

4. Поступление веществ в растительную клетку. Пассивное и активное поступления.
Из внешней среды в клетку растения непрерывно поступают питательные вещества. Это естественный процесс, без которого жизнедеятельность клетки была бы невозможна. Причем клетка осуществляет поглощение, несмотря на ограниченную проницаемость плазмалеммы. Живая клетка обладает способностью к избирательному накоплению питательных веществ. Анализируя данные таблицы можно сделать вывод, что ионный состав одноклеточной водоросли и окружающей среды резко различается. Клетка накапливает калий в концентрации, превышающей его содержание в морской воде в десятки раз. Вместе с тем морская вода содержит значительно большее количество натрия по сравнению с клеткой. Таким образом, клетка, несмотря на наличие полупроницаемой мембраны, обладает способностью к избирательному накоплению растворенных веществ.
На протяжении истории физиологии растений учеными были созданы многочисленные теории относительно механизмов проникновения растворенных веществ в клетку. Многие из этих теорий оказались несостоятельными, в частности из-за того, что под проникновением веществ в клетку понималось только их поступление в клеточный сок.
В настоящее время не вызывает сомнений, что поступление солей происходит в виде ионов и это проходит в несколько этапов. Питательные вещества могут поступать и накапливаться в клеточной оболочке, цитоплазме, вакуоли. Особенно важным этапом является поступление веществ в цитоплазму. Экспериментально показано, что питательные вещества могут поступать в цитоплазму и не проникать в вакуоль. Это хорошо видно из опытов Н.Г. Холодного по влиянию солей на форму плазмолиза. В растворах разных солей форма плазмолиза различна. Кальций повышает вязкость цитоплазмы, и, как следствие, форма плазмолиза в растворах его солей вогнутая.
Калий снижает вязкость форма плазмолиза выпуклая. Изменение вязкости цитоплазмы свидетельствует о том, что ионы проникли в нее. Вместе с тем наличие плазмолиза показывает, что соли если и проникли в вакуоль, то в малой степени.
Поглощение питательных веществ клеткой может быть пассивным и активным. Пассивное поглощение - это поглощение, не требующее затраты энергии. Оно связано с процессом диффузии и идет по градиенту концентрации данного вещества. Как уже рассматривалось выше, с термодинамической точки зрения направление диффузии определяется химическим потенциалом вещества. Чем выше концентрация вещества, тем выше его химический потенциал. Передвижение идет в сторону меньшего химического потенциала. Необходимо отметить, что направление движения ионов определяется не только химическим, но также электрическим потенциалом. Следовательно, пассивное передвижение ионов может идти по градиенту химического и электрического потенциала. Таким образом, движущей силой пассивного транспорта ионов через мембраны является электрохимический потенциал.
Электрический потенциал на мембране трансмембранный потенциал может возникать в силу разных причин:
Поступление ионов идет по градиенту концентрации (градиенту химического потенциала), однако благодаря разной проницаемости мембраны с большей скоростью поступает либо катион, либо анион. В силу этого на мембране возникает разность электрических потенциалов, что, в свою очередь, приводит к диффузии противоположно заряженного иона.
При наличии на внутренней стороне мембраны белков, фиксирующих определенные ионы. За счет фиксированных зарядов создается дополнительная возможность поступления ионов противоположного заряда (доннановское равновесие).
В результате активного (связанного с затратой энергии) транспорта либо катиона, либо аниона. В этом случае противоположно заряженный ион может передвигаться пассивно по градиенту электрического потенциала.
Активный транспорт - это транспорт, идущий против градиента электрохимического потенциала, т. е. по направлению от меньшего к большему его значению. Активный транспорт не может происходить самопроизвольно и требует затраты энергии, выделяющейся в процессе метаболизма. Активный перенос имеет решающее значение, поскольку обеспечивает избирательное концентрирование необходимых для жизнедеятельности клетки веществ.
Имеется ряд доказательств существования активного транспорта ионов. В частности, это опыты по влиянию внешних условий. Так, оказалось, что поступление ионов зависит от температуры. В определенных пределах с повышением температуры скорость поглощения веществ клеткой возрастает. В отсутствие, кислорода, в атмосфере азота, поступление ионов резко тормозится и может даже наблюдаться выход солей из клеток корня наружу. Под влиянием дыхательных ядов, таких, как KCN, СО, поступление ионов также затормаживается. С другой стороны, увеличение содержания АТФ усиливает процесс поглощения. Все это указывает на то, что между поглощением солей и дыханием существует тесная связь. Как известно дыхание является основным поставщиком энергии в клетке. Многие исследователи приходят к выводу о тесной взаимосвязи между поглощением солей и синтезом белка. Так, хлорамфеникол специфический ингибитор синтеза белка - подавляет и поглощение солей.
Способность клетки к избирательному накоплению питательных солей, зависимость поступления от интенсивности обмена служат доказательством того, что наряду с пассивным имеет место и активное поступление ионов. Оба процесса часто идут одновременно и бывают настолько тесно связаны, что их трудно разграничить.
Поступление веществ в клеточную стенку (1-й этап). Поглощение веществ клеткой начинается с их взаимодействия с клеточной оболочкой. Еще работами Д. А. Сабинина и И. И. Колосова было показано, что клеточная оболочка способна к быстрой адсорбции ионов. Причем эта адсорбция в ряде случаев носит обменный характер. В дальнейшем в опытах с выделенными клеточными оболочками было показано, что их можно рассматривать как ионообменник. На поверхности клеточной оболочки оказываются адсорбированными ионы Н+ и НСО3-, которые в эквивалентных количествах меняются на ионы, находящиеся во внешней среде. Ионы могут частично локаливоваться в межмицеллярных и межмолекулярных промежутках клеточной стенки, частично связываться и фиксироваться в клеточной стенке электрическими зарядами. Первый этап поступления характеризуется большой скоростью и обратимостью. Поступившие ионы легко вымываются. Это пассивный диффузионный процесс, идущий по градиенту электрохимического потенциала. Объем клетки, доступный для свободной диффузии ионов, включает клеточные стенки и межклеточные промежутки, т. е. апопласт или свободное пространство. По расчетам свободное пространство (СП) может занимать в растительных тканях 5-10% объема. Поскольку в клеточную оболочку входят амфотерные соединения (белки), заряд которых меняется при разных значениях рН, то в зависимости от значения рН скорость адсорбции катионов и анионов также может меняться.
Поступление веществ через мембрану (2-й этап). Для того чтобы проникнуть в цитоплазму и включиться в метаболизм клетки, вещества должны пройти через мембрану - плазмалемму. Перенос веществ через мембрану может идти пассивным и активным путем. При пассивном поступлении веществ через мембрану основой переноса и в этом случае является диффузия. Скорость диффузии зависит от толщины мембраны и от растворимости вещества в липидной фазе мембраны. Поэтому неполярные вещества, которые растворяются в липидах (органические и жирные кислоты, эфиры), легче проходят через мембрану.
 Однако
большинство веществ, которые важны для
питания клетки и ее метаболизма не могут
диффундировать через липидный слой и
транспортируются с помощью белков,
которые облегчают проникновение воды,
ионов, сахаров,
Однако
большинство веществ, которые важны для
питания клетки и ее метаболизма не могут
диффундировать через липидный слой и
транспортируются с помощью белков,
которые облегчают проникновение воды,
ионов, сахаров,
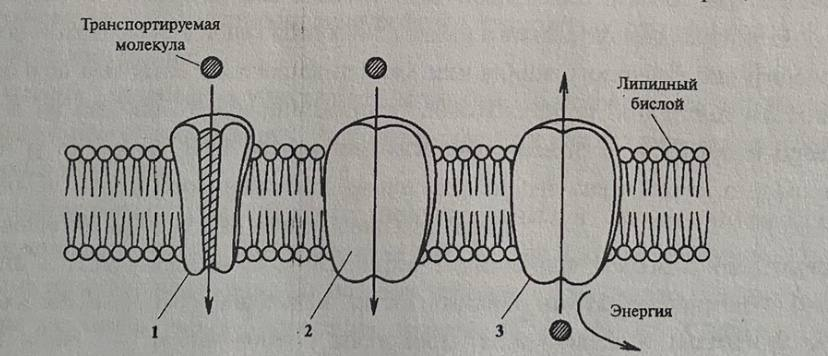
аминокислот и других полярных молекул в клетку. В настоящее время показано существование трех типов таких транспортных белков: каналы, переносчики, помпы (рис. 4).
Каналы - это трансмембранные белки, которые действуют как поры. Иногда их называют селективными фильтрами. Транспорт через каналы, как правило, пассивный Специфичность транспортируемого вещества определяется свойствами поверхности поры.
Как правило, через каналы передвигаются ионы. Скорость транспорта зависит от их величины и заряда. Если пора открыта, то вещества проходят быстро. Однако каналы открыты не всегда. Имеется механизм «ворот», который под влиянием внешнего сигнала открывает или закрывает канал.
Долгое время представлялась труднообъяснимой высокая проницаемость мембраны (10 мкм/с) для воды - вещества полярного и нерастворимого в липидах. В настоящее Время открыты интегральные мембранные белки, представляющие канал через мембрану для проникновения воды - аквапорины. Способность аквапоринов к транспорту воды регулируется процессом фосфорилирования. Было показано, что присоединение и отдача фосфатных групп к определенным аминокислотам аквапоринов ускоряет или тормозит проникновение воды, но не влияет на направление транспорта.
Переносчики - это специфические белки, способные связываться с переносимым веществом. В структуре этих белков имеются группировки, определенным образом ориентированные на наружную или внутреннюю поверхность. В результате изменения конформации белков вещество передается наружу или внутрь. Поскольку для транспорта каждой отдельной молекулы или иона переносчик должен изменить конфигурацию, скорость транспорта вещества в несколько раз меньше, чем происходит перенос через каналы. Показано наличие транспортных белков не только в плазмалемме, но и в тонопласте. Транспорт с помощью переносчиков может быть активным и пассивным, В последнем случае такой транспорт идет по направлению электрохимического потенциала и не требует затрат энергии. Этот тип переноса называется облегченной диффузией. Благодаря переносчикам он идет с большей скоростью, чем обычная диффузия.
Согласно представлениям о работе переносчиков, ион (М) реагирует со своим переносчиком (X) на поверхности мембраны или вблизи нее. Эта первая реакция может включать или обменную адсорбцию, или какое-то химическое взаимодействие. Ни сам переносчик, ни его комплекс с ионом не могут перейти во внешнюю среду. Однако комплекс переносчика с ионом (МХ) подвижен в самой мембране и передвигается к ее противоположной стороне. Здесь этот комплекс распадается и высвобождает ион во внутреннюю среду с образованием предшественника переносчика (Х'). Этот предшественник переносчика снова передвигается к внешней стороне мембраны, где вновь превращается из предшественника в переносчик, который на поверхности мембраны может соединиться с другим ионом. При введении в среду вещества, способного образовать прочный комплекс с переносчиком, перенос вещества блокируется. Опыты, проведенные на искусственных липидных мембранах, показали, что перенос ионов может проходить под влиянием некоторых антибиотиков, вырабатываемых бактериями и грибами, - ионофоров.
Транспорт с участием переносчиков обладает свойством насыщения, т. е. при увеличении концентрации веществ в окружающем растворе скорость поступления сначала возрастает, а затем остается: постоянной. Это объясняется ограниченным количеством переносчиков. Переносчики специфичны, т. е. участвуют в переносе только определенных веществ и, тем самым, обеспечивают избирательность поступления. Это не исключает того, что один и тот же переносчик может обеспечивать перенос нескольких ионов. Например, переносчик К+, обладающий специфичностью для этого иона, также переносит Rb+ и Na+, но не транспортирует Cl- или незаряженные молекулы сахарозы.
Транспортный белок, специфичный для нейтральных кислот, хорошо переносит аминокислоты глицин, валин, но не аспарагин или лизин. Благодаря разнообразию и специфичности белков осуществляется избирательная их реакция с веществами, находящимися в среде, и, как следствие, их избирательный перенос.
Насосы (помпы) - интегральные транспортные белки, осуществляющие активное поступление ионов. Термин «насос» показывает, что поступление идет с потреблением свободной энергии и против электрохимического градиента. Энергия, используемая для активного поступления ионов, поставляется процессами дыхания и фотосинтеза и в основном аккумулирована в АТФ.
Как известно, для использования энергии, заключенной в АТФ, это соединение должно быть гидролизовано по уравнению АТФ + НОН -> АДФ + Фи. Ферменты, осуществляющие гидролиз АТФ, называются аденозинтрифосфатазы (АТ Фазы). В мембранах клеток обнаружены различные АТФазы: К+ - Na+ - АТФаза; Ca+2 - АТФаза; Н+ - АТФаза.
H+- АТФаза (H+ - насос или водородная помпа) является основным механизмом aктивного транспорта в клетках растений, грибов и бактерий. Н+ - АТФаза функционирует в плазмалемме и обеспечивает выброс протонов из клетки, что приводит к образованию электрохимической разности потенциалов на мембране. Н* АТФаза переносит протоны в полость вакуоли и в цистерны аппарата Гольджи.
Расчет показывает, что для того, чтобы 1 моль соли диффундировал против градиента концентрации, необходимо затратить около 4600 Дж. Вместе с тем при гидролизе АТФ выделяется 30660 Дж/моль. Следовательно, этой энергии АТФ должно хватить для транспорта нескольких моль соли. Имеются данные, показывающие прямо пропорциональную зависимость, существующую между активность АТ Фазы и поступлением ионов. Необходимость молекул АТФ для осуществления переноса подтверждается еще и тем, что ингибиторы, нарушающие аккумуляцию энергии дыхания в АТФ (нарушение сопряжения окисления и фосфорилирования), в частности динитрофенол, тормозят поступление ионов.
Насосы делят на две группы:
1. Электрогенные, которые осуществляют активный транспорт иона какого-либо oдного заряда только в одном направлении. Этот процесс ведет к накоплению заряда одного типа на одной стороне мембраны.
2. Электронейтральные, при которых перенос иона в одном направлении сопровождается перемещением иона такого же знака в противоположном либо перенос двух ионов с одинаковыми по величине, но разными по знаку зарядами в одинаковом направлении.
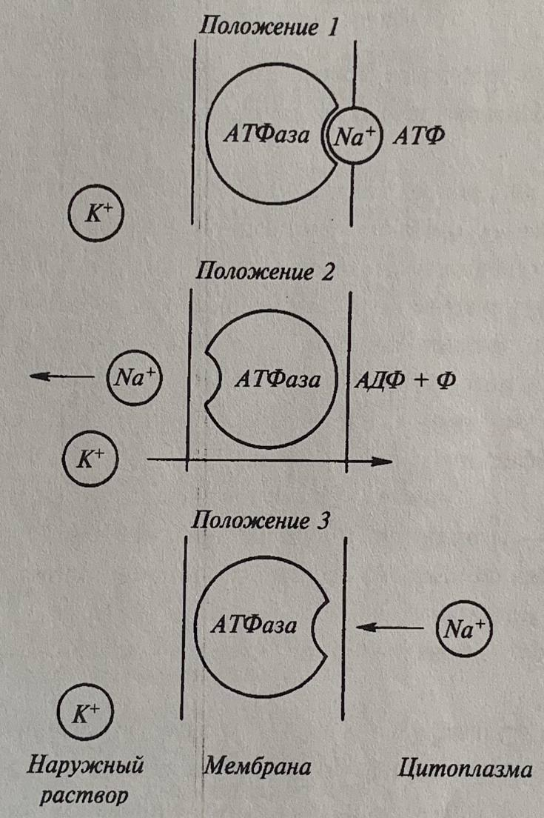
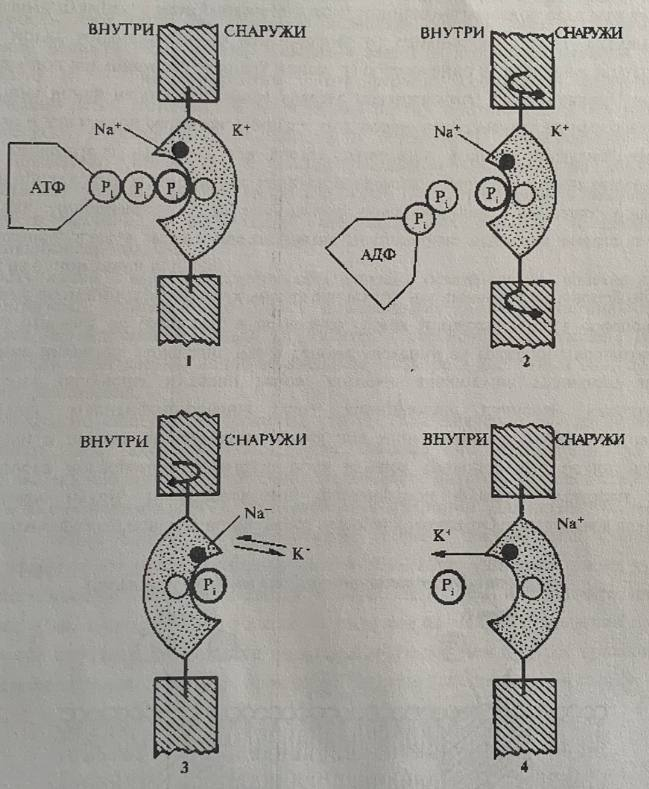
Рассмотрим механизм работы насоса на примере K+ - Na+ - АТФазы. Это электрогенный насос, поскольку переносит из клетки три иона натрия, а в клетку два иона калия. Это приводит к возникновению положительного заряда на мембране. Предполагается, Что АТФаза располагается поперек мембраны и происходит образование комплекса фермента с АТФ на внутренней поверхности мембраны, а также связывание ионов натрия. Освобождаемая при распаде АТФ энергия используется для изменения конфигурации самой АТФазы, благодаря чему участок фермента, связывающий Na+, поворачивается и оказывается по другую сторону мембраны (рис. 5, 6). На внешней поверхности мембраны происходит реакция ионного обмена Na+ на К+ и обратный поворот ферментного комплекса. Возвращение фермента в исходное состояние сопровождается освобождением ионов калия и неорганического фосфата. В этом случае АТФаза осуществляет гидролиз АТФ и использует выделяющуюся энергию для переноса ионов, непосредственно являясь переносчиком. Поэтому такой тип транспорта получил название первично-активный. Первично-активный транспорт связан с гидролизом АТФ или окислительно-восстановительными реакциями в электротранспортной цепи хлоропластов и митохондрий. Примером последнего служит непосредственное использование энергии дыхания на перенос ионов против градиента концентрации без предварительного накопления в АТФ. Механизм этого явления заключается в том, что в результате процесса дыхания на одной стороне мембраны (наружной) накапливаются ионы водорода, при этом внутренняя сторона мембраны заряжается отрицательно. Катионы поступают внутрь, притягиваясь к отрицательно заряженной внутренней стороне мембраны.
 Однако
существует и другой механизм активного
транспорта веществ, который
Однако
существует и другой механизм активного
транспорта веществ, который
называют вторично-активным. При этом переносчиками являются специальные белки, а энергия АТФ, высвобождаемая с помощью АТФазы, затрачивается на их перемещение в мембране. Благодаря работе Н+- АТФазы происходит выход протонов из клетки и на мембране возникает разность электрохимических потенциалов (Др. Н+). Это и используется для транспорта других ионов (веществ) при участии переносчиков. Так как первично-активный транспорт Н+ против градиента электрохимического потенциала опосредует транспорт другого иона (или молекулы) по градиенту электрохимического потенциала, поэтому такой тип транспорта называют вторично-активным.
Схемы вторично-активного транспорта
приведены на рисунке 7.
Если переносчик транспортирует два вещества в противоположном направлении, то такой встречный перенос получил название антипорт. Примером является поступление Na+, Mg+2, Mn+2 в антипорте с протонами при работе Н+- АТФазы. Вещества могут поступать и в одном направлении с выкачиваемым ионом. При этом переносчик осуществляет совместный однонаправленный перенос веществ или ионов, т. е. симпорт.
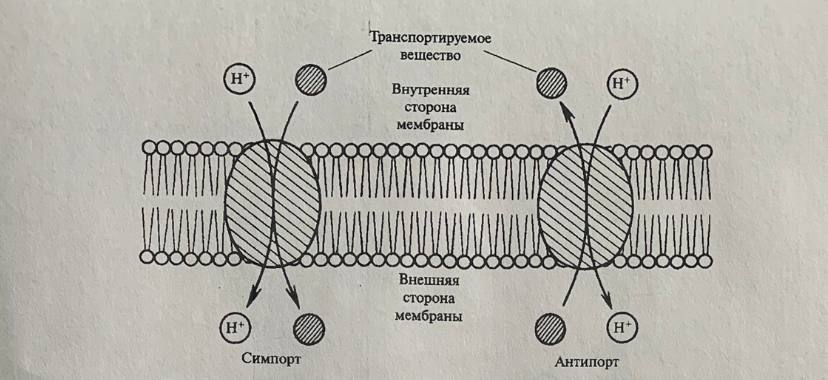 Экспериментальные
данные позволяют заключить, что в
симпорте с протонами в растительную
клетку поступают аминокислоты, сахара,
Сl-,
NO3-
и другие анионы. Перенос ионов калия
при низких концентрациях происходит
благодаря симпорту с протонами, а при
высоких идет через специальные калиевые
каналы.
Экспериментальные
данные позволяют заключить, что в
симпорте с протонами в растительную
клетку поступают аминокислоты, сахара,
Сl-,
NO3-
и другие анионы. Перенос ионов калия
при низких концентрациях происходит
благодаря симпорту с протонами, а при
высоких идет через специальные калиевые
каналы.
Таким образом, перенос ионов через мембрану может осуществляться активным пассивным путем. В обеспечении транспортной функции мембран и из-бирательности поглощения большую роль играют транспортные белки: каналы, переносчики и помпы. В настоящее время для многих транспортных белков гены клонированы.
Идентифицированы гены, кодирующие калиевые каналы.
Ha арабидопсисе получены мутации генов, которые влияют на транспорт и восстановление нитратов. Показано, что в геноме растений за транспорт веществ через мембраны отвечает не один ген, а несколько. Такая множественность обеспечивает выполнение функции в различных частях растений, что позволяет транспортировать вешества из одной ткани в другую.
Наконец, клетка может «заглатывать» питательные вещества вместе с водой (пиноцитоз). Пиноцитоз - это впячивание поверхностной мембраны, благодаря которому происходит заглатывание капелек жидкости с растворенными веществами. Явление пиноцитоза известно для клеток животных. Сейчас доказано, что оно характерно и для клеток растений. Процесс этот можно подразделить на несколько фаз: 1) адсорбция ионов на определенном участке плазмалеммы; 2) впячивание, которое происходит под влиянием заряженных ионов; 3) образование пузырьков с жидкостью, которые могут мигрировать по цитоплазме; 4) слияние мембраны, окружающей пиноцитозный пузырек, с мембранами лизосом, эндоплазматической сети или вакуоли и включение веществ в метаболизм. С помощью пиноцитоза в клетки могут попадать не только ионы, но и различные растворимые органические вещества.