

Полезные материалы за все 6 курсов / Учебники, методички, pdf / Ферменты. Методичка. БХ-1
.pdfКФ 1.9. Действующие на гем как донор
Подкласс 1.9 представляет собой оксидоредуктазы, действующие на гемсодержащие соединения в качестве донора. Представителем является III комплекс дыхательной цепи – цитохром с оксидаза (КФ 1.9.3.1), находящаяся во внутренней мембране митохондрий и катализирующая окисление железа в цитохроме с.
4цитохром c (Fe2+) + 4Н+ + О2 4 цитохром c (Fe3+) + 2Н2О
Всостав цитохром с оксидазы входят два цитохрома а и а3 (гемсодержащие белки, осуществляют перенос электронов), а также три иона Сu+. Восстановление кислорода до воды сопровождается переносом 4 протонов в межмембранное пространство из матрикса митохондрий.
КФ 1.10. Действующие дифенолы и родственные вещества как доноры
В результате реакции, катализируемой сукцинатдегидрогеназой, образуется убихинол, который представляет собой субстрат для фермента другого подкласса оксидоредуктаз (действующих на дифенолы и родственные группы доноров) – убихинон-цитохром с редуктазы (КФ 1.10.2.2).
|
|
|
|
OH |
|
|
|
|
|
O |
|||||
H C |
|
O |
|
|
|
CH3 |
H C |
|
O |
|
|
|
|
CH3 |
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
||||||||||
3 |
|
|
|
|
|
|
3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
+ 2 цитохром c (Fe3+) |
|
|
|
|
|
|
|
|
+ 2 цитохром c (Fe2+) |
H3C |
|
O |
|
|
|
H3C |
|
O |
|
|
|
|
|||
|
|
|
|
R |
|
|
|
|
|
R |
|||||
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
OH |
|
|
|
|
|
O |
|||||
убихин о л |
убихин о н |
|
Этот фермент представляет собой II комплекс дыхательной цепи. Содержит цитохромы b-562, b-566 и c1 (гемсодержащие белки, осуществляющие перенос электронов). В зависимости от условий убихинон-цитохром с редуктаза переносит 2 или 4 протона через внутреннюю митохондриальную мембрану из матрикса митохондрий в межмембранное пространство митохондрий.
КФ 1.11. Действующие на пероксид как акцептор
Подкласс 1.11 оксидоредуктаз (гидропероксидазы) включает в себя ферменты, действующие на пероксид водорода в качестве акцептора электронов. В качестве примеров можно привести каталазу (КФ 1.11.1.6) и глутатионпероксидазу (КФ 1.11.1.9).
Каталаза представляет собой гемопротеин, субстратом для которого служит не только пероксид водорода, но и органические перекиси, а также этанол:
Н2О2 Н2О+ О2 Глутатионпероксидаза также катализирует обезвреживание пероксида
водорода и органических перекисей. Глутатион (GSH), участвующий в реакции, представляет собой трипептид и состоит из аминокислот: глутаминовой кислоты
11
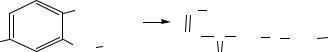
цистеина и глицина. В состав фермента входит селен. SH-группы цистеина непосредственно принимают участие в химической реакции. В результате образуется продукт - дисульфид глутатиона (GSSG).
2 GSH + H2O2 GSSG + 2 H2O
Каталаза и глутатионпероксидаза являются важными компонентами антиоксидантной системы организма.
КФ 1.13. Действующие на один донор с включением молекулярного кислорода (оксигеназы)
Примером данного подкласса является фермент, участвующий в катаболизме тирозина гомогентизат-1,2-диоксигеназа (КФ 1.13.11.5). Фермент катализирует окисление гомогентизиновой кислоты до малеинацетоацетата:
OH |
CH COOH |
+O2
HO |
|
CH |
C |
CH2 C |
CH2 COOH |
||
CH2 |
COOH |
O |
|
|
|
|
|
|
|
|
O |
|
|||
|
ãî ì î ãåí òè çàò |
|
4-м алеи н ацето ацетат |
||||
Для проявления своей активности гомогентизат-1,2-диоксигеназа требует наличия Fe2+. Наследственный дефект фермента приводит к развитию алкаптонурии.
КФ 1.14. Действующие на пару доноров с включением или восстановлением молекулярного кислорода
Неспецифическая монооксигеназа (КФ 1.14.14.1) имеет несколько названий, наиболее распространенное из них цитохром Р450. Суперсемейство цитохрома Р450 локализуется в мембранах эндоплазматического ретикулума и митохондрий. Фермент является одновременно гемо- и флавопротеидом, причѐм флавиновый нуклеотид выступает в качестве второго донора электрона.
RH + восстановленный флавопротеин + O2
ROH + окисленный флавопротеин + H2O
Для обеспечения функционирования цитохрома Р450 необходимо восстановление флавопротеина, которое осуществляется НАДФН- цитохромаР450-редуктазой. Донор электронов – НАДФН. Цитохром Р450 и НАДФН-цитохромР450-редуктаза составляют микросомную монооксигеназную систему. В этом случае, общую реакцию можно записать следующим образом:
RH + НАДФН + О2 ROH + Н2О + НАДФ+
Суперсемейство изоформ цитохрома Р450 катализируют реакции гидроксилирования, N-окисления, окислительного дезаминирования,
12

сульфоокисления, N-, S- и O-деалкилирования, десульфирования, эпоксидирования, реакции восстановления нитро- и азогрупп. Субстратами для фермента являются эндогенные метаболиты (стероиды, жирные кислоты, простагландины и др.), а также многие чужеродные вещества (ксенобиотики), включая лекарственные вещества.
Образование оксигрупп повышает гидрофильность исходно липофильных эндогенных метаболитов и ксенобиотиков, что облегчает их последующее выведение из организма.
Другим примером фермента, относящегося к данному подклассу является тирозин-3-монооксигеназа (КФ 1.14.16.2), катализирующая образование диоксифенилаланина (ДОФА) – предшественника в синтезе катехоламинов. В активном центре фермента находится ион железа (II). В качестве кофермента выступает восстановленный птеридин – тетрагидробиоптеридин (Н4БП), который в результате реакции окисляется до дигидроптеридина (Н2БП).
OH |
|
|
|
|
OH |
|
||||
|
|
|
H4ÁÏ + O2 |
|
|
|
|
|
OH |
|
|
|
|
|
|
|
|
|
|
||
|
|
+ |
|
|
|
|
|
+ H2ÁÏ |
+ H2O |
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
||
|
CH2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
CH |
|
||||
|
|
|
|
|
|
|
2 |
|
|
|
|
CH NH2 |
|
|
|
|
CH NH |
|
|||
|
|
|
|
|
|
|
2 |
|
||
COOH |
|
|
|
|
COOH |
|
||||
|
|
|
|
|
|
|
|
|||
òè ðî çè í |
|
|
|
ÄÎ ÔÀ |
|
|||||
В данной реакции один атом молекулярного кислорода присоединяется к продукту реакции, а другой атом используется для образования воды. Такие ферменты называют монооксигеназами.
Диоксигеназы используют оба атома кислорода для образования продукта реакции.
КФ 1.15. Действующие на супероксид как акцептор
Подкласс 1.15 содержит единственный фермент – супероксиддисмутазу (КФ 1.15.1.1), которая представляет собой металлопротеин, содержащий в своѐм составе ионы (Cu2+, Zn2+, Mn2+, Fe2+). Супероксидный радикал О2- является побочным продуктом многих реакций (например, окислительного дезаминирования биогенных аминов). Кроме того, О2- активно образуется НАДФ-оксидазой в активированных нейтрофилах. Этот радикал обладает сильной разрушительной силой, повреждает не только клеточные мембраны, но и белки, нуклеиновые кислоты. Именно супероксиддисмутаза осуществляет
детоксикацию О2-. .
О2-. + О2-. + 2Н+ Н2О2 + О2
Продукт реакции Н2О2 обезвреживается впоследствии под действием каталазы и различных пероксидаз.
13
КФ 2. ТРАНСФЕРАЗЫ
Трансферазы – класс ферментов, катализирующих реакции переноса каких-либо функциональных групп с одного субстрата на другой, либо реакции обмена субстратов между собой этими группами. В этих реакциях участвуют два субстрата:
S-G + S' S'-G + S
Трансферазы занимают ключевые позиции в энергетическом и пластическом обменах веществ. В зависимости от химической природы переносимых групп трансферазы подразделяют на 9 подклассов.
Номер |
Название подкласса |
КФ 2.1 |
Переносящие одноуглеродные группы |
КФ 2.2 |
Переносящие альдегидные, кетогруппы |
КФ 2.3 |
Ацилтрансферазы |
КФ 2.4 |
Гликозилтрансферазы |
КФ 2.5 |
Переносящие алкил- и арилгруппы, отличные от метильной |
|
группы |
КФ 2.6 |
Переносящие азотсодержащие группы |
КФ 2.7 |
Переносящие фосфорсодержащие группы |
КФ 2.8 |
Переносящие серусодержащие группы |
КФ 2.9 |
Переносящие селенсодержащие группы |
Обычно перенос химических групп, катализируемый трансферазами, осуществляется с помощью коферментов (пиридоксальфосфат, биотин и др.), образующих ковалентные промежуточные соединения с субстратом. В этом случае донором является кофермент, присоединяющий группу подлежащую переносу.
Трансферазы весьма разнообразны по механизму катализа, специфичности к субстратам, рН-зависимости. Молекулы многих трансфераз имеют четвертичную структуру. Некоторые трансферазы выполняют регуляторную функцию в клеточном метаболизме.
КФ 2.1. Переносящие одноуглеродные группы
Трансферазы, входящие в подкласс 2.1. переносят одноуглеродные группы (-CH3, -COOH, -CH2OH и т. д.). Эти ферменты участвуют в обмене аминокислот, адреналина, креатина, нуклеотидов и др. Кроме того, реакции метилирования необходимы для обезвреживания собственных метаболитов (адреналин), а также лекарственных веществ и ксенобиотиков. Донорами одноуглеродных остатков служат активная форма незаменимой аминокислоты метионина – S- аденозилметионин, активные формы фолиевой кислоты (формил-Н4-фолат, метилен-Н4-фолат).
В качестве примера приведѐм реакцию обезвреживания адреналина, которая катализируется цитоплазматическим ферментом катехол-О- метилтрансферазой (КФ 2.1.1.6):
14

HO |
H |
HO |
H |
|
N |
||
|
|
N |
|
|
|
|
|
|
|
|
CH3 |
SAM |
|
|||||
|
|
|
|
|
|
|
|
|
CH3 |
||
|
|
|
|
OH |
SAG |
|
|
|
|||
|
|
|
|
|
|
|
OCH3 |
||||
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
||
|
|
OH |
|
|
|
OH |
|||||
|
|
|
|
|
|
|
|
||||
АДРЕН АЛИН |
|
М ЕТИЛАДРЕН АЛИН |
|||||||||
Донором одноуглеродного остатка служит S-аденозилметионин (SAM), который в ходе реакции превращается в S-аденозилгомоцистеин (SAG).
КФ 2.2. Переносящие альдегидные, кетогруппы
Важными ферментами, относящимися к этому подклассу, являются транскетолазы и трансальдолазы пентозофосфатного пути превращения глюкозы. В этом процессе образуется не только важный углевод – рибозо-5-фосфат, но и восстановленные коферменты НАДФН, являющиеся донорами электронов в пластическом обмене. Транскетолаза (КФ 2.2.1.1) в качестве кофермента содержит тиаминпирофосфат и катализирует реакцию:
седогептулозо-7-фосфат + D-глицеральдегид-3-фосфат D-рибозо-5-фосфат + D-ксилулозо-5-фосфат
Трансальдолаза (КФ 2.2.1.2) также участвует во взаимопревращениях фосфорилированных моносахаров:
седогептулозо-7-фосфат + D-глицеральдегид-3-фосфат D-эритрозо-4-фосфат + D-фруктозо-6-фосфат
КФ 2.3. Ацилтрансферазы
Ферменты, относящиеся к данному подклассу, переносят ацильные остатки. В клетках животных в транспорте жирных кислот через мембрану митохондрий, где они окисляются в результате -окисления, принимает участие фермент карнитин-пальмитоилтрансфераза (КФ 2.3.1.21). Для реакции необходим карнитин, при участии которого жирные кислоты транспортируются через мембрану митохондрий:
АцилSКоА + карнитин ацилкарнитин + НSКоА
Фермент, участвующий в биотрансформации лекарственных веществ ариламин-N-трансфераза (КФ 2.3.1.5) катализирует реакцию переноса ацетатного остатка с ацетилSKoA на метаболизируемое вещество:
15

|
|
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
H2N |
|
S |
|
|
|
|
|
|
NH2 + CH3 C |
|
SKoA |
|
|
|
|
|
|
|
|
|
|
|
|
NH C CH3 + HSKoA |
|||||
|
|
|
|
|
|
|
|
|
|
H2N |
|
S |
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
O |
|
|
O |
|
|
|
|
O |
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
сульф ан илам ид ацетил-SKoA |
|
N-ацетилсульф ан илам ид |
|||||||||||||||||||||||||
γ-Глутамилтрансфераза (КФ 2.3.2.2) также входит в 2.3. подкласс трансфераз:
(5-L-глутамил)-пептид + аминокислота пептид + 5-L-глутамил-аминокислота
(5-L-глутамил)-пептид представляет собой трипептид – глутатион. Фермент принимает участие в транспорте аминокислот через цитоплазматическую мембрану в клетки. Определение активности γ- глутамилтрансферазы в сыворотке используется в лабораторной диагностике.
КФ 2.4. Гликозилтрансферазы
Ферменты, относящиеся к данному подклассу, переносят гликозильные остатки. Как правило, донором углеводного остатка служит уридиндифосфатмоносахарид (УДФ-глюкуронил, УДФ-глюкоза и др.). Например, непосредственным донором глюкозных остатков в синтезе гликогена служит уридиндифосфатглюкоза (УДФ-глюкоза) – продукт взаимодействия глюкозо-1- фосфата и УТФ. Фермент гликогенсинтаза (КФ 2.4.1.11) катализирует реакцию:
УДФ-глюкоза + (глюкоза)n УДФ + (глюкоза)n+1
Гликогенсинтаза из тканей животных представляет собой комплекс каталитической субъединицы и белка гликогенина. качестве затравки гликогенсинтазе небходим гликозилированный гликогенин.
К гликозилтрансферазам также относят гликогенфосфорилазу (фермент, катализирующий распад гликогена).
Фермент с широкой субстратной специфичностью УДФглюкуронилтрансфераза (КФ 2.4.1.17) участвует в обезвреживании метаболитов организма, лекарственных веществ и ксенобиотиков:
акцептор + УДФ-глюкуроновая кислота акцептор β-D-глюкуронид + УДФ
УДФ-глюкуронилтрансферазы составляют семейство ферментов с широким кругом субстратов, включая фенолы, спирты, амины и жирные кислоты. Фермент имеет важное значение в катаболизме гема, катализирует образование глюкуронидов билирубина.
КФ 2.6. Переносящие азотсодержащие группы
Очень важный подкласс трансфераз – ферменты, переносящие аминогруппы – аминотрансферазы. Эти ферменты играют огромную роль в
16

обмене аминокислот. Реакция переаминирования необходима для синтеза заменимых аминокислот, а также в катаболизме аминокислот. Кроме того, определение их активности в сыворотке крови используют в диагностике ряда заболеваний. Важное диагностическое значение имеют два фермента – аспартатаминотрансфераза (КФ 2.6.1.1) и аланинаминотрансфераза (КФ 2.6.1.2). Реакция, катализируемая аспартатаминотрансферазой представлена ниже:
КФ 2.7. Переносящие фосфорсодержащие группы
Чрезвычайно важный подкласс – трансферазы, переносящие остатки фосфорной кислоты – фосфотрансферазы или киназы. Без этих ферментов не могут начаться очень многие метаболические циклы в организме. Они переносят остаток фосфорной кислоты от аденозинтрифосфорной кислоты (АТФ) на какойлибо субстрат. Тем самым этот субстрат активируется и дальше способен вступать в реакции. Гексокиназа (КФ 2.7.1.1) и глюкокиназа (КФ 2.7.1.2) катализируют фосфорилирование глюкозы по шестому атому углерода:
Глюкоза + АТФ Глюкозо-6-Ф + АДФ
Реакции фосфорилирования белковых молекул по остаткам гидроксильных групп (радикалы серина, треонина и тирозина) необходимы для изменения функциональной активности белковой молекулы. При этом для некоторых ферментов фосфорилированная форма активна (гликогенфосфорилаза, липаза), тогда как для других, наоборот, фермент активен в дефосфорилированной форме (гликогенсинтетаза). Реакция фосфорилирования белковых молекул катализируется протеинкиназами:
протеин + АТФ фосфопротеин + АДФ
В подподкласс 2.7.11. входят большое количество протеинкиназ, включая рецепторную тирозиновую протеинкиназу (КФ 2.7.11.1), цАМФ-зависимую протеинкиназу (КФ 2.7.11.11), протеинкиназу С (КФ 2.7.11.13), кальций кальмодулин-зависимую протеинкиназу (КФ 2.7.11.17), митогенактивируемую протеинкиназу (МАПК) (КФ 2.7.11.24), киназу митогенактивируемой протеинкиназы (МАПКК) (КФ 2.7.11.25) и др.
17
КФ 3. ГИДРОЛАЗЫ
Ферменты, относящиеся к классу гидролаз, катализируют реакции гидролиза. При этом происходит разрыв связи в субстрате с присоединением воды:
A-B + H2O OH + B-H
Большинство гидролаз не содержат коферментов. Лишь немногие из них
осуществляют катализ в присутствии ионов металлов – главным образом Zn2+, Co2+, Ca2+, Mg2+.
Гидролазы локализованы во всех органеллах клетки, однако наибольшее их количество в лизосомах, где под их действием осуществляется гидролиз высомолекулярных соединений (белков, липидов, углеводов). Пищеварительные ферменты также относят к классу гидролаз.
В зависимости от типа связи, которую разрывают гидролазы делят на 13 подклассов. Наиболее важные из них: эстеразы, разрывающие сложноэфирные связи, гликозидазы и пептидазы.
Номер |
Название подкласса |
КФ 3.1 |
Действующие на эфирные связи. |
КФ 3.2 |
Гликозидазы |
КФ 3.3 |
Действующие на другие эфирные связи. |
КФ 3.4 |
Пептидазы |
КФ 3.5 |
Действующие на углерод-азот связи, отличные от |
|
пептидных |
КФ 3.6 |
Действующие на кислотные ангидриды |
КФ 3.7 |
Действующие на углерод-углерод связи |
КФ 3.8 |
Действующие на связи, образованные галогенами |
КФ 3.9 |
Действующие на связи, образованные фосфором и |
|
азотом |
КФ 3.10 |
Действующие на связи, образованные серой и азотом |
КФ 3.11 |
Действующие на связи, образованные углеродом и |
|
фосфором |
КФ 3.12 |
Действующие на дисульфидные связи |
КФ 3.13 |
Действующие на связи, образованные углеродом и |
|
серой |
КФ 3.1. Действующие на эфирные связи.
Эстеразы катализируют разрыв различных сложноэфирных связей. Среди них можно выделить гидролазы карбоновых кислот, гидролазы фосфомоноэфиров, эндонуклеазы, продуцирующие монофосфаты и многие другие. Деление на подклассы в зависимости от строения эфирной связи. Всего выделен 31 подподкласс, среди них:
КФ 3.1.1. гидролазы, действующие на карбоновые эфиры КФ 3.1.2. гидролазы тиоэфиров КФ 3.1.3. гидролазы фосфомоноэфиров
КФ 3.1.4. гидролазы фосфодиэфиров КФ 3.1.5. гидролазы фосфотриэфиров
18

КФ 3.1.6. гидролазы, действующие на эфирные связи, образованные серной кислотой
КФ 3.1.11. экзодеокирибонуклеазы, продуцирующие 5'-фосфомоноэфиры
В качестве примера карбоксиэстеразы (гидролазы, действующие на карбоновые эфиры), относящейся к 3.1.1 подподклассу, можно привести пищеварительный панкреатический фермент триглицерол липазу (КФ 3.1.1.3), катализирующий распад эфирных связей в триглицеридах:
Фермент предпочтительно гидролизует внешние эфирные связи и действует только на поверхности раздела фаз вода-липиды.
Кэтому же подклассу относят ферменты: 11-цис-ретинилпальмитат гидролазу (КФ 3.1.1.21), катализирующую гидролиз эфиров витамина А в кишечнике до 11-цис-ретинола и остатка пальмитиновой кислоты (фермент
активируется желчными кислотами), холинэстеразу (КФ 3.1.1.8), фосфолипазу А2 (КФ 3.1.14), липопротеинлипазу (КФ 3.1.1.34), гормонзависимую липазу жировой ткани (КФ 3.1.1.79), широкоспецифичную микросомальную эстеразу (КФ 3.1.1.1), катализирующую реакции гидролиза различных эфиров, образованных карбоновыми кислотами и спиртами.
К3.1.2 подподклассу гидролаз (гидролаз тиоэфиров) относят фермент, участвующий в синтезе кетоновых тел ацетоацетил-КoA гидролазу (КФ 3.1.2.11):
CH3 C |
|
C |
C SKoA + H2O |
|
CH3 C |
|
C |
|
C |
OH |
+ |
HSKoA |
||||
|
|
|
|
|||||||||||||
|
|
H2 |
|
|
|
|
|
|
H2 |
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
O |
|
|
|
O |
|
|
|
|||||||
O |
|
O |
|
|
|
|||||||||||
ацетоацетил-КоА |
|
ацетоацетат |
|
|
|
|||||||||||
Щелочная фосфатаза (КФ 3.1.3.1) и кислая фосфатаза (КФ 3.1.3.2) являются представителями гидролаз фосфомоноэфиров (3.1.3 подподкласса), расщепляют эфирные связи, образованные фосфорной кислотой и спиртом:
R-O-PO3 2-+ Н2О ROH + HPO3 2-
К гидролазам монофосфоэфиров (3.1.3 подподкласс) также относят многочисленную группу широкоспецифичных протеинфосфатаз (КФ 3.1.3.16), дефосфорилирующих белковые молекулы:
фосфопротеин + Н2О протеин + HPO3 2- Дефосфорилирование глюкозы катализирует глюкозо-6-фосфатаза (КФ
3.1.3.9):
глюкозо-6-Ф + Н2О глюкоза + HPO3 2-
Глюкозо-6-фосфатаза – фермент печени, необходимый для высвобождения глюкозы при распаде гликогена или глюконеогенезе.
19

КФ 3.2. Гликозидазы
Гликозидазы – ферменты гидролизующие гликозидные связи. Эти ферменты играют очень важную роль в обмене углеводов. Примером гликозидазы служит пищеварительный фермент -амилаза (КФ 3.2.1.1), катализирующая гидролиз внутренних 1,4- -D-глюкозидных связей в молекуле крахмала и других полимерных углеводов.
(глюкоза)n + mH2O (глюкоза )n-m + m(глюкоза)
Другими примерами гликозидаз, работающих в пищеварительном тракте являются сахаразо-изомальтазный комплекс, гликоамилазный комплекс, трегалаза.
сахаразо-изомальтазный комплекс Сахароза + H2O ---------------- глюкоза + фруктоза
Лизоцим (КФ 3.2.1.17) – фермент антибактериальной защиты катализирует гидролиз 1,4-β-связей между N-ацетилмураминовой кислотой N-ацетил-D- глюкозамином в пептидогликанах клеточной стенки бактерий.
КФ 3.3. Действующие на другие эфирные связи.
Примером гидролазы, относящейся к этому подподклассу является микросомальная эпоксидгидролаза (КФ 3.3.2.9), катализирующая реакцию:
+H2O
|
H |
|
H |
|
|
|
|
O |
H |
OH |
OH |
|
|||
|
|
||
H |
|
|
|
|
|
|
|
эп о ксид бен зан трацен а |
бен зан трацен дио л |
|
|
Это важный фермент печени, который вовлечен в метаболизм многих ксенобиотиков, таких как 1,3-бутадиен oксид, бензо-[a]пирен 4,5-оксид и др. Микросомальная эпоксидгидролаза участвует в обезвреживании эпоксидов, прежде всего эпоксидов полиароматических углеводородов, образующихся при участии цитохрома Р450.
КФ 3.4. Действующие на пептидные связи (пептидазы)
Гидролазы, расщепляющие пептидные связи – пептидазы и протеиназы представляют собой обширный подкласс гидролаз. Их субстратами являются белки и полипептиды. Подкласс включает 26 подподклассов в зависимости от каталитической специфичности ферментов, некоторые из которых:
20