

Таламус (зрительный бугор)
Таламус или зрительный бугор является частью промежуточного мозга, в который кроме него входят еще эпиталамус, subthalamus, и гипоталамус. Зрительный бугор представляет собой сложнейшее полифункциональное образование головного мозга. Таламус тесно связан морфологически и функционально с корой больших полушарий и подкорковыми структурами и поэтому принимает непосредственное участие во всех видах деятельности головного мозга, имеющих отношение, главным образом, к соматическим функциям.

Рис.4. Пространственное расположение и соотношение ядер таламуса.
Особенная роль в деятельности мозга принадлежит таламокортикальным взаимоотношениям. Многие реакции корковых нейронов на афферентные сигналы формируются в ядрах зрительного бугра. Тут же формируется и пространственно-временная структура импульсных ответов корковых нейронов (pattern) на сенсорные стимулы.
Крупнейший специалист в области физиологии зрительного бугра американский ученый A.E.Уолкер в 1966 году весьма образно охарактеризовал эту область мозга. Он писал, что «…в таламусе, этой огромной нейронной массе, расположенной ростральнее mesencephalon, лежит ключ к тайнам церебральной коры». Это, согласно A.E.Уолкеру, обусловлено тем, что таламус служит релейной станцией для поступления к коре головного мозга сенсорных сигналов и получает информацию от рецепторов всех органов чувств, кроме, обонятельных.
Предполагается, что таламус служит функции пропускания и фильтрации сенсорных сигналов, идущих в кору больших полушарий головного мозга.
Пути зрительного бугра. Таламус является важным этапом на пути проведения чувствительности. К таламусу подходят следующие чувствительные проводники (с противоположной стороны).
1. Медиальная петля с ее бульбо-таламическими волокнами (осязание, мышечно-суставное чувство, чувство вибрации и другие) и спиноталамическим путем (болевые и температурные ощущения).
2. Lemniscus trigemini – от чувствительного ядра тройничного нерва (чувствительность лица) и волокна от ядер языкоглоточного и блуждающего нервов (чувствительность глотки, гортани, а также внутренних органов).
3. Зрительные тракты, заканчивающиеся в pulvinar зрительного бугра и в corpus geniculatum laterale (зрительные пути).
4. Латеральная петля, заканчивающаяся в corpus geniculatum mediale (слуховые пути).
Таким образом, к зрительному бугру притекают импульсы экстероцептивной чувствительности, воспринимающей раздражения извне (боль, температуру, прикосновения, свет и прочее), проприоцептивной (мышечно-суставное чувство, чувство положения и движения) н интероцептивной чувствительности (от внутренних органов). Такое средоточие всех видов чувствительности в зрительном бугре станет понятным, если принять во внимание, что на определенных этапах эволюции нервной системы зрительный бугор был главным и конечным чувствительным центром, определяющим общие двигательные реакции организма рефлекторного порядка путем передачи раздражения на центробежные двигательные аппараты.
Функциональное деление ядер таламуса. В 1980 году Ф.Н.Серков и В.Н.Казаков в монографии «Нейрофизиология таламуса» дали обстоятельную структурную и функциональную классификацию ядер зрительного бугра. Связи с корой головного мозга и нейронное строение ядер таламуса позволили разделить их на шесть групп: релейные сенсорные ядра, релейные моторные ядра, релейные несенсорные, ассоциативные ядра, неспецифические и ядра с подкорковой связью.
К релейным сенсорным ядрам относятся: латеральное коленчатое тело (GL) – релейное ядро зрительной сенсорной системы, проецируется в поля 17,18,19 зрительной коры. Следующее ядро – медиальное коленчатое тело (GM) – релейное ядро слуховой сенсорной системы, проецируется в поля 41, 42, 22 теменной коры. Далее – вентральное заднелатеральное ядро (VPL) – релейное ядро соматической и висцеральной чувствительности туловища и конечностей и внутренних органов и вентральное заднемедиальное ядро (VPM) – релейное ядро соматической и вкусовой сенсорной систем, идущих от лица и языка, проецируется в поля 3,1,2 соматосенсорной коры (задняя центральная извилина. Наконец, релейное ядро проприоцептивной чувствительности (VIM – вентральный отдел VL), которое проецируется в поля 3 и 6 соматосенсорной и премоторной коры.
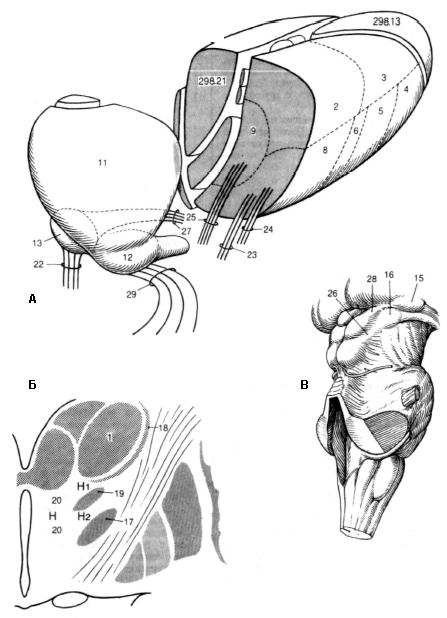
Рис.5. Детальное описание ядер и путей таламуса. 1 – вентролатеральные ядра (nucl. ventrolaterals). Лежат латерально от внутренней мозговой полоски; 2 – заднее латеральное ядро (nucl. lateralis posterior). Расположено между подушкой и дорсальным латеральным ядром. Связано с корой теменной доли; 3 – дорсальное латеральное ядро (nucl. lateralis dorsalis). Расположено спереди и сверху относительно остальных вентролатеральных ядер. Связано с корой нижней теменной дольки и задней части поясной извилины; 4 – переднее вентральное ядро (nucl. ventralis anterior). Самое переднее из вентролатеральных ядер. Соединяется с внутрипластинчатыми ядрами, бледным шаром и зубчатым ядром мозжечка, а также имеет двухсторонние связи с корой прецентральной извилины; 5 – вентральное промежуточное ядро (nucl.ventralis intermedius). Расположено позади переднего вентрального ядра. Входит в состав пути, связывающего ядра мозжечка и красное ядро с моторной корой; 6 – медиальное вентральное ядро (nucl. ventralis medialis) – нечетко очерченная группа нейронов с неясной функцией спереди от задних вентральных ядер; 7 – задние вентральные ядра. Представлены двумя группами; 8 – заднелатеральное вентральное ядро (nucl. ventralis posterolateralis). Латеральная часть заднего вентрального ядерного комплекса. На нейронах этого ядра заканчиваются волокна медиальной петли и спиноталамического пути и заканчиваются в постцентральной извилине; 9 – заднемедиальное вентральное ядро (nucl.ventralis posteromedialis). Находится между центральным срединным и заднелатеральным вентральным ядрами. К нему подходят волокна тройничной петли; 10 – задние ядра (nuclei posterioris), три ядерных комплекса; 11 – ядро подушки (nucl.pulvinar). Занимают заднюю часть таламуса. Входит в состав проводящих путей слуховой и зрительной сенсорных систем, а также связан с другими ядрами таламуса, зрительным и слуховым корковыми центрами речи; 12 – латеральное коленчатое тело – дорсальная часть (nucl. corporis geniculati lateralis, pars dorsalis). Нейроны этого ядра входят в состав зрительного пути; 13 – медиальное коленчатое тело – дорсальная часть (nucl.corporis geniculati media-lis, pars dorsalis). Состоит из мелких нейронов; 14 – разрезы переднего таламуса; 15 – ядро латерального коленчатого тела, вентральная часть (nucl. corporis geniculati lateralis, pars ventralis). К нейронам ядра подходят волокна от сетчатки. Входит в состав рефлекторной дуги зрачкового рефлекса; 16 – ядро медиального коленчатого тела, вентральная часть (nucl. corporis geniculati medialis, pars ventralis). Основная часть соответствующего коленчатого тела имеет отношение к слуховому пути; 17 – субталамическое ядро (nucl. subtha-lamicus – corpus Luysii). Находится между внутренней капсулой и неопределенной зоной; 18 – ретикулярные ядра (nucl. reticulares thalami). Расположены в латеральной части таламуса между наружной мозговой пластинкой и внутренней капсулой; 19 – неопредленная зона, zona incerta. Является продолжением ретикулярных ядер таламуса и связана с бледным шаром; 20 - ядра полей Н, HI и Н2 (nucl. regionum Н, HI, Н2). Группы диффузно расположенных нейронов в составе полей Фореля. Поле Н лежит медиально от zona incerta и спереди от красного ядра. Поле HI расположено между таламусом и неопределенной зоной. Поле Н2 находится между zona incerta и субталамическим ядром; 21 – пути и пучки таламуса (tractus et fasciculi thalamicus); 22 – латеральная петля (lemniscus lateralis). Образована волокнами слухового пути, которые вступают в медиальное коленчатое тело; 23 – медиальная петля (lemniscus medialis). Ее волокна заканчиваются на нейронах вентрального заднелатерального ядра; 24 – спинномозговая петля (lemniscus spinalis). Путь, проводящий болевую чувствительность. Заканчивается в вентральном заднелатеральном ядре; 25 – тройничная петля (lemniscus trigeminalis). Образована отростками нейронов ядер тройничного нерва, которые заканчиваются в вентральном заднемедиальном ядре; 26 – ручка нижнего холмика. Соединяет нижний холмик с медиальным коленчатым телом; 27 – слуховая лучистость (radiatio acustica). Начинается от медиального коленчатого тела, проходит через заднюю ножку внутренней капсулы и заканчивается в височных извилинах; 28 – ручка верхнего холмика (brachium collicu superior). Соединяет верхний холмик с латеральным коленчатым телом. Связывает зрительный путь с экстрапирамидной системой; 29 – зрительная лучистость (radiatio optica Gratiolet). Начинается от латерального коленчатого тела, проходит через заднюю ножку внутренней капсулы и заканчивается в зрительной коре (область шпорной борозды).
К релейным моторным ядрам относятся: вентролатеральное ядро (VL) – релейное соматическое и моторное ядро, проводящее мозжечковые и лентикулярные сигналы, которое проецируется в 4 и 6 поля моторной и премоторной коры. Следующее ядро – наружный отдел вентрального переднего ядра (VA) – релейное моторное ядро, проводящее паллидарные и лентикулярные сигналы, проецируется в теже поля, а также поле 13 орбитофронтальной коры. Вентральное медиальное ядро (VM) – релейное моторное ядро, проводящее церебеллярные и стриопаллидарные импульсы, проецируется в поля 13 и 14 орбитофронтальной коры.
К релейным несенсорным ядрам относится группа передних ядер – вентральное (AV), дорсальное (AD) и медиальное (AM) – это релейные ядра маммиллоталамического тракта, переключающие сигналы к цингулярной коре – в поля 24,23 (для AV), поле 29 (AD) и поля 24,32 (AM).
К ассоциативным ядрам относятся: подушечное ядро (Pulvinar) – ядро, проводящее зрительные и слуховые сигналы в ассоциативные поля теменной (поя 5, 7, 21) и зрительной (поля 18 и 19) коры, латеральные – заднее (LP) и дорсальное (LD) ядра, проводящие зрительные, слуховые, соматические и висцеральные сигналы в теменную ассоциативную область (поля 5, 7, 21), медиодорсальное ядро (MD) обслуживает проведение периферических и центральных сигналов в орбитофронтальную кору (поля 8 и 12), группа задних ядер (PO), которые проводят разномодальные сигналы к третичным зонам основных видов чувствительности.
К неспецифическим ядрам относятся: интраламинарная группа (CL, Pc, Sm), проецируется на весь неокортекс, а также парафасцикулярный комплекс (Pf, sPf, CM) – ядра проводящие разномодальные сигналы к обширным территориям коры головного мозга, крупноклеточный отдел вентрального переднего ядра (VAmc), осуществляющее передачу ретикулярные неспецифические влияния на обширные территории коры головного мозга, ретикулярное таламическое ядро (R), обеспечивающее контроль активности релейных систем таламуса с помощью возвратного торможения.
К ядрам с подкорковой связью относятся ядра средней линии таламуса (Pv, CM, Rb, Ru, GS). Эти ядра не имеют проекций в коре головного мозга и осуществляют внутриталамическую интеграцию неспецифических ядер таламуса.
Организация связей с корой головного мозга. С появлением и развитием коры головного мозга усложняется и совершенствуется чувствительная функция, появляется способность тонкого анализа, дифференцировки и локализации раздражении. Основная роль в чувствительной функции переходит к коре головного мозга.
Однако ход чувствительных путей остается прежним, возникает лишь продолжение их от зрительного бугра к коре головного мозга. Зрительный бугор становится в основном лишь передаточный станцией на пути импульсов от периферии к коре.
Действительно, существуют многочисленные таламо-кортикальные пути (tractus thalamo-corticalis), с чувствительными нейронами (в основном, третьими), о которых следует упомянуть: во-первых, это третьи нейроны кожной и глубокой чувствительности (болевого, температурного, осязательного, суставно-мышечного чувства), начинающиеся из вентро-латерального отдела зрительного бугра, проходящие через внутреннюю капсулу в область задней центральной извилины и теменную долю; во-вторых, зрительные пути от первичных зрительных центров (corpus geniculatum laterale и radiatio optica) или пучок Грациоле, в область fissurae calcarinae затылочной доли; в- третьих, слуховые пути от первичных слуховых центров (corpus geniculatum mediale).
Рис.6. Связи ядер таламуса с корой головного мозга. Однородными цветами показаны ядра и их проекции в коре головного мозга.
Помимо названных связей, зрительный бугор имеет пути, связывающие со стриопаллидарной системой.
Аналогично тому, как thalamus opticus является на определенных этапах развития нервной системы высшим чувствительным центром, стриопаллидарная система была конечным двигательным аппаратом, осуществляющим достаточно сложную рефлекторную деятельность. Поэтому связи зрительного бугра с названной системой весьма интимны, и весь аппарат в целом может быть назван таламостриопаллидарной системой с воспринимающим звеном в виде thalamus opticus и двигательным в виде стриопаллидарного аппарата.
О связях зрительного бугра с корой головного мозга – в направлении таламус-кора уже было сказано. Кроме того, существует мощная система проводников обратного направления, от коры головного мозга к зрительным буграм. Эти пути исходят из различных отделов коры головного мозга (tractus сorti-cothalamicus). Наиболее массивным из них является тот, который начинается из лобной доли.
Наконец, следует упомянуть о связях зрительного бугра с гипоталамусом, где сосредоточены подкорковые центры вегетативной иннервации.

Рис.7. Схема основных восходящих и нисходящих путей центральной нервной системы (восходящие пути обозначены синим цветом, нисходящие – красным, стрелки указывают направление проведения нервных импульсов, гигантские пирамиды двигательной области коры головного мозга обозначе-ны треугольниками красного цвета): 1 – латеральный кортикоспинальный (пирамидный) путь; 2 – передний кортикоспиналь-ный (пирамидный) путь; 3 – перекрест пирамид; 4 – тонкое ядро; 5 – двигате-льные ядра черепных не-рвов; 6 – передний и латеральный кортикоспиналь-ные (пирамидные) пути; 7 – спинномозговая петля; 8 – неперекрещенные волокна корково-ядерного пути; 9 – внутренняя капсула; 10 и 16 – гигантские пирамиды нижних отделов передней центральной извилины; 11 – чечевицеобразное ядро; 12 – таламокортикальные пучки; 13 – гигантские пирамиды верхних отделов передней центральной извилины; 14 – хвостатое ядро; 15 – III желудочек; 17 – вентролатеральные ядра таламуса; 18 – перекрещенные волокна корково-ядерного пути; 19 – медиальная петля и петля тройничного нерва; 20 – чувствительные узлы черепных нервов; 21 – чувствительные волокна в составе черепных нервов; 22 – чувствительные ядра черепных нервов; 23 – задние и наружные дугообразные волокна; 24 – клиновидное ядро; 25 – клиновидный пучок; 26 – спинномозговой узел; 27 – чувствительные волокна спинномозговых нервов; 28 – тонкий пучок; 29 – латеральный спиноталамический путь; 30 – нейроны заднего рога спинного мозга.
Связи ядерных образований таламической области весьма многочисленны, сложны и в деталях изучены еще недостаточно. В последнее время на основании, главным образом, электрофизиологических исследований, предлагается делить таламо-кортикальные системы на специфические (связанные с определенными проекционными зонами коры) и неспецифические, которые начинаются от медиальной группы ядер зрительного бугра (срединный центр, интраламинарные, ретикулярные и другие ядра).
Функции зрительного бугра. Таламус является своеобразным коллектором чувствительных путей, местом, в котором концентрируются все пути, проводящие чувствительные импульсы, идущие от противоположной половины тела. Кроме того, в переднее его ядро по сосцевидно-таламическому пучку поступают обонятельные импульсы.
Таламические ядра, получающие импульсы от строго определенных участков тела и передающие эти импульсы в соответствующие ограниченные зоны коры (первичные проекционные зоны), называются проекционными, специфическими или переключающими ядрами. К ним относятся вентролатеральные ядра. Переключающие ядра для зрительных и слуховых импульсов заложены соответственно в латеральных и медиальных коленчатых телах, прилежащих к задней поверхности зрительных бугров.
Наличие в проекционных ядрах таламуса, прежде всего, в вентролатеральных ядрах, определенного соматотопического представительства делает возможным при ограниченном по объему патологическом очаге в таламусе развитие расстройства чувствительности и сопряженных с этим двигательных нарушений в какой-либо ограниченной части противоположной половины тела.
Ассоциативные ядра, получая чувствительные импульсы от переключающих ядер, подвергают их частичному обобщению – синтезу. В результате из этих таламических ядер к коре большого мозга направляются импульсы, уже усложненные, вследствие синтеза поступающей сюда информации. Следовательно, таламус является не только промежуточным центром переключения, но может быть и местом частичной переработки чувствительной импульсации.
Кроме переключающих и ассоциативных ядер, в таламусе находятся, как уже упоминалось, интраламинарные (парафасцикулярное, срединное и медиальное, центральные, парацентральное ядра), ретикулярные ядра, как будто не имеющие специфической функции. Они рассматриваются как часть ретикулярной формации и объединяются под названием неспецифической диффузной таламической системы. Будучи связанной с корой больших полушарий и структурами лимбико-ретикулярного комплекса, эта система принимает участие в регуляции тонуса коры больших полушарий и играет определенную роль в сложном механизме формирования эмоций и соответствующих им выразительных непроизвольных движений, мимики, плача и смеха.
Таким образом, к таламусу по афферентным путям сходится информация практически от всех рецепторных зон. Эта информация подвергается существенной переработке. Отсюда к коре больших полушарий направляется лишь часть ее, другая же и, вероятно, большая часть принимает участие в формировании безусловных и, возможно, некоторых условных рефлексов, дуги которых замыкаются на уровне таламуса и образований стриопаллидарной системы. Следовательно, таламус являются важнейшим звеном афферентной части рефлекторных дуг, обусловливающих инстинктивные и автоматизированные двигательные акты, в частности привычные локомоторные движения (ходьба, бег, плавание, езда на велосипеде, катание на коньках и прочее).
Поражение ядер зрительного бугра. Сложность организации и многообразие функций таламуса определяет полиморфизм возможных клинических проявлений его поражения.
При поражении зрительного бугра могут возникать симптомы выпадения его функций или симптомы раздражения. В первом случае на противоположной стороне наблюдается гемианестезия, касающаяся всех видов чувствительности, как поверхностной, так и глубокой. Расстройства чувствительности более выражены в дистальных отделах конечностей, где выпадения мышечно-суставного чувства выражены, обычно, особенно резко. Поэтому в конечностях наблюдается одностороннее расстройство координации движений, так называемая, сенситивная гемиатаксия, связанная с нарушением глубокой чувствительности. Вследствие поражения подкорковых зрительных центров (corpus geniculatum laterale) возникает также и гемианопсия – утрата зрения в противоположных поражению полях зрения. Наконец, при поражении зрительного бугра может наблюдаться парез (неполный паралич) мимической мускулатуры, тоже на противоположной стороне, сказывающийся только при эмоциональных мимических движениях, например при улыбке или смехе. При движениях «по заданию» этих расстройств может не отмечаться.
Поражение вентролатеральной части таламуса обычно ведет к повышению порога чувствительности на стороне, противоположной патологическому очагу, при этом меняется аффективная окраска болевых и температурных ощущений.
При поражении заднелатеральной части таламуса может проявиться таламический синдром Дежерина-Русси, включающий в себя жгучие, жестокие, подчас невыносимые таламические боли в противоположной половине тела в сочетании с нарушением поверхностной и особенно глубокой чувствительности, с явлениями гиперпатии и дизестезии. Характер этих «центральных» болей с трудом описывается больными, чаще это – мучительное ощущение жжения, холода, нестерпимой боли. Локализуются они неясно и обычно диффузны. Интенсивность их усиливается в зависимости от внешних раздражении и, особенно, эмоций. Характерна в соответствующей части противоположной половины тела гипалгезия (снижение чувствительности) в сочетании с гиперпатией – в виде резкого чувства неприятного, с неточной локализацией при уколе и температурных раздражениях, иногда извращенное восприятие раздражения, неточность локализации его и иррадиация.
Нередко наблюдается повышенная аффективность (раздражительность), насильственный смех и плач. Возможно присоединение ряда вегетативных расстройств. Все эти симптомы легко объясняются той ролью и значением зрительного бугра, о чем было сказано выше. Этот синдром чаще возникает при развитии ишемии в латеральных артериях таламуса (а. thalamica lateralis) – ветвях задней мозговой артерии. Иногда при этом на стороне, противоположной патологическому очагу, возникает преходящий гемипарез (паралич) и развивается гомонимная гемианопсия (выпадение поля зрения на противоположной поражению стороне). Следствием расстройства глубокой чувствительности может быть сенситивная гемиатаксия и псевдоастериогноз (невозможность опознания знакомых предметов по ощупывании при закрытых глазах).
В случае поражения медиальной части таламуса, зубчато-таламического пути, по которому к таламусу проходят импульсы от мозжечка, и руброталамических связей на противоположной патологическому очагу стороне появляется атаксия в сочетании с атетоидным или хореоатетоидным гиперкинезом (непроизвольные движения, выражающиеся медленной тонической судорогой мышц конечностей, лица, туловища), особенно выраженным в кисти и пальцах («таламическая» рука). В таких случаях характерна тенденция к фиксации руки в определенной позе: плечо прижато к туловищу, предплечье и кисть согнуты и пронированы, основные фаланги пальцев согнуты, остальные разогнуты. Пальцы руки при этом совершают медленные вычурные движения атетоидного характера.
Таламический инсульт. Сосудистая катастрофа в бассейне средней мозговой артерии сопровождается инсультом с сенсорными и моторными расстройствами.
Сенсомоторный инсульт. При этом у больного наблюдаются сенсорные расстройства, которые сопровождают моторные нарушения на той же стороне в виде гемипареза, повышения сухожильных рефлексов и симптома Бабинского.

Рис.8. Характерное нарушение движения (парез) после перенесенного таламического инсульта.
Эти явления наиболее ярко проявляются при увеличении зоны инфаркта до задней части внутренней капсулы, смежной с вентролатеральными ядрами. При инфаркте вентролатеральных, медиальных или таламопариетальных отделов таламуса развивается грубое нарушение поверхностной и глубокой чувствительности.
Инфаркты в латеральных отделах таламуса (бассейн ветвей a. thalamogeniculata) проявляются двигательными расстройствами (неловкость и атаксия), входящими в структуру таламического синдрома Дежерина-Русси и имеют клиническую особенность, которая связана с нарушением моделей движения в результате повреждения экстрапирамидных волокон, идущих от базальных ганглиев через ansa lenticularis либо от верхних ножек мозжечка и красного ядра, которые образуют синапсы в вентролатеральных ядрах таламуса. Подобные нарушения наблюдаются также при поражении волокон от заднего бедра внутренней капсулы, которое примыкает к вентролатеральной части таламуса. Двигательные расстройства всегда сопровождаются ощущением боли с крайне неприятным чувственным оттенком, включающим, как правило, гиперестезию, гипералгезию и аллодинию (восприятие неболевого раздражения как болевого), а также дистонические феномены.
Даже при расстройствах мышечно-суставного чувства у пациентов могут наблюдаться характерные черты мозжечкового типа гемиатаксии, гиперметрии, осцилляций (подергиваний) и дисдиадохокинеза. У некоторых больных утрачивается способность стоять и ходить, что становится преобладающим симптомом и называется «таламическая астазия». Такие нарушения движений, как гемидистония и подергивания в руке, могут развиться через несколько недель после инсульта, особенно у пациентов с расстройством чувствительности и атаксией. Когнитивные способности и поведение сохраняются у пациентов с латеральными таламическими инфарктами.