавления. Первая константа отражает полидисперсность препаратов ДНК, тогда как вторая – их гетерогенность.
Как отмечено выше, ДНК локализуется в ядрах, митохондриях и пластидах соматических клеток. Поэтому в зависимости от локализации ДНК различают ядерные (хромосомные) и экстраядерные (экстрахромосомные) детерминанты наследственности. Кроме того, известны транспозируемые генетические элементы (инсерционные последовательности и транспозоны).
ДНК выполняет разнообразные функции:
1)хранит генетическую (наследственную) информацию, записанную в виде последовательности нуклеотидов;
2)передает наследственную информацию из ядра в цитоплазму. Для этого с ДНК снимает копию и-РНК и переносит информацию к рибосомам – месту синтеза белка;
3)передает наследственную информацию от материнской клетки к дочерним клеткам, для чего перед делением клетки ДНК реплицируется.
ДНК – носитель генетической информации. Впервые понятие ген было сформулировано в 1941 г. Д. Бидлом и Э. Татумом: ген – это участок молекулы ДНК, несущий информацию об одном белке-ферменте. В настоящее время геном называют участок молекулы ДНК, кодирующий первичную структуру полипептида, и понятие о гене расширилось. Известны гены, кодирующие:
а) белки-ферменты; б) структурные белки;
в) т-РНК (много копий); г) р-РНК (много копий);
д) регуляторные (или функциональные) – включают и выключают другие гены;
е) гены-модуляторы – усиливают или подавляют проявление других генов.
132
Открытие принципа комплементарности дало ключ к познанию явлений наследственности и изменчивости. Затем было обнаружено, что молекула ДНК обладает уникальным свойством – способностью к самоудвоению, или редупликации. Под действием ферментов молекула ДНК частично раскручивается, и к каждой нити ДНК в соответствии с принципом комплементарности притягиваются свободные нуклеотиды, ранее синтезированные в цитоплазме. В результате из одной двуспиральной молекулы ДНК образуются две идентичные двуспиральные молекулы ДНК. При редупликации молекулы ДНК процесс расхождения материнских нитей и синтеза на них дочерних нитей происходит одновременно, поэтому в каждой из обеих дочерних молекул ДНК одна нить является материнской, а вторая синтезированной вновь. Такой способ удвоения называется
полуконсервативным.
Репликация состоит из большого числа последовательных этапов, которые включают узнавание точки репликации, расплетание родительского дуплекса, удержание его цепей на расстоянии друг от друга, инициацию синтеза новых дочерних цепей, их элонгацию, закручивание цепей в спираль и терминацию репликации. Каждому этапу соответствует свой фермент. Весь комплекс репликативных ферментов называют реплисомой, или ДНКретиказной системой. Единица репликации получила название репликона. ДНК ряда вирусов реплицируется по принципу «катящегося кольца».
В одной из цепей происходит разрыв, после чего к 3'-концу этой цепи присоединяются нуклеотиды. Разорванная родительская цепь удлиняется комплементарно другой цепи, вытесняя свой собственный 5'-конец. Затем по мере того, как вытесненная цепь сматывается с кольца, начинается ее репликация. После завершения синтеза новой дочерней цепи новообразованный линейный дуплекс отщепляется с помощью нуклеазы и появляется линейная вирусная молекула ДНК. Другая дочерняя цепь повторяет процесс репликации: ее 3'-конец удлиняется, а 5'-конец сматывается и служит матрицей для синтеза новой дочерней цепи. На такой матрице можно получить множество новых линейных дуплексов. ДНК бактерий представляет собой кольцевую двойную спираль. Репликация ДНК происходит при сохранении кольцевой формы. Репликация происходит обычно в двух направлениях, то есть образуются две репликативные вилки. Обе вилки появляются в одной точке и удаляются от нее одновременно в обоих направлениях, пока вновь не встретятся. В этом месте два синтезированных дочерних двухцепочных кольца разделяются, при этом каждое из них содержит одну материнскую и одну дочернюю цепь.
Скорость репликации у бактерий очень велика: у некоторых из них новая ДНК синтезируется со скоростью более 45000 нуклеотидных остатков в минуту из расчета на одну вилку.
Репликация у эукариот происходит намного сложнее, так как ДНК организована в нуклеосомы и находится в составе волокон хроматина. Репликация происходит тоже в двух направлениях, но вилки движутся в 10 раз медленнее, чем у бактерий. Кроме того, репликация ДНК у эукариот начинается одновременно во многих точках, и из каждой точки одновременно
133
в противоположных направлениях движутся две репликативные вилки, что резко ускоряет процесс репликации в целом.
Многочисленные исследования, особенно опыты А. Корнберга, Т. Корнберга, позволили установить молекулярный механизм процесса репликации. Были обнаружены три вида ДНК-полимераз (ДНК-полимераза I, II и III). Оказалось, что для действия ДНК-полимеразы требуется предшествующая ДНК. То есть одна из цепей предшествующей молекулы ДНК служит затравкой (или праймером). ДНК-полимераза последовательно добавляет нуклеотиды к 3'-концу одной из цепей, служащей затравкой и матрицей. Следовательно, синтез новой цепи ДНК происходит в направлении от 5' к 3'. В то же время было доказано, что ДНК-полимераза сама по себе без затравки не может начать синтез новой молекулы ДНК. Она способна лишь удлинять уже существующую цепь и только при наличии цепи, играющей роль матрицы. Нуклеотиды присоединяются к цепи-затравке в соответствии с нуклеотидной последовательностью цепи-матрицы по принципу комплементарности. Продуктом ДНК-полимеразной реакции является дуплекс с комплементарно спаренными основаниями. ДНК-полимеразе необходимы как цепь-затравка, так и свободная цепь-матрица. Следовательно, этот фермент не способен полностью осуществить репликацию целой молекулы ДНК, если она является двухцепочньм кольцом, одноцепочным кольцом или линейным дуплексом, в котором все основания спарены. Открытие трех видов ДНК-полимераз позволило понять, каким образом происходит удлинение цепи ДНК. А. Корнберг показал, что у бактерий за элонгацию цепи отвечает преимущественно ДНК-полимераза III. Работа этого фермента требует затравки и матрицы. Удлинение цепи происходит путем присоединения нуклеотидов к 3' -концу затравки (то есть цепь элонгируется в направлении от 5' к 3'). ДНК-полимераза I и ДНК-полимераза III обладают не только полимеразной активностью, но также 3'-5' и 5'-3' – экзонуклеазными активностями, то есть они, кроме удлинения цепи, способны отщеплять концевые нуклеотиды от любого конца цепи ДНК. Функции ДНК-полимеразы II остаются неизвестными.
Долгое время оставался нерешенным вопрос о присоединении нуклеотидов к 5'-концу. ДНК-полимеразы способны присоединять нуклеотиды лишь к 3'-концу, но в репликативной вилке находятся 3'-конец одной цепи и 5'-конец другой цепи, который тоже должен удлиняться. Оставался нерешенным и вопрос об изначальной затравке процесса репликации. Ответы на эти вопросы нашел Р. Оказаки, который обнаружил, что в процессе репликации ДНК у бактерий основная масса новообразовавшейся ДНК находится в виде кусков. Эти куски получили название фрагментов Оказаки.
Эти фрагменты являются короткими отрезками молекулы ДНК, образующимися в процессе прерывистой репликации. Затем эти фрагменты сшиваются друг с другом в единую цепь. Это открытие показало, что одна из цепей ДНК реплицируется непрерывно в направлении от 5' к 3' (в направлении движения вилки).
Эта цепь получила название ведущей цепи. Другая цепь синтезируется прерывисто с образованием фрагментов Оказаки за счет присоединения
134
нуклеотидов тоже к 3'-концу (в направлении, противоположном движению вилки). Затем фрагменты Оказаки сшиваются с помощью ферментов во вторую дочернюю цепь, получившую название запаздывающей цепи.
Было выяснено, что праймером (затравкой) для синтеза фрагментов Оказаки служат короткие отрезки РНК, комплиментарные матричной ДНК. Образование этой РНК-затравки осуществляется с помощью фермента примазы. РНК-праймер состоит из нескольких рибонуклеотидных остатков, к которым ДНК-полимераза III присоединяет множество дезоксирибонуклеотидных остатков. Образуется фрагмент Оказаки, комплиментарный соответствующему участку цепи-матрицы. Затем РНК-праймер удаляется с помощью ДНК-полимеразы I, которая обладает зкзонуклеазной активностью. По мере отщепления рибонуклеотидных звеньев, каждый из них замещается на соответствующий дезоксирибонуклеотид с помощью ДНК-полимеразы I. В качестве затравки используется 3'-конец предыдущего фрагмента Оказаки. Присоединение фрагмента Оказаки к цепи ДНК осуществляется с помощью фермента ДНК-лигазы.
Для репликации необходимо разделение цепей родительской двухцепочной ДНК. Этот процесс протекает с помощью ферментов хеликаз. Они расплетают короткие участки ДНК непосредственно перед репликативной вилкой.
Как только небольшой участок ДНК оказывается расплетенным, к каждой из цепей присоединяется несколько молекул ДНК-связывающего белка (ДСБ). Эти молекулы препятствуют образованию комплементарных пар и обратному воссоединению цепей. Во время репликации короткий участок молекулы ДНК вращается за счет кратковременного разрыва одной из цепей ДНК, который быстро восстанавливается после 1-2 оборотов. Восстановление разрывов происходит при участии фермента ДНК-гиразы.
Основной функцией ДНК является информационная. Порядок расположения нуклеотидов в молекуле ДНК определяет порядок расположения аминокислот в молекулах белков, то есть их первичную структуру. Набор белков определяет свойства организма (клетки). Информация о структуре белков записана в виде генов – участков молекулы ДНК. Молекулы ДНК не только хранят наследственную информацию, но также воспроизводят ее в процессе редупликации и передают по наследству в процессе деления клеток и размножения.
Прокариоты обычно содержат только одну копию ДНК, и почти всегда в каждой молекуле ДНК присутствует одна копия любого гена. В прокариотической ДНК встречаются регуляторные и сигнальные последовательности нуклеотидов и практически отсутствуют нетранслируемые участки. Организация генов в клетках эукариотических организмов более сложная. Оказалось, что в ДНК эукариот имеются фрагменты, присутствующие во множестве копий. Это высокоповторяющиеся последовательности. Кроме этих, имеются и умеренные повторы. Например, у мыши ДНК почти 10% состоит из коротких фрагментов, повторяющихся миллионы раз (высокоповторяющиеся последовательности), на 20% из фрагментов, повторяющихся порядка 100 раз (умеренные повторы) и лишь на 70% из уникальных (неповторяющихся) последовательностей. Число повторяющихся
135
последовательностей различно у разных видов, эти участки, по-видимому, не транслируются. В ДНК эукариот имеются также многочисленные палиндромы
– участки, в которых обнаруживаются обращенные последовательности с осевой симметрией второго порядка, напоминающие последовательности, узнаваемые рестриктирующими эндонуклеазами. Для сравнения, в литературе палиндромы – это слова или целые фразы, одинаково читаемые как слева направо, так и справа налево.
Среди палиндромов встречаются короткие и длинные. Короткие палиндромные последовательности выполняют функцию особых сигналов. Роль длинных палиндромов пока не установлена. Структурно же длинные палиндромные последовательности могут образовывать кресты путем спаривания оснований внутри каждой из двухцепочных петель.
Принято считать, что подавляющее большинство функционирующих генов – это уникальные последовательности. Однако имеются исключения. Например, гены, кодирующие структуру белков-гистонов и р-РНК, располагаются в виде тандемных повторов. Большинство умеренно повторяющихся последовательностей не транскрибируется, то есть они не функционируют в качестве матриц. По-видимому, повторяющиеся последовательности выполняют структурную или регуляторную функции. Локализованы эти последовательности в основном в прицентромерном гетерохроматине.
ДНК непосредственного участия в синтезе белков не принимает. В клетках человека (животных и растений) молекулы ДНК находятся в ядре и отделены ядерной мембраной от цитоплазмы, где происходит синтез белка. Информацию несет посредник – и-РНК, которая по принципу комплиментарности считывает (копирует) с ДНК информацию при участии фермента РНК-полимеразы. Переписывание информации происходит с одной нити ДНК и называется транскрипцией. Если в переписываемой нити ДНК стоит нуклеотид гуанин (Г), то фермент РНК-полимераза включает в РНК цитозин (Ц); если тимин (Т) – РНК-полимераза включает аденин (А); если стоит аденин (А), фермент включает урацил (У). По длине каждая из молекул и-РНК в сотни раз короче ДНК. Информационная РНК является копией не всей молекулы ДНК, а только ее части – одного гена, несущего информацию о структуре белка. Готовая и-РНК отходит от ДНК и направляется к месту синтеза белка.
Благодаря процессу транскрипции в клетке осуществляется передача информации от ДНК к белку по цепочке: ДНК – и-РНК-белок. Перевод информации с «языка» нуклеотидов на «язык» аминокислот осуществляется с помощью генетического кода.
Генетический код – это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в ДНК и и-РНК (табл. 3). Участок молекулы ДНК, состоящий из трех нуклеотидов, называется триплетом или кодоном. Каждая аминокислота шифруется более чем одним кодоном (от 2 до 6).
Свойства генетического кода:
136
1.Код триплетен – каждая из 20 аминокислот зашифрована последовательно расположенными тремя нуклеотидами. Из 4 нуклеотидов (так как существует 4 варианта азотистых оснований) можно создать 64 различные комбинации по 3 нуклеотида в каждом (4*4*4 = 64).
2.Код вырожден – каждая аминокислота шифруется более чем одним кодоном (от двух до шести), исключение составляют аминокислоты: метионин, который кодируется только триплетом АУГ, и триптофан – УГГ.
3.Код специфичен – каждый кодон шифрует только одну аминокислоту.
4.Код универсален – один триплет кодирует одну и ту же аминокислоту
увсех живых организмов.
5.Код неперекрываем – каждый нуклеотид входит лишь в какой-либо один триплет и переписывание информации происходит строго потриплетно.
6.Триплеты УАА, УАГ, УГА обозначают прекращение синтеза одной полипептидной цепи, так как к ним нет аминокислот. Они находятся в конце каждого гена.
|
|
|
|
|
|
Таблица 3. |
|
|
|
Генетический код |
|
|
|
||
|
|
|
|
|
|
|
|
Аминокислота |
|
Кодирующие триплеты – кодоны |
|
|
|||
Аланин |
ГЦУ |
ГЦЦ |
ГЦА |
ГЦГ |
|
|
|
Аргинин |
ЦГУ |
ЦГЦ |
ЦГА |
ЦГГ |
АГА |
|
АГГ |
Аспаргин |
ААУ |
ААЦ |
|
|
|
|
|
Аспаргиновая |
ГАУ |
ГАЦ |
|
|
|
|
|
кислота |
|
|
|
|
|
|
|
Валин |
ГУУ |
ГУЦ |
ГУА |
ГУГ |
|
|
|
Гистидин |
ЦАУ |
ЦАЦ |
|
|
|
|
|
Глицин |
ГГУ |
ГГЦ |
ГГА |
ГГГ |
|
|
|
Глутамин |
|
|
ЦАА |
ЦАГ |
|
|
|
Глутаминовая |
|
|
ГАА |
ГАГ |
|
|
|
кислота |
|
|
|
|
|
|
|
Изолейцин |
АУУ |
АУЦ |
АУА |
|
|
|
|
Лейцин |
ЦУУ |
ЦУЦ |
ЦУА |
ЦУГ |
УУА |
|
УУГ |
Лизин |
|
|
ААА |
ААГ |
|
|
|
Метионин |
|
|
|
АУГ |
|
|
|
Пролин |
ЦЦУ |
ЦЦЦ |
ЦЦА |
ЦЦГ |
|
|
|
Серин |
УЦУ |
УЦЦ |
УЦА |
УЦГ |
АГУ |
|
АГЦ |
Тирозин |
УАУ |
УАЦ |
|
|
|
|
|
Треонин |
УЦУ |
АЦЦ |
АЦА |
АЦА |
|
|
|
Триптофан |
|
|
|
УГГ |
|
|
|
Фениланин |
УУУ |
УУЦ |
|
|
|
|
|
Цистеин |
УГУ |
УГЦ |
|
|
|
|
|
В ДНК запрограммирована вся наследственная информация, и-РНК переписывает информацию с участка ДНК (гена) и переносит ее в цитоплазму на рибосому. У эукариот и-РНК еще незрелая, поэтому в ядре и при выходе из
137
него происходит её процессинг – дозревание (вырезание неинформативных участков и другие процессы), в результате чего РНК укорачивается. Дозревшая и-РНК переносит информацию о синтезе белка в рибосому. Информация закодирована в виде триплетов. Один триплет (кодон) кодирует место одной аминокислоты в белковой молекуле, а последовательность аминокислот в белковой молекуле. Перевод информации с и-РНК на последовательность аминокислот называется трансляцией. В и-РНК существуют триплеты: инициирующий АУГ (определяет начало синтеза белка) и терминирующие УАГ, УАА, УГА (заканчивают синтез белка).
Одномоментно в рибосоме помещается 2 триплета: один – в пептидильном, другой – в аминоацильном (аминокислотном) участке рибосомы. К аминоацильному участку во время синтеза белка подтягиваются аминокислоты, а в пептидильном находится пептид (полипептид).
Вцитоплазме клетки всегда имеется не менее 20 различных аминокислот
исоответствующих им т-РНК. С помощью специфических ферментов аминокислоты узнаются, активируются и присоединяются к т-РНК, которая переносит их к месту синтеза белка в рибосому. В рибосоме в и-РНК находится кодон, а у т-РНК есть антикодон.
Если в рибосоме на и-РНК будет триплет АУГ, то к нему подойдет т-РНК с комплементарным антикодоном УАЦ; если ГГГ – то т-РНК с антикодоном
ЦЦЦ. После этого между аминокислотой, находящейся в пептидильном участке, и аминокислотой, находящейся в аминоацильном участке, происходит образование пептидной связи. Данная реакция осуществляется на большой субъединице рибосомы. Затем т-РНК, находящаяся в пептидильном участке, вытесняется из него и «уходит» в цитоплазму за другой аминокислотой, а рибосома передвигается на следующий триплет (ААА), который будет в аминоацильном участке рибосомы, триплет же ГГГ окажется в пептидильном участке. Так происходит считывание информации. Когда рибосома окажется на терминирующем триплете, синтез белка заканчивается. Наращивание аминокислот в белковой молекуле в процессе синтеза белка называется элонгацией (удлинением). Синтез одной молекулы белка длится всего 3-4 с. Каждый этап синтеза катализируется соответствующими ферментами и снабжается энергией за счет расщепления АТФ.
После окончания синтеза белка и образования первичной структуры формируется вторичная, третичная, а иногда и четвертичная структура белка,
ион становится способным выполнять свои функции. Сходство и различие организмов определяется набором белков. Каждый вид имеет только ему присущий набор белков, то есть они являются основой видовой специфичности, а также обуславливают индивидуальную специфичность организмов. На Земле нет двух людей, у которых все белки были бы одинаковыми (за исключением монозиготных близнецов). ДНК каждой клетки несет в себе информацию не только о структурных белках, определяющих форму клетки, но и всех белках-ферментах, белках-гормонах и др. Практически все признаки клеток и организма в целом определяются белками. Таким образом, в ДНК заключена вся информация о структуре и деятельности клеток, о всех признаках каждой клетки и организма в целом. Эта информация называется наследственной.
138
Вклетках имеются системы репарации ДНК, устраняющие повреждения
вмолекуле ДНК Повреждения ДНК возникают при действии излучений, ряда химических веществ, приводящих к частичной местной денатурации молекулы. В поврежденных участках блокируется репликация ДНК. Известны три механизма, репарации ДНК: фотореактивация, эксциционная репарация и
пострепликативная репарация.
Явление фотореактивации – это восстановление биологической активности клеток или молекул ДНК после повреждения ультрафиолетовым излучением в результате последующего воздействия видимым светом. Фотореактивация может быть неферментативной и ферментативной. Первая заключается в мономеризации пиримидиновых димеров при действии УФизлучения с длиной волны 240 нм. Вторая связана с действием видимого света и участием специального фермента – дезоксирибопиримидинфотолиазы. Фермент в темноте образует комплекс с димерами пиримидиновых оснований (т.е. с разрушенными участками ДНК). На свету комплекс распадается с мономеризацией димеров. Происходит восстановление структуры молекулы на месте повреждения.
Эксцизионная репарация – это репарация по типу выщепления – замещения и связана с удалением поврежденного участка ДНК. Эксцизионная репарация – многоступенчатый процесс, состоящий в узнавании димера, инцизии (надрезания одной цепи ДНК вблизи димера), эксцизии (удалении димера), ресинтезе ДНК, восстановлении репарируемой цепи путем сшивания скелета молекулы. Каждый этап эксцизионной репарации осуществляется при участии ферментов.
Пострепликативная репарация (или рекомбинашионная репарация) – это способ быстрого восстановления нативной структуры дочерних молекул ДНК. При этом тиминовые димеры остаются в исходных родительских нитях. Разновидностью пострепликативой репарации является SOS-репараиия (или репарация, склонная к ошибкам). Она протекает весьма медленно, и восстановление первичной структуры ДНК происходит неточно. Эта репарация часто встречается в клетках с большим количеством мутаций.
Значение генетики для теории и практики медицины огромно.
Для создания гибридных, рекомбинантных молекул ДНК, а стало быть, и организмов с новыми признаками необходимо выделить ген из какого-либо организма или искусственно синтезировать его, клонировать (размножить) и перенести в другой организм. Инструментами генной инженерии являются ферменты: рестриктазы (разрезающие молекулы ДНК) и лигазы (сшивающие её). В качестве векторов-переносчиков используются вирусы.
Внастоящее время ученые разных стран работают над получением с помощью генной инженерии ряда других необходимых биологически активных веществ, вакцины против гепатита В, активатора профибринолизина (противосвертывающий препарат), интерлейкина-2 (иммуномодулятор) и др.
В клетки животных чужеродные гены вводят в виде отдельных молекул ДНК или в составе векторов-вирусов, способных вносить в геном клетки чужую ДНК. Обычно применяют два метода: 1) ДНК добавляют в среду инкубации клеток; 2) производят микроинъекции ДНК непосредственно в ядро (что более эффективно).
139
Тестовые задания к главе 6
1.Классификация живых организмов по способам питания подразделяется на:
1)автотрофные, гетеротрофные и миксотрофные;
2)автотрофные, когда питание осуществляется за счет использования неорганических веществ природы при построении организма;
3)гетеротрофные, когда питание осуществляется за счет использования низкомолекулярных органических соединений при построении организма.
2.Во время каких периодов происходил расцвет и вымирание крупных ящеров ?
1) юрский период (195 млн. лет назад); 2) меловой период (136 млн. лет назад); 3) палеогеновый (42 млн. лет назад).
3.Вещество, содержащееся в ядрах клеток (ДНК – дезоксирибонуклеиновая кислота) является:
1)носителем генетической информации;
2)не является носителем генетической информации;
3)регулирует обмен веществ в клетке.
140
Глава 7 ВАРИАЦИИ ГЕОМАГНИТНОГО ПОЛЯ
Рассматирвается структура геомагнитного поля, вариации гомагнитного поля под влиянием гелиомагнитных возмущений и влияния геомагнитных бурь на гео- и биофизические объекты Земли.
История изучения геомагнитного поля
О существовании магнетизма человечеству было известно с VIII века до н.э. Свойство камня притягивать к себе железо описывается в различных сочинениях греческих ученых. Так, например, об этом факте упоминается в работах Фалеса Милетского (640-546 гг. до н.э.).
В1187 г. был изобретен компас, которым пользовались моряки во время плавания. В 1269 г. П. Перегин излагает все известные в его время свойство магнита и дает первые названия полюсов магнита. Конец стреки, указывающий на север П.Перегин предложил назвать северным полюсом, а противоположный – южным. Флавио Джойя, живший в начале ХIV века усовершенствовал компас. Он посадил компасную стрелку на острие и снабдил её бумажным кругом, разделённым на 32 части (румбов) под названием «роза ветров». Новым в развитии науки о земном магнитизме послужило открытие магнитного склонения Х.Колумбом во время его плавания из Европы в Америку 13-17 сентября 1492 г. при пересечении Атлантического океана. Им было замечено отклонение стрелки от истинного меридиана и изменение оклонения от места к месту. Эти факты послужили началом измерения на земном шаре магнитного склонения. В 1544 году Г.Гартман открыл магнитное наклонение. В письме графу А.Прусскому он сообщил, что магнитная стрелка не только отклоняется к востоку приблизительно на 9 градусов, но и наклоняется к низу. Так был открыт полный вектор магнитного поля Земли, который характеризуется двумя угловыми величинами (склонением и наклонением).
В1682 году Г.Ташаром, наблюдая склонение в г. Луво в Сиаме обнаружил его суточные коллебания. Позднейшие наблюдения подтвердили существенные вариации наклонения и склонения. Это послужило организации систематических наблюдений на земном шаре за элементами земного магнетизма.
ВРоссиии первым, кто начал заниматься изучением земного магнетизма был М.В. Ломоносов. В 1759 году в своём труде «Рассуждения о большой точности морского пути» им впервые была высказана мысль, что земной шар имея общее магнитное поле состоит из разнородно-намагниченных блоков и отдельных его частей, которые в совокупности образуют неоднороднонамагниченный шар, чем и объясняется неодинаковые значения склонения в разных частях земного шара.
И.М. Симонов в 1835 и К. Гаусс в 1839 году дали теоретическое обоснование метода измерения горизонтальной составляющей геомагнитного поля.
По инициативе академика Г.Вильда в 1870 году возобновились вариационные наблюдения магнитного поля в Петербурге. В 1878 году они
141
были перенесены в г.Павловск, где была создана первоклассная магнитная абсерватория.
В настоящее время геомагнитные абсеврватории имеются во многих странах мира. Приведем основные теоретические положения земного магнетизма.
7.1.Структура и свойства геомагнитного поля
Давно экспериментально установлено, что земной шар обладает магнитным полем. Наибольшая напряженность постоянного магнитного поля (модуль полного вектора) на поверхности Земли достигает 0,7 Э (эрстеда). Если считать, что этот момент сосредоточен в магните, расположенном на расстоянии примерно 400 км от центра Земли, то направление оси намагничения этого магнита составит угол 11,5 с осью вращения Земли. Создаваемое им поле называют дипольным.
Рис. 36. Элементы земного магнетизма.
Магнитное поле Земли (как векторное поле) характеризуется вектором напряженности (Нт), который в направлении магнитного меридиана можно разложить на две составляющие: горизонтальную Н, действующую в горизонтальной плоскости, и вертикальную Z, перпендикулярную к Н. Горизонтальную составляющую в свою очередь можно разложить на силу, направленную вдоль географического меридиана, так называемую северную составляющую X, и силу, перпендикулярную к меридиану, – восточную составляющую У. Стрелка компаса в каждой точке Земли направлена вдоль магнитного меридиана, т.е. вдоль условной плоскости земной поверхности, совпадающей с направлением ГМП.
Полный вектор магнитного поля Земли наряду с изменением абсолютной величины претерпевает изменения в пространстве. Положение вектора в пространстве может быть характеризовано двумя угловыми величинами – склонением и наклонением. Магнитное склонение D является углом в горизонтальной плоскости, т.е. углом между географическим меридианом (линия север – юг) и магнитным меридианом данного места на Земле. Магнитное наклонение – угол между горизонтальной плоскостью и

142
направлением напряженности полного вектора ГМП (рис. 36). Полный вектор ГМП и его элементы связаны между собой следующими соотношениями:
|
|
|
|
|
|
Z |
|
Y |
|
|
T |
H 2 Z 2 , |
tg I |
, tg D |
, |
||||||
H |
X |
|||||||||
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
||
T H sec I , H |
|
X 2 |
Y 2 , X |
|
H cos D, |
Y |
H sin D. |
|||
Полный вектор и его вертикальная и горизонтальная составляющие измеряются в эрстедах и гаммах (1Э = 105 гамм), склонение и наклонение – в угловых градусах и минутах. В течение магнитоспокойных и магнитовозмущенных дней элементы магнитного поля претерпевают различные изменения.
Геомагнитное поле различно по своей структуре и динамическим свойствам.
С одной стороны, ГМП можно подразделить по степени изменяемости, по характеру динамики на постоянное (основное) и переменное поле. Постоянным называется такое магнитное поле, которое освобождено от всех типов вариаций с периодом до года. Правда, и в этом случае оно не остается строго постоянным, так как подвержено вековым колебаниям. Во всяком случае, период изменения постоянного магнитного поля Земли очень большой и составляет многие сотни лет.
Переменное магнитное поле Земли представляет собой изменения поля с различными периодами внутри года (от долей секунды до месяцев).
Сдругой стороны, если рассматривать структуру или причины, лежащие
воснове возникновения ГМП, то ГМП можно подразделить на поле однородного намагничения, материковое поле, аномальное, внешнее и поле вариаций.
По классификации Б.М. Яновского, ГМП является суммой нескольких полей: Но – поля, создаваемого однородной намагниченностью земного шара; Нт – поля, создаваемого неоднородностью глубоких слоев земного шара, материкового поля; На – поля, обусловленного различной намагниченностью верхних частей коры, аномального поля; Не – межпланетное магнитное поле;
Н– поля вариаций, вызванного импульсивно-волновым характером космической среды. Итак, полное поле
НТ = Но+Нт+Не+Нц+ Н.
Аномальное поле, т.е. искаженное ГМП, считают состоящим из полей трех видов. Материковые аномалии, площадь которых соизмерима с континентами. Причина материковых аномалий однозначно не установлена. Высказаны гипотезы о том, что большую роль в их возникновении играют мощные вихревые токи. Б.М. Яновский указывает, что таких материковых аномалий насчитывается шесть, причем наиболее интенсивной является Восточно-Азиатская материковая аномалия.
Принято считать, что там, где магнитная ось пересекает поверхность Земли, располагаются геомагнитные полюса. Эти полюса не совпадают с географическими и отстоят от соответствующих географических полюсов – северный геомагнитный полюс на 800 км, а южный геомагнитный полюс на 1000 км. Координаты современного северного геомагнитного полюса
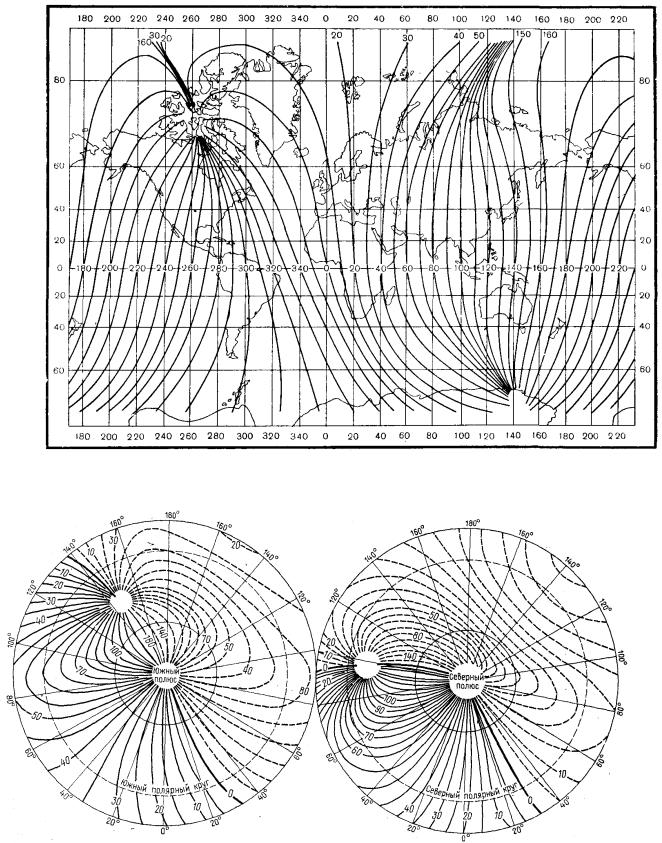
143
(р=78°30' с. ш. и A,=68°48' з. д., а южного геомагнитного полюса (p=78°30/ ю.
ш. и К =111°30' в. д.
Силовые линии земного магнитного поля выходят приблизительно из центра Земли (поскольку магнитный диполь расположен эксцентрично) через южное полушарие и, огибая Землю, вновь направляются к центру, но через северное полушарие (рис. 37, 38).
Рис. 37. Карта магнитных меридианов.
Рис. 38. Изогоны в районах географических полюсов.
144
Сплошные кривые – положительные значения склонения, пунктирные – отрицательные, жирные – нулевые.
В северном полушарии расположен южный геомагнитный полюс, в южном полушарии – северный геомагнитный полюс, но в обиходной речи его называют по аналогии с географическим – южным, что неправильно.
Рис. 39. Вертикальная составляющая поля мировых аномалий для эпохи
1942 г. (Вестин, 1947).
Стрелками показана горизонтальная составляющая; значения приведены в миллиэрстедах, отрицательным значениям соответствуют пунктирные изодинамы.
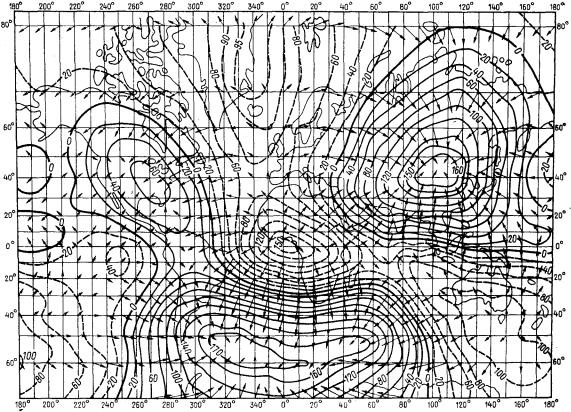
Кдругому виду относятся региональные аномалии, занимающие площадь
вдесятки или сотни квадратных километров (рис. 39). Такие аномалии обусловлены тектоническими и структурными причинами. Не обязательно региональные аномалии отличаются высокой намагниченностью (как, например, Курская магнитная аномалия), но всегда констатируется отличие магнитных свойств пород данного района от других. Наконец, третий вид аномалий – локальные; возникают там, где магнитные породы залегают непосредственно у поверхности Земли. Площадь локальных аномалий небольшая, обычно не превышает десять – двадцать квадратных километров.
На северном геомагнитном полюсе напряженность поля составляет 0,6Э, на южном 0,7Э, а на магнитном экваторе 0,35Э. Следует заметить, что приводимые выше координаты геомагнитных полюсов относятся только к условно однородно намагниченному шару идеальной формы. На самом же деле в силу различных причин структура геомагнитного поля значительно сложнее. Поэтому реальные, так называемые магнитные полюса, где магнитная стрелка занимает вертикальное положение, имеют координаты
145
(эпоха 1970 года): =75° с.ш., = 101° з.д. – северный полюс и =70° ю.ш., = 140° в.д.– южный полюс.
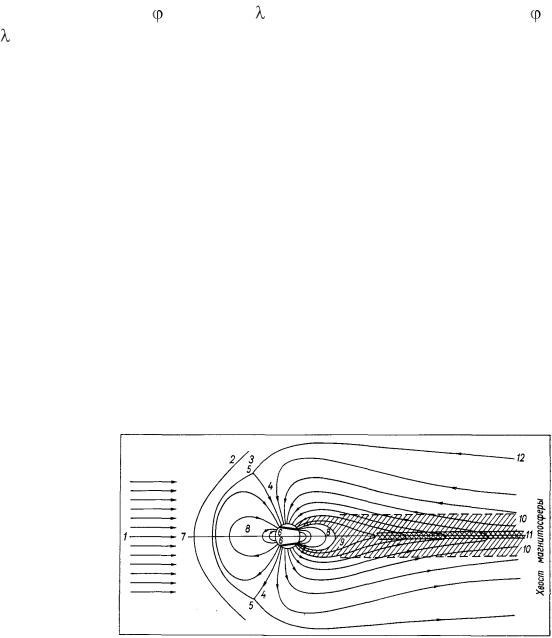
Исследования Ю.И. Витинского, А.И. Оль, Б.И. Сазонова показали что, наличие в межпланетном пространстве солнечного ветра, состоящего из квазинейтральной плазмы, существенно изменяет распределение геомагнитного поля в пространстве. Солнечный ветер весьма эффективно взаимодействует с магнитным полем Земли, так как проводимость его очень высока. Вследствие этого при набегании солнечного ветра на геомагнитное поле в проводящей плазме индуцируются электрические поля, возникает система электрических токов, магнитное поле которых накладывается на геомагнитное поле и сильно его искажает (рис. 40). Это индуцированное магнитное поле уничтожает магнитное поле Земли внутри потока солнечного ветра и усиливает геомагнитное поле перед фронтом солнечного ветра. В результате этого магнитное поле Земли как бы вырезает в потоке солнечного ветра некоторую полость, имеющую в стороне, обращенной к Солнцу, приблизительно полусферическую форму, а с ночной стороны эта полость вытягивается в виде очень длинного цилиндра, имеющего в длину сотни тысяч километров. Эта полость, ограничивающая геомагнитное поле конечной областью пространства, получила название магнитосферы.
Рис. 40. Меридиональное сечение магнитосферы Земли.
1 – солнечный ветер, 2 – отошедшая ударная волна, 3 – переходная область, 4 – дневные полярные каспы, 5 – нейтральные точки, 6 – авроральные овалы, 7 – геомагнитный экватор, 8 – область захваченной радиации (внутренняя магнитосфера), 9 – ночной касп, 10 – плазменный слой, 11 – нейтральный слой, 12 – магнитопауза.
Так как скорость солнечного ветра сверхзвуковая, то при обтекании геомагнитного поля солнечным ветром возникает стационарная «отошедшая ударная волна», т.е. граница, отделяющая области пространства с существенно различными параметрами плазмы и магнитного поля.
На некотором расстоянии от ударной волны расположена магнитопауза, т.е. граница полости, содержащей геомагнитное поле. Между ударной волной и магнитопаузой находится переходная область, характеризующаяся возрастанием температуры плазмы солнечного ветра и изменением направления и напряженности магнитного поля, вмороженного в эту плазму –
146
поле несколько усиливается и становится тангенциальным по отношению к магнитопаузе.
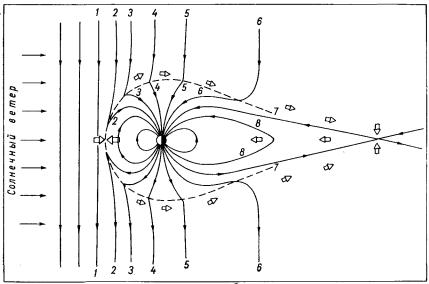
Солнечный ветер благодаря магнитогидродинамическому взаимодействию с геомагнитным полем «сдувает» часть силовых линий с дневной стороны на ночную, в хвост магнитосферы (рис. 41). При этом на дневной стороне северной и южной полярных шапок Земли образуются воронкообразные области, отделяющие замкнутые силовые линии на дневной стороне магнитосферы от разомкнутых силовых линий, уходящих в хвост. Эти области называются дневными полярными каспами (от английского сиар
– выступ, мыс). Плазма переходного слоя может сравнительно легко проникать к Земле вдоль силовых линий, ограничивающих каспы. Внутри каспов геомагнитное поле ослаблено и область схождения силовых линий, образующих касп, условно называется нейтральной точкой.
Рис. 41. Схема пересоединения межпланетного магнитного поля, направленного на юг, с геомагнитным полем и переноса силовых линий на ночную сторону магнитосферы.
1 – положение некоторой силовой линии до пересоединения, 2-8 – последовательные положения этой линии после пересоединения; светлые стрелки – направление движения плазмы.
Геомагнитные силовые линии, образующие хвост (шлейф) магнитосферы, имеют противоположное направление по обе стороны от лежащего посередине нейтрального слоя, где напряженность магнитного поля очень мала. В этом слое происходит замыкание части противоположно направленных силовых линий, т.е. магнитное поле в этом слое в основном перпендикулярно плоскости геомагнитного экватора.
Процессы в нейтральном слое приводят к возникновению так называемых авроральных явлений, разыгрывающихся в двух овальных зонах, расположенных вблизи северного и южного геомагнитных полюсов, получивших название авроральных овалов. К авроральным явлениям относятся полярные магнитные бури, или суббури, полярные сияния, ионосферные возмущения. Все эти явления возникают в результате
147
грандиозного бурного процесса, развивающегося в магнитосфере (магнитосферного возмущения). Причиной магнитосферного возмущения является либо усиление солнечного ветра, иногда имеющее характер ударной волны, распространяющейся от Солнца вдоль потока солнечного ветра, либо приход к границе магнитосферы вмороженного в движущуюся плазму магнитного поля, имеющего компоненту, перпендикулярную плоскости эклиптики и направленную на южный полюс эклиптики, либо совместное действие обеих причин. Наиболее важную роль играет приход направленного к югу межпланетного магнитного поля. Силовые линии такого поля могут пересоединяться с силовыми линиями геомагнитного поля.
Когда силовые линии входят в хвост магнитосферы, они сближаются, и напряженность магнитного поля в хвосте возрастает. Это приводит к возникновению неустойчивого состояния магнитного поля в хвосте. Вследствие какого-либо внешнего импульса может начаться бурное пересоединение противоположно направленных силовых линий через нейтральный слой. В сильно искривленных силовых линиях возникают значительные максвелловские напряжения, линии стремятся сократиться и перемещаются по направлению к Земле, увлекая за собой плазму, заполняющую плазменный слой хвоста. Вследствие возникновения плазменных неустойчивостей в процессе пересоединения силовых линий энергия магнитного поля, накопленная в хвосте, переходит в кинетическую энергию частиц. При этом возрастает тепловая энергия частиц, и, кроме того, частицы приобретают скорость направленного движения. Частицы перемещаются сперва вдоль хвоста, а затем, огибая замкнутые силовые линии внутренней (или замкнутой) магнитосферы, они устремляются вдоль границы между замкнутыми и разомкнутыми силовыми линиями и приходят в авроральные овалы. Кроме того, частицы испытывают добавочное ускорение, перемещаясь из области более слабого магнитного поля хвоста в более сильное поле вблизи замкнутой магнитосферы. Наиболее ускоренные частицы прорываются в замкнутую магнитосферу и образуют там кольцевой ток, ответственный за падение геомагнитного поля во время главной фазы магнитной бури.
Ускоренные частицы, пришедшие из хвоста магнитосферы в авроральные овалы (их называют авроральными частицами), увеличивают ионизацию ионосферы, что приводит к усиленному поглощению радиоволн в нижних слоях ионосферы и к значительному увеличению проводимости ионосферы, вызывающему (при наличии электрического поля) ионосферные токи, магнитные поля которых регистрируются на земной поверхности. Так возникают возмущения в нижних слоях ионосферы и магнитные бури. Одновременно в верхней ионосфере наблюдаются возмущения другого типа, характеризующиеся падением концентрации заряженных частиц и увеличением высоты верхней ионосферы. Эти возмущения вызываются, повидимому, электромагнитными дрейфами и разогревом верхней атмосферы во время вторжения авроральных частиц. Наконец, авроральные частицы, сталкиваясь с атомами и молекулами воздуха, возбуждают их свечение, т.е. полярные сияния.