Биологические особенности вирусов бактерий (бактериофагов). Особенности взаимодействия вирулентного и умеренного фагов с бактериальной клеткой. Морфология и ультраструктура фагов.
Ответ.
Бактериофаги (греч. phagos – пожирающий, лат. bacteriophaga – разрушающий бактерии) – это вирусы бактерий, обладающие способностью специфически и избирательно проникать в бактериальные клетки, репродуцироваться в них и при выходе потомства вызывать в большинстве случаев разрушение (лизис) бактерий.
Вирусы бактерий объединены в класс Bacteriophagae. Согласно Международной классификации и номенклатуре вирусов в зависимости от типа нуклеиновой кислоты бактериофаги подразделяются на ДНК- и РНК-содержащие. Большинство фагов относится к ДНК-содержащим вирусам с нуклеокапсидом, организованным по принципу смешанной симметрии.
Биологические особенности вирусов бактерий (бактериофагов).
простоорганизованные неклеточные существа;
избирательный внутриклеточный паразитизм (внутри БАКТЕРИИ);
не растут на питательных средах;
обладают определённой наследственностью, воспроизводя себе подобных;
геном – дву- или однонитевые ДНК или РНК;
обладают высокой специфичностью в отношении поражаемой клетки;
имеют антигенную обособленность от клетки-хозяина;
обладают выраженными антигенными свойствами.
Взаимодействие фага с бактериальной клеткой. По механизму взаимодействия различают вирулентные и умеренные фаги.
Вирулентные фаги, проникнув в бактериальную клетку, автономно репродуцируются в ней и вызывают лизис бактерий. Процесс взаимодействия вирулентного фага с бактерией протекает в виде нескольких стадий и весьма схож с процессом взаимодействия вирусов человека и животных с клеткой хозяина. Однако для фагов, имеющих хвостовой отросток с сокращающимся чехлом, он имеет особенности. Эти фаги адсорбируются на поверхности бактериальной клетки с помощью фибрилл хвостового отростка. В результате активации фагового фермента АТФазы происходит сокращение чехла хвостового отростка и внедрение стержня в клетку. В процессе «прокалывания» клеточной стенки бактерии принимает участие фермент лизоцим, находящийся на конце хвостового отростка. Вслед за этим ДНК фага, содержащаяся в головке, проходит через полость хвостового стержня и активно впрыскивается в цитоплазму клетки. Остальные структурные элементы фага (капсид и отросток) остаются вне клетки.
После биосинтеза фаговых компонентов и их самосборки в бактериальной клетке накапливается до 200 новых фаговых частиц. Под действием фагового лизоцима и внутриклеточного осмотического давления происходит разрушение клеточной стенки, выход фагового потомства в окружающую среду и лизис бактерии. Один литический цикл (от момента адсорбции фагов до их выхода из клетки) продолжается 30-40 мин. Процесс бактериофагии проходит несколько циклов, пока не будут лизированы все чувствительные к данному фагу бактерии.
Взаимодействие фагов с бактериальной клеткой характеризуется определенной степенью специфичности. По специфичности действия различают поливалентные фаги, способные взаимодействовать с родственными видами бактерий, моновалентные фаги, взаимодействующие с бактериями определенного вида, и типовые фаги, взаимодействующие с отдельными вариантами (типами) данного вида бактерий.
Умеренные фаги лизируют не все клетки в популяции, с частью из них они вступают в симбиоз, в результате чего ДНК фага встраивается в хромосому бактерии. В таком случае геномом фага называют профаг. Профаг, ставший частью хромосомы клетки, при ее размножении реплицируется синхронно с геном бактерии, не вызывая ее лизиса, и передается по наследству от клетки к клетке неограниченному числу потомков.
Биологическое явление симбиоза микробной клетки с умеренным фагом (профагом) называется лизогенией, а культура бактерий, содержащая профаг, получила название лизогенной. Это название отражает способность профага самопроизвольно или под действием ряда физических и химических факторов исключаться из хромосомы клетки и переходить в цитоплазму, т. е. вести себя как вирулентный фаг, лизирующий бактерии.
Лизогенные культуры по своим основным свойствам не отличаются от исходных, но они невосприимчивы к повторному заражению гомологичным или близкородственным фагом и, кроме того, приобретают дополнительные свойства, которые находятся под контролем генов профага. Изменение свойств микроорганизмов под влиянием профага получило название фаговой конверсии. Последняя имеет место у многих видов микроорганизмов и касается различных их свойств: культуральных, биохимических, токсигенных, антигенных, чувствительности к антибиотикам и др. Кроме того, переходя из интегрированного состояния в вирулентную форму, умеренный фаг может захватить часть хромосомы клетки и при лизисе последней переносит эту часть хромосомы в другую клетку. Если микробная клетка станет лизогенной, она приобретает новые свойства. Таким образом, умеренные фаги являются мощным фактором изменчивости микроорганизмов. Например, профаг придает дифтерийной палочке способность продуцировать экзотоксин.
Основные этапы взаимодействия фагов и бактерий.
1. Адсорбция (взаимодействие специфических рецепторов).
2. Внедрение вирусной ДНК (инъекция фага) осуществляется за счет лизирования веществами типа лизоцима участка клеточной стенки, сокращения чехла, вталкивания стержня хвоста через цитоплазматическую мембрану в клетку, впрыскивание ДНК в цитоплазму.
3. Репродукция фага.
4. Выход дочерних популяций.
Морфология.
Размер – 20-200 нм.
По форме бактериофаги подразделяются на следующие морфологические группы или типы:
- нитевидные фаги; (ЛипотриксИн)
- мелкие фаги без отростка; (ПлазмаКортикоМикрин)
- кубические фаги с аналогом (рудиментом) отростка; (ТектиЦистоЛевин)
- фаги с коротким отростком (хвостом); (ПодоФузеллы)
- фаги с длинным отростком и несокращающимся чехлом; (Сифовиры)
- фаги с длинным отростком и сокращающимся чехлом. (Миовиры)
Бактериофаги разных морфологических типов и семейств значительно отличаются друг от друга по своему строению. Бактериофаги первого морфологического типа представляют собой палочковидные или нитевидные структуры (Lipothrixviridae и Inoviridae).
Бактериофаги второго морфологического типа состоят из одной головки без отростка, относятся к семействам Plasmaviridae, Corticoviridae и Microviridae.
Бактериофаги третьего морфологического типа имеют головку и небольшие выступы или аналоги отростка (Tectiviridae, Cystoviridae и Leviviridae).
Бактериофаги четвертого морфологического типа содержат головку и короткий отросток (Podoviridae и Fuselloviridae).
Бактериофаги пятого морфологического типа состоят из головки и длинного отростка, чехол которого не способен сокращаться (Siphoviridae).
Шестой морфологический тип объединяет бактериофаги, состоящие из головки и отростка, окруженного сокращающимся чехлом (Myoviridae).
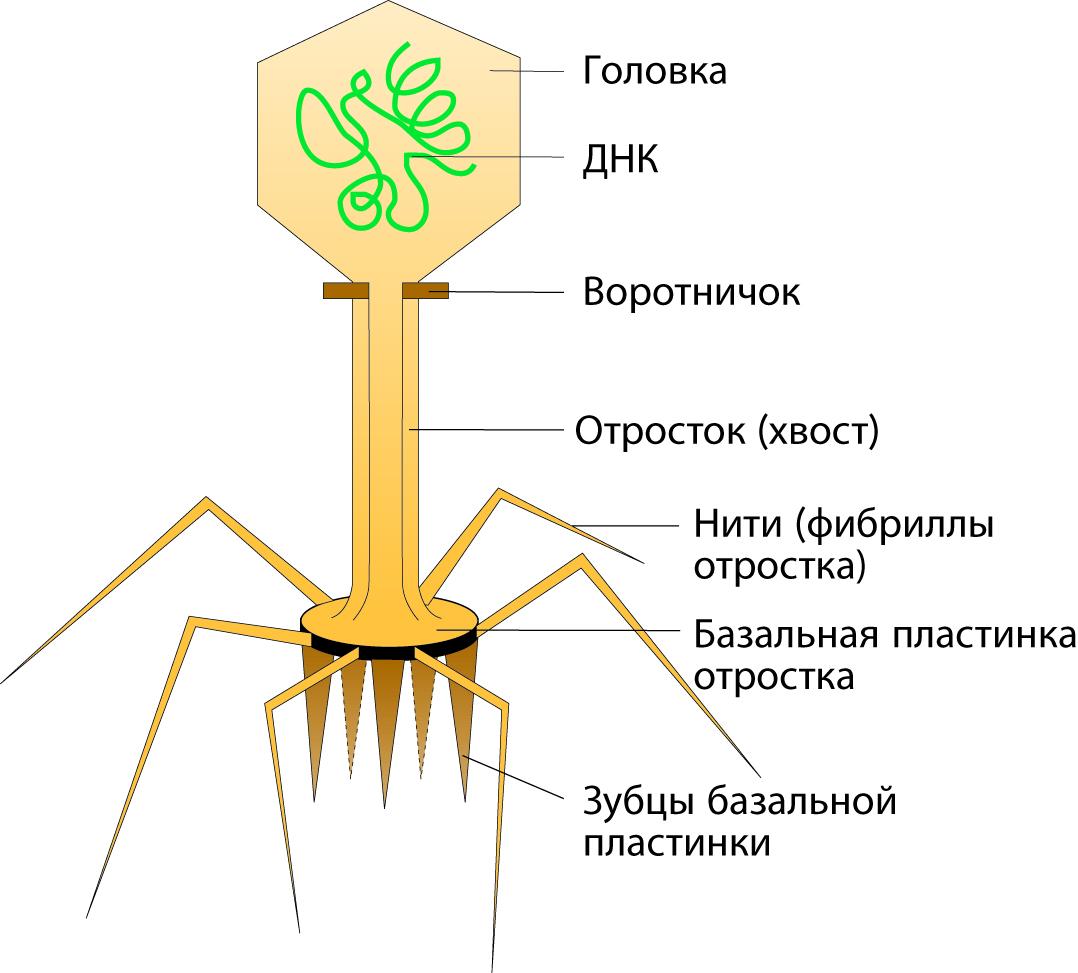
Типичная фаговая частица состоит из головки и хвостового отростка.
Длина хвостового отростка обычно в 2-4 раза больше диаметра головки.
Размеры бактериофагов колеблются от 2 до 200 нм. Чем крупнее бактериофаги, тем больше у них генов и сложнее их жизненный цикл.
Большинство бактериофагов напоминают сперматозоиды (головастики, барабанные палочки), то есть относятся к шестому морфологическому типу.
Ультраструктура.
Головка фага имеет округлую или овальную форму диаметром 60-95 нм. Внутри головки содержится геном бактериофага, представленный нуклеиновой кислотой. Нуклеиновые кислоты бактериофагов могут быть однонитевыми, двунитевыми, линейными, кольцевыми. У большинства фагов геном образует спирально упакованная двойная нить ДНК. Нуклеиновая кислота бактериофага окружена белковой оболочкой – капсидом. Капсид состоит из белковых молекул (капсомеров), организованных по принципу кубической симметрии. Нуклеиновая кислота и капсид вместе составляют нуклеокапсид.
Хвостовой отросток бактериофагов организован по принципу спиральной симметрии. Отросток имеет длину до 250 нм и толщину 10-25 нм. Он состоит из полого стержня и сократительного чехла, который присоединяется к воротничку, окружающему стержень около головки. Белковый стержень является продолжением белковой оболочки головки. Стержень заканчивается шестиугольной базальной пластинкой с шестью шипами (зубцами). От каждого зубца отходит по одной нити (фибриллы) длиной 150 нм. У Т-чётных фагов концы фибрилл опущены вниз, а у нечётных фагов концы нитей загнуты вверх. Базальная пластинка и нити обусловливают адсорбцию бактериофага на бактериальной клетке. У некоторых бактериофагов в дистальной части хвостового отростка содержится лизоцим (эндолизин), облегчающий проникновение нуклеиновой кислоты бактериофага в бактериальную клетку.
Фаговая частица содержит 40-50% нуклеиновой кислоты (РНК или ДНК), 50-60% белка, до 12-17% углеводов, 2% липидов.