

Промежуточный мозг
Организм находится в непрерывном взаимодействии с окружающей средой. Это взаимодействие очень многогранно, оно обусловлено, с одной стороны, степенью сложности организации животного, а с другой – изменениями, которые постоянно происходят во внешней среде и в самом организме. Так как внешняя среда служит для организма не только источником, откуда он черпает материал для своего существования, но и таит в себе различные для него опасности, то вполне понятно, что организм должен очень четко воспринимать различного рода раздражения и не менее четко на них реагировать. В связи с этим и развились высокодифференцированные органы нервной системы, приспособленные воспринимать и анализировать раздражения, идущие не только из внешней среды, но и из всех без исключения органов и тканей самого организма, и координировать его деятельность, проявляющуюся в поведении, а также работу всех его органов и происходящий в них обмен веществ.
Таким образом, нервная система, являясь общей управляющей инстанцией, обеспечивает целостность организма, как живой системы, взаимообусловленность всех его частей, единство организма и внешней среды.
Все эти функции чрезвычайно важны, хотя мы и не замечает этого в повседневной жизни.
Промежуточный мозг в этом представлении есть той структурой, которая уже может обеспечивать все означенные функции. Промежуточный мозг – diencephalon – занимает довольно значительный участок головного мозга с обширной полостью третьего желудочка. В его состав входят таламус или зрительный бугор, представленный множественными ядрами, эпиталамус, состоящий из непарной железы – рудимента третьего, теменного глаза – эпифиза, который прикреплен к таламусу ядрами уздечки, субталамической областью и ядрами гипоталамуса.
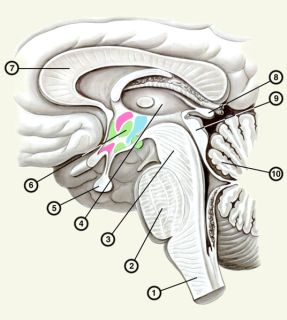
Рис.1. Ствол головного мозга (на сагиттальном разрезе): 1 – продолговатый мозг; 2 – мост; 3 – ножки мозга; 4 – таламус; 5 – гипофиз; 6 – проекция субталамической области и гипоталамических ядер; 7 – мозолистое тело; 8 – шиш-ковидное тело; 9 – бугры четверохолмия; 10 – мозжечок.
В состав промежуточного мозга входит гипофиз, придаток мозга – hypophysis – связанный с гипоталамусом посредством воронки. Гипофиз состоит из трех частей неодинакового происхождения, строения и различных по функции. У высших животных гипофиз у состоит из трех частей: дорзальной – нервной – neirohypohpysis, вентральной – железистой – adenohypohpysis и промежуточной. Железистая часть выделяет инкреты непосредственно в кровеносные сосуды (в кровь), а промежуточная и нервная – в просвет третьего мозгового желудочка. Отделы гипофиза выделяют множество гормонов, среди которых есть тропные, то есть такие которые управляют выделением гормонов другими железами внутренней секреции и конечные гормоны, имеющие место приложения в организме.
Филогенез и онтогенез промежуточного мозга. В филогенезе структуры промежуточного мозга появляются и развиваются на разных его этапах. У низших позвоночных животных уже есть некоторые эпиталамические, субталамические и гипоталамические структуры. У круглоротых и таламус, и гопоталамус уже достаточно выражены. Таламус разделяется на дорсальную и вентральную части, включающие в себя несколько ядер. Гипоталамус также имеет множество ядерных образований.
У костистых рыб происходит усложнение в организации таламуса и дифференциация ряда ядерных групп. У амфибий и рептилий развиваются зрительные бугры. Они становятся специальными структурами и связывают средний мозг с конечным мозгом. У рептилий промежуточный мозг выполняет, вместе с некоторыми структурами конечного мозга, высшие интегративные функции.
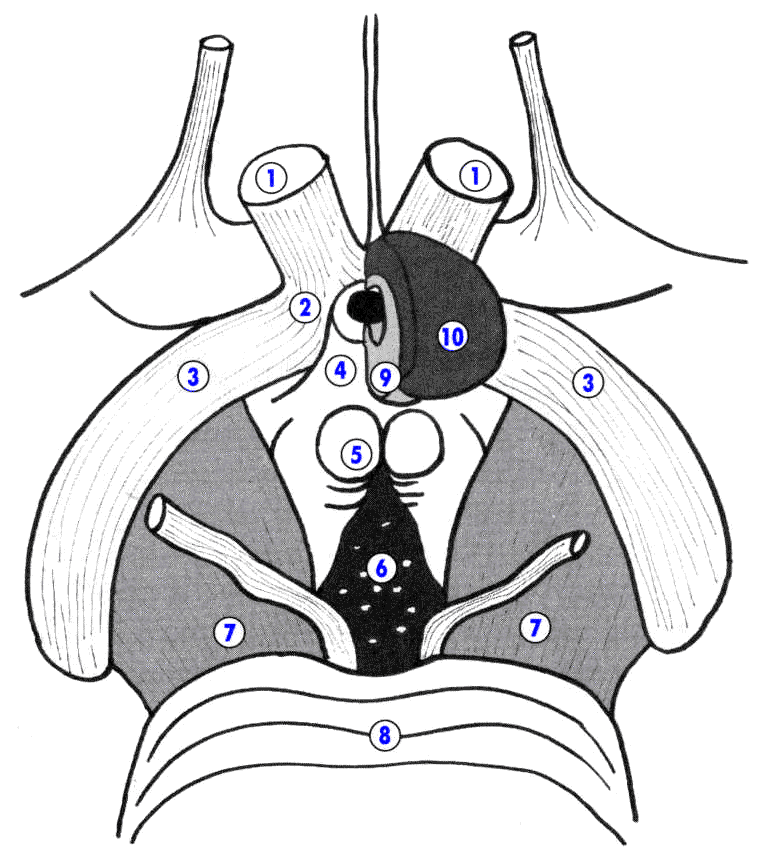
Рис.2. Промежуточный мозг c вентральной поверхности (схематично, увеличено). 1 – зрительный нерв – n. opticus (II); 2 – зрительный перекрест – chiasma opticum; 3 – зри-тельный тракт – tractus opticus; 4 – воронка – infundibulum; 5 – сосцевидное тело – corpus mammillarae; 6 – заднее продырявленное пространство – substantia perfora-ta posterior; 7 – ножка мозга – pedunculus cerebri; 8 – мост – pons; 9 – нейрогипофиз – neurohypophysis; 10 – аденогипофиз – adenohypophysis.
Дорсальный таламус у рептилий отделяется от вентрального, на этом этапе выделяются четкие ядра с характерным строением и связями. В дорсальном таламусе их насчитывается девять, в вентральном – семь. В переднем отделе гипоталамуса у рептилий появляются зачатки супраоптического и паравентрикулярного ядер. У птиц дифференциация ядер гипоталамуса хорошо выражена, тогда как таламус усложняется незначительно. Особенно бурно идет развитие дорсального таламуса у млекопитающих, так как он выполняет функцию основного коллектора сенсорных путей к коре больших полушарий. У млекопитающих наряду с развитием релейных (переключательных) ядер таламуса развиваются ассоциативные ядра. Происходит усиленное развитие таламуса и формируется таламонеокортикальная система интеграции.
Что касается гипофиза, то он образуется из эктодермы глотки первоначально в виде кармашкообразного выпячивания (карман Ратке), которое затем обособляется от стенки глотки и в виде пузырька примыкает к области воронки.
Эпителий стенок пузырька образует разветвленную железу. Затем просвет железы исчезает, но остаются тяжи от железистых клеток, окруженные большим количеством кровеносных сосудов. Еще позднее обособляется промежуточная доля гипофиза, непосредственно граничащая с полостью воронки. У наземных животных за счет стенки воронки возникает нервная часть гипофиза, состоящая из нервных клеток.
В онтогенезе структуры промежуточного мозга возникают из стенок второго мозгового пузыря (diencephalon) пятипузырной стадии развития головного мозга. Глазные пузырьки превращаются в глазные бокалы и глазные стебельки. Позже из них строятся сетчатка глаза и зрительные проводящие пути. Из боковых стенок второго пузыря формируются структуры дорсального (thalamus) и вентрального (subthalamus) таламусов. Верхней стенкой являются эпиталамические структуры, нижней – гипоталамус.
Из верхней части задней стенки забугорья все структуры образуются во время онтогенеза из крыловидной пластинки (пограничная борозда и основная пластинка заканчиваются на уровне среднемозгового пузыря). Здесь не возникают ни моторные (двигательные), ни вегетативные ядра, а, следовательно, нет и соответствующих черепно-мозговых нервов. Ядра промежуточного мозга являются либо сенсорными (переключательными), либо интегративными (ассоциативными). Имеются здесь и неспецифические ядра.
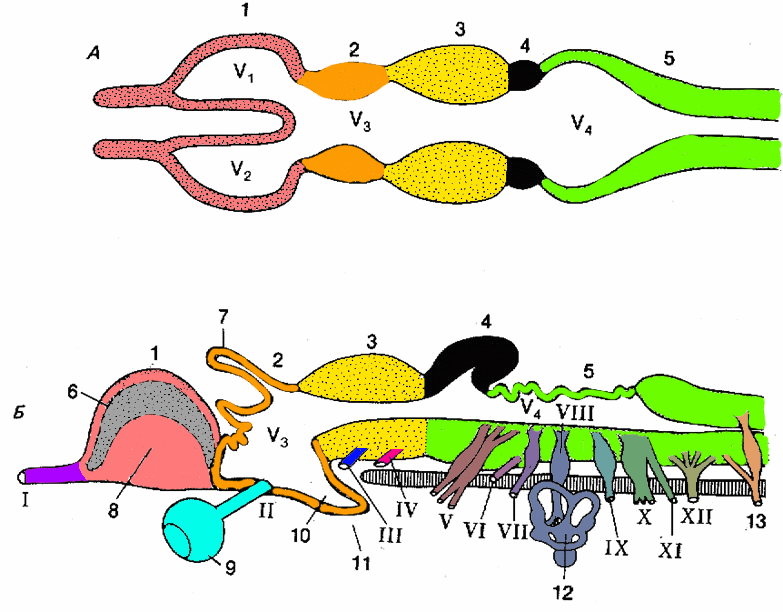 Рис.3.
Схема строения головного мозга низших
позвоночных (костистые рыбы), уже имеющих
все подразделения мозга: А – горизонтальный
разрез; Б – сагиттальный разрез, на
котором показано отхождение головных
нервов: 1 – передний мозг, продольный
срез проходит между парными передними
мозговыми пузырями (полушариями), однако
показано внутреннее строение правого
полушария: плащ (6) и базальный ганглий
(8); 2 – промежуточный мозг с дорсальным
выпячиванием, эпифизом (7) и вентральным
выпячиванием, воронкой (10), к которой
прилегает гипофиз (11), по бокам показаны
глаза (9), на горизонтальном срезе в
стенках промежуточного мозга можно
видеть зрительные бугры; 3 – средний
мозг; 4 – мозжечок, особо развито
дорсальное выпячивание; 5 – продолговатый
мозг с тонкой крышей над ромбовидной
ямкой. Внутри мозга находится система
полостей, местами расширяющихся в
желудочки мозга: V1, V2 – боковые желудочки
полушарий переднего мозга; V3 – третий
желудочек, полость промежуточного
мозга; V4 – четвертый желудочек. Головные
нервы: I – обонятельный, II – зрительный,
III – глазодвигательный, IV – блоковый,
V – тройничный, VI – отводящий, VII –
лицевой, VIII – равновесно-слуховой, IX –
языкоглоточный, Х – блуждающий, XI –
добавочный, XII – подъязычный; 12 – слуховой
лабиринт; 13 – первый спинномозговой
нерв.
Рис.3.
Схема строения головного мозга низших
позвоночных (костистые рыбы), уже имеющих
все подразделения мозга: А – горизонтальный
разрез; Б – сагиттальный разрез, на
котором показано отхождение головных
нервов: 1 – передний мозг, продольный
срез проходит между парными передними
мозговыми пузырями (полушариями), однако
показано внутреннее строение правого
полушария: плащ (6) и базальный ганглий
(8); 2 – промежуточный мозг с дорсальным
выпячиванием, эпифизом (7) и вентральным
выпячиванием, воронкой (10), к которой
прилегает гипофиз (11), по бокам показаны
глаза (9), на горизонтальном срезе в
стенках промежуточного мозга можно
видеть зрительные бугры; 3 – средний
мозг; 4 – мозжечок, особо развито
дорсальное выпячивание; 5 – продолговатый
мозг с тонкой крышей над ромбовидной
ямкой. Внутри мозга находится система
полостей, местами расширяющихся в
желудочки мозга: V1, V2 – боковые желудочки
полушарий переднего мозга; V3 – третий
желудочек, полость промежуточного
мозга; V4 – четвертый желудочек. Головные
нервы: I – обонятельный, II – зрительный,
III – глазодвигательный, IV – блоковый,
V – тройничный, VI – отводящий, VII –
лицевой, VIII – равновесно-слуховой, IX –
языкоглоточный, Х – блуждающий, XI –
добавочный, XII – подъязычный; 12 – слуховой
лабиринт; 13 – первый спинномозговой
нерв.
Гипоталамус отделяется от соседних участков мозга на 2-м месяце внутриутробной жизни. Вслед за этим начинается формирование шести гипоталамических ядер, которые представляют собой скопление нейронов с определенными функциями. Дифференцировка входящих в них клеток продолжается до 6-го месяца внутриутробной жизни, а завершается еще позднее. В четырех из шести ядер, вырабатываются гормоны, которые по системе сосудов направляются в аденогипофиз.
Гипоталамо-аденогипофизарную систему составляют супрахиазматиче-ское ядро, вентромедиальное, дорсомедиальное и аркуатное ядра. Сосудистая система появляется на 14-й неделе в виде первых капиллярных петель, и ее формирование завершается к моменту рождения. Синтезируемые в этих ядрах регуляторные пептиды обнаруживаются в аденогипофизе уже на 10-й неделе развития плода. Однако, по некоторым наблюдениям, в первые три месяца внутриутробной жизни, а возможно, до первой половины беременности гипофиз не подчиняется контролю гипоталамуса. Это объясняется незрелостью нейросекреторных клеток и недостаточным развитием портальной системы сосудов.
Вазопрессин появляется в гипофизе плода на 15-17-й, а окситоцин – на 18-19-й неделях внутриутробного развития. К 6-му месяцу беременности со-держание их значительно возрастает. Установление гипоталамического контроля над эндокринными железами происходит к концу внутриутробного развития.