

Другие органеллы скелетного мышечного волокна
Среди органелл общего значения в скелетном мышечном волокне хорошо развитая агранулярная (гладкая) эндоплазматическая сеть, которая называется саркоплазматическим ретикулумом (СПР). Саркоплазматический ретикулум при помощи ферментов накапливает ионы кальция. В некоторых участках плазмолемма мышечного волокна образует глубокие впячивания в виде трубочек, которые проходят перпендикулярно волокну через всю его толщину. Они называются Т-трубочками. Т-трубочки окружают каждую миофибриллу. К Т-трубочкам с обеих сторон подходят цистерны СПР – Т-цистерны. Вместе с Т-трубочками Т-цистерны образуют триады – особую мембранную систему. Триады играют важную роль в инициации мышечного сокращения.
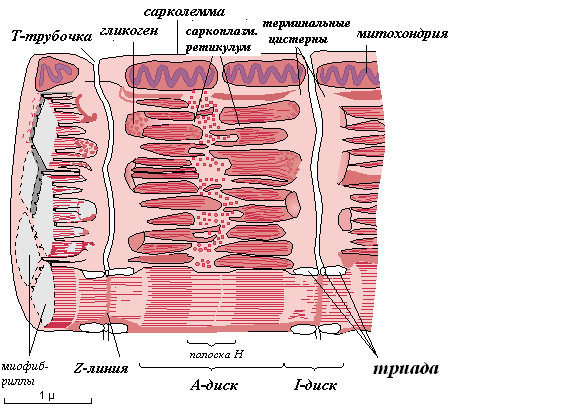
Мышечные волокна содержат также крупные митохондрии с многочисленными кристами.
Включения, характерные для скелетной мышечной ткани, - гликоген, липидные капли, которые используются для получения энергии, а также пигмент миоглобин. Миоглобин является железосодержащим пигментом, аналогичным гемоглобину, и способен связывать кислород.
Механизм мышечного сокращения
Общепринятой теорией является модель мышечного сокращения, предложенная Х.Хаксли (в 1954 г.). Это теория скольжения нитей. Суть этой теории в следующем:
нервный импульс проходит по нервному волокну и передаётся на плазмолемму мышечного волокна;
электрический импульс идёт по Т-трубочке вглубь мышечного волокна и передаётся на лежащие рядом Т-цистерны СПР;
мембраны СПР после деполяризации становятся проницаемыми для ионов;
ионы кальция выходят из Т-цистерны, связываются с молекулами ТnC;
при связывании ионов кальция изменяется конфигурация тропонина и на актиновых филаментах открываются активные центры для связывания головок миозина (которые в состоянии расслабления были закрыты TnI);
головки миозина начинают взаимодействуют с молекулами актина: они изгибаются в шарнирных областях и создают тянущиеся усилия;
это вызывает скольжение
Для возвращения головки миозина в исходное положение необходима энергия АТФ, которая распадается благодаря АТФ-азной активности миозина.
О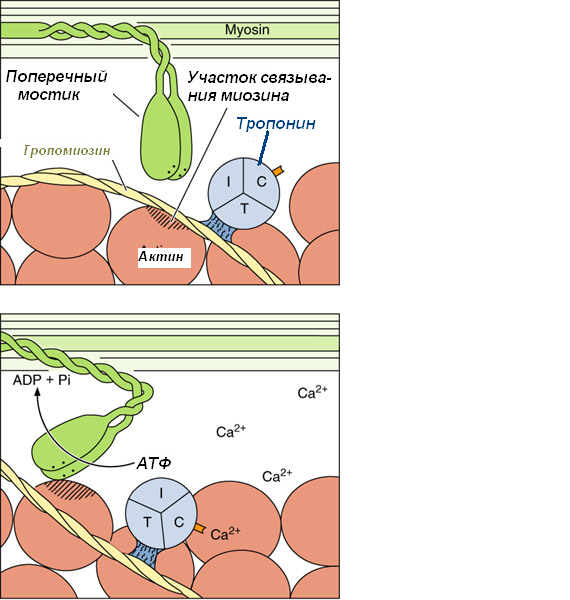 тдельное
мышечное сокращение является результатом
сотен циклов образования и разъединения
мостиков. Активное сокращение, которое
приводит к полному взаимному перекрыванию
между тонкими и толстыми филаментами,
продолжается до тех пор, пока не будут
удалены ионы Са.
тдельное
мышечное сокращение является результатом
сотен циклов образования и разъединения
мостиков. Активное сокращение, которое
приводит к полному взаимному перекрыванию
между тонкими и толстыми филаментами,
продолжается до тех пор, пока не будут
удалены ионы Са.
При отсутствии нервных импульсов ионы кальция вновь откачиваются в СПР, и тропонин-тропомиозиновый комплекс вновь закрывает участки связывания миозина на актине.
Длина филаментов при сокращении не изменяется, но поскольку актиновые филаменты заходят глубже между миозиновыми филаментами:
длина I-дисков уменьшается;
длина Н-полоски уменьшается, хотя длина А-диска остается неизменной;
Z-линии сближаются и уменьшается протяженность саркомера в целом.
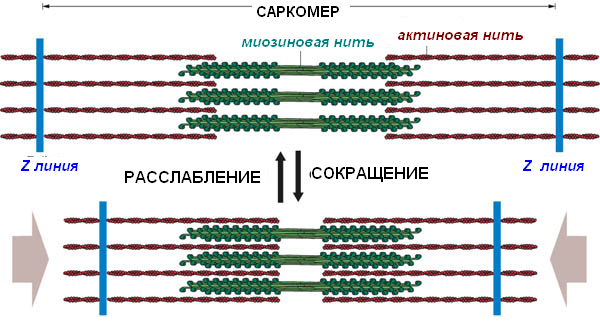
Конформационные изменения в сокращенной скелетной мышце
в сравнении с покоящейся мышцей
|
Сокращенная мышца |
Диск А |
Без изменений |
Диск I |
Укорачивается |
Полоска Н |
Укорачивается |
Z-линия |
Приближаются друг к другу |