

290 |
L. Scott et al. |
Table 13.1 Samples of Azokh Cave coprolites and deposits
Lab |
Unit |
Depth cm |
Type |
Age |
MIS |
no. |
|
|
|
|
|
5360 |
|
|
Modern soil |
|
|
|
|
|
outside |
|
|
5306 |
Unit |
80 |
Cave |
<200 yrs |
1 |
|
I |
|
sediment |
|
|
5246 |
Unit |
224, |
Coprolite # |
? |
5 |
|
II/I |
reworked, |
0804 |
|
|
|
|
in burrow |
|
|
|
5153 |
Unit |
233 |
Coprolite # |
c. |
5 |
|
II |
|
0812 |
100 kyrs |
|
5302 |
Unit |
238.5 |
Cave |
c. |
5 |
|
II |
|
sediment |
100 kyrs |
|
|
|
|
close to |
|
|
|
|
|
5153 |
|
|
5304 |
Unit |
278.5 |
Cave |
100–130 |
5 or |
|
II |
|
sediment |
kyrs |
6 |
5305 |
Unit |
337 |
Cave |
130– |
6 |
|
II or |
|
sediment |
160-kyrs |
|
|
III |
|
|
|
|
immersion magnification. Five sediment samples for comparison were also chemically processed in the same way as the coprolites for phytolith investigation (Table 13.1).
Criteria for the Identification of Phytolith Types
Nine different grass silica short cell (GSSC) phytoliths were classified morphologically following the International Code for Phytolith Nomenclature (ICPN Working Group et al. 2005). Four different morphological variations of the bilobate morphotype within the Poaceae are recognized in this study (Rossouw 2009). Variant 1 possesses orbicular lobes that are symmetrical in planar view, and it has a central portion or neck equal or greater than one third of total length of body. Variant 2 has a comparatively short central portion with orbicular to ovate lobes that are symmetrical in planar view and with the length of its central portion equal to or less than one third of total length of body. Variant 3 is always asymmetrical in planar view with the length of its central portion less than one third of total length of body. This type is comparable to the irregular complex dumbbell types recognized by Twiss et al. (1969) and the ‘Other lobate’ category in Fredlund and Tieszen (1994). The fourth variant, or Stipa-type bilobate, is a predominantly pooid morphotype, which appear trapezoidal or tabular in side view with generally ovate to scutiform lobes (Mulholland 1989; Fredlund and Tieszen 1994; Rossouw 2009).
Other GSSC-phytoliths that were identified include polylobates, commonly produced by the C3 Pooideae subfamily and C3 Panicoid species as well as cross and saddle morphotypes, which are primarily produced in the C3/C4 Panicoideae and C4 Chloridoideae subfamilies, respectively. Trapezoidal, rondel and oblong morphotypes are largely produced by the C3 Pooideae and Danthionoideae subfamilies. Trapezoids are six-sided, square or rectangular silica bodies with few sides parallel (Rossouw 2009). Planar margins are angular and not medially constricted. The trapezoid category is equivalent to types 1b, 1d and 1f in Twiss et al. (1969), the rondel types described by Mulholland (1989), the conical and pyramidal types in Fredlund and Tieszen (1994) and rondel types g, h and i in Thorn (2004). The rondel is cylindrical or semi-cylindrical, tapers distally, and resembles a truncated cone (Mulholland 1989). It is circular, elliptical or acicular in planar view (Rossouw 2009). This morphotype compares to type 1a in Twiss et al. (1969) and the conical type in Fredlund and Tieszen (1994). The oblong morphotype includes six-sided silica bodies that are at least twice as long as broad with parallel or nearly parallel sides. Oblong-shaped phytoliths are defined as having smooth, sinuous or crenate planar edges and trapezoidal cross-sections (Rossouw 2009). It corresponds with types 1c, 1g and 1h in Twiss et al. (1969), and the “longer forms with more polygonal cross-sections” in Mulholland (1989, p. 495). There are also elongated long-cell morphotypes as well as acicular (trichome) and bulliform morphotypes. All other unidenti- fied morphotypes, which represent a variety of plant families that include dicotyledons or gymnosperms, were classified as “indeterminate”. In this provisional study we did not attempt to identify this group because we did not have reference material from the local plants.
Results
Diatoms
Less than one diatom per gram was obtained from only one of the coprolites and a single diatom was found in 5 g of deposits of Unit I. The following species were among the diatoms recorded: Hantzschia amphioxys (Fig. 13.2a), Pinnularia borealis (Fig. 13.2b) and Nitzschia sp. (Fig. 13.2c). Both H. amphioxys and P. borealis are aerophilic taxa (preferring shallow or running water) and therefore indicative of moist conditions e.g., lake shores, ground water springs, wetlands or wet soils (Denham et al. 2009).
13 Coprolites and Plant Microfossils |
291 |
Fig. 13.2 Diatoms recorded in the two Azokh Cave coprolites,
Hantzschia amphioxys (44 µm) (a), Pinnularia borealis (31 µm) (b),
Nitzschia sp. (54 µm) (c) and Pliocaenicus (44 µm) (d)
A fourth diatom type represents Pliocaenicus (Fig. 13.2d), a genus which has been recorded from a Tertiary freshwater environment in China (Stachura-Suchoples and Jahn 2009). The diatom specimen from the Unit 1 is obscured by sediment but could tentatively be Achnanthes sp., a genus which may be found in a large variety of environments and therefore least informative.
Phytoliths
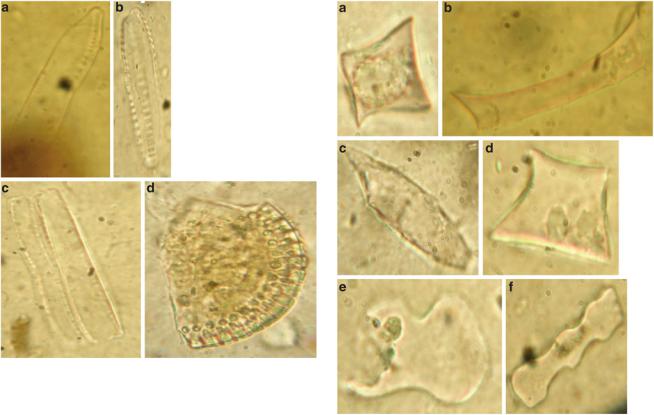
Both the coprolites and studied cave deposits were very rich in siliceous phytoliths ranging from well preserved to broken, etched and damaged. Phytoliths in the coprolite 5246 were more corroded than 5153, while those deposits of Unit II (5302) from which the latter was derived were better preserved in comparison with other levels. Examples of the recorded phytoliths are shown in Fig. 13.3a–f. Counts show that GSSC-phytoliths (grass silica short cell phytoliths) make up more than 60% of the total number of phytoliths counted in coprolite 5153, while 5246 has a lower percentage (Fig. 13.4a). The rest consist of indeterminate phytoliths of various plants which could include a variety of unidentified dicotyledonous and gymnosperm species and include hair bases (Fig. 13.3c), trichomes and phytoliths of leaves,
Fig. 13.3 Some phytolith types from the Azokh coprolites: Trapezoid (14 µm) (a); Oblong (32 µm) (b); Basal view of trichome (42 µm) (c); Rondel (20 µm) (d); Bilobate Variant 2 (24 µm) (e); Polylobate (26 µm) (f)
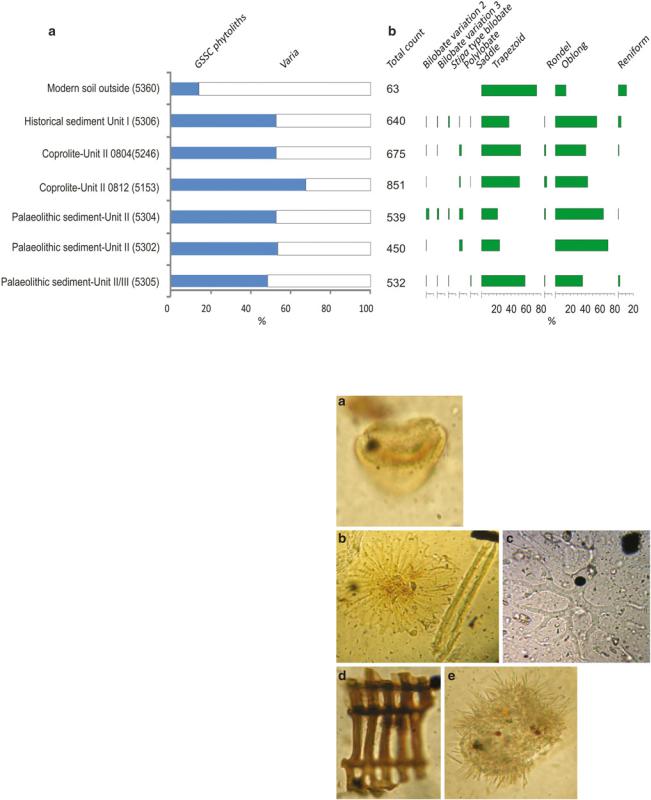
branches or fruits. In view of this high proportion of unidentified forms we cannot make an accurate reconstruction of the vegetation without further research on these types. The GSSC-phytoliths are composed of several types (Fig. 13.4b) that include comparatively few bilobate morphotypes (Fig. 13.3e) representing less than 2% of the GSSC-component. Polylobate morphotypes (Fig. 13.3f) account for less than 3% of the total number of GSSC-phytoliths counted. The highest frequencies of epidermal short cells in the coprolites are represented by trapezoidal, rondel and oblong morphotypes (totalling c. 90%) (Fig. 13.3 a, b and d).
Pollen and Other Microfossils
The coprolites were poor in pollen and there was not enough for determining past vegetation composition. Only two pollen grains and some possible spores were found in coprolite 5153. One is Asteraceae, most probably belonging to Artemisia (not illustrated) of which the morphology is
292 |
L. Scott et al. |
Fig. 13.4 Diagrams of phytoliths from Azok Cave surface soil, coprolites and deposits showing the ratio between GSSC (grass silica short cell) phytoliths and indeterminate phytoliths (a), and the percentages of different types of GSSC phytolith types (b)
well known, but Echinops or Centaurea cannot be excluded. The second pollen type (Fig. 13.5a) is as yet unidentified. Possible fern or bryophyte spores or algae also occur. The preservation of the two pollen grains was reasonable enough to reveal their morphological characteristics suggesting that their low concentration is possibly not a result of destructive processes but due to low pollen availability. Their brownish color suggests that they are indeed fossil and not modern contaminants.
Long silica structures (longer than 200 microns) were observed in coprolite 5153. They resemble sponge spicules (Fig. 13.5b) and are indicative of flooding or could have been derived from drinking water. In soils outside caves, this is indicative of flooding from a stream. Azokh 1 is seasonally wet (Murray et al. 2016; Marin-Monfort et al. 2016), and they may have formed at this time, but when the cave is dry, they may have entered the animal’s alimentary tract through drinking water. Also recorded in coprolite 5153 are comparatively large radial leaf trichomes of c. 174 and 200 micron diameters (Fig. 13.5b, c). Microscopic charcoal occurred in small numbers as brown and black woody plant remains (Fig. 13.5d) derived from occasional natural fires or fine aerial dust ingested accidentally inside or outside the cave from hearths. A spiny “corpuscle” (Fig. 13.5e) in coprolite 5153 is similar to structures that have been seen in hyena coprolites from sites in South Africa (L. Scott unpublished data), and in a picture of an unknown structure derived from a stool (http://www.dpd.cdc.gov/dpdx/html/ ImageLibrary/A-F/Artifacts/body_Artifacts_il6.htm).
Fig. 13.5 Other microfossil types in the Azokh coprolites, unidenti- fied pollen (22 µm) (a), trichome (174 µm) (left) and sponge spicule (218 µm) (right) (b), trichome (c. 200 µm) (c), woody fibre (38 µm) (d) and unidentified corpuscule (44 µm) (e)