

Сравнительная характеристика мышечных тканей
|
Тип мышечной ткани |
||
Поперечно-полосатая скелетная |
Поперечно-полосатая сердечная |
Гладкая |
|
Источник развития |
Миотомы мезодермы |
Миоэпикардиальная пластинка (висцеральный листок спланхнотома) |
Мезенхима |
Структурно-функциональная единица |
Мышечное волокно - миосимпласт |
Клетка - кардиомиоцит |
Клетка - гладкий миоцит |
Расположение ядер |
многочисленные, по периферии |
В центре (одно или два) |
В центре (одно) |
Исчерченность |
Актин и миозин формируют характерные полоски |
Актин и миозин формируют характерные полоски |
Характерных полосок нет |
Сократительный аппарат |
Миофибриллы |
Миофибриллы |
Миофиламенты |
Т-система |
Т-трубочки на уровне A-I дисков; триады |
Т-трубочки на уровне Z-линии, диады |
Нет Т-трубочек, нет триад и диад, есть кавеолы |
Z-линия |
есть |
есть |
Нет, имеются плотные тельца |
Кальций-связывающий белок |
тропонин |
тропонин |
кальмодулин |
Межклеточные соединения |
нет |
Вставочные диски – комплекс межклеточных контактов |
Щелевые контакты (нексусы) |
Источник регенерации |
Деление миосателлитоцитов, гипертрофия |
внутриклеточная гипертрофия |
Деление миоцитов, гипертрофия |
Источник иннервации |
Соматический отдел нервной системы |
Вегетативный отдел нервной системы |
Вегетативный отдел нервной системы |
Характер сокращения |
Тетанический произвольный |
Ритмический непроизвольный |
Тонический непроизвольный
|
Несократительные функции |
Участие в терморегуляции и углеводном обмене |
Синтез гормона – натрийуретического пептида (секреторные кардиомиоциты правого предсердия) |
Синтез эластических волокон, коллагена III типа |
Скелетная мышечная ткань развитие
Источником развития скелетной мышечной ткани являются миотомы сомитов. Стадии развития:
Миобластическая стадия. Клетки миотомов превращаются в миобласты и мигрируют к местам закладки мышц. Миобласты делятся митозом. Часть миобластов обособляется в виде миосателлитоцитов, которые сохраняют свойства малодифференцированных клеток до конца жизни.
Миосимпластическая стадия. Миобласты располагаются в виде цепочек и сливаются друг с другом. Образуются миосимпласты. В их цитоплазме образуются миофибриллы из сократительных белков, которые начинают синтезироваться ещё в миобластах. Миофибриллы лежат на периферии миосимласта, ядра занимают центральное положение.
Стадия миотубул. В симпластах увеличивается число миофибрилл. Длина их увеличивается.
Стадия зрелого мышечного волокна. В эту стадию объём миофибрилл увеличивается до такой степени, что они занимают основную массу волокна, смещаясь в центр и сдвигая ядра на периферию.
СТРОЕНИЕ МЫШЕЧНОГО ВОЛОКНА.
Мышечные волокна являются структурно-функциональным элементом скелетной мышечной ткани. Они имеют длину до 20-30 см, толщину около 100 мкм. Состоят из двух частей:
симпласта;
миосателлитоцитов.
Мышечное волокно снаружи покрыто сарколеммой. Сарколемма состоит из толстой базальной мембраны и плазмолеммы мышечного волокна. Между базальной мембраной и плазмолеммой в отдельных участках имеются углубления, в которых расположены миосателлитоциты. Миосателлитоциты – камбиальные клетки скелетной мышечной ткани. Миосателлитоцит – типичная одноядерная клетка, окруженная своей плазмолеммой, имеет слабо развитые органеллы.
Миосимпласт содержит множество (до несколько тысяч) ядер, лежащих на периферии волокна, под сарколеммой.
Протоплазму волокна называют саркоплазмой. В ней находятся органеллы общего значения (за исключением центриолей), органеллы специального значения – миофибриллы, и включения.

СТРОЕНИЕ МИОФИБРИЛЛ.
Миофибриллы, числом до двух тысяч в одном волокне, занимают основную часть волокна. Их длина равна длине волокна, диаметр до 2 мкм. В каждой миофибрилле при световой микроскопии обнаруживается исчерченность – чередование светлых и тёмных дисков. В поляризованном свете тёмные диски имеют двойное лучепреломление и поэтому тёмные диски называются анизотропными, или А-дисками. Светлые диски не имеют двойного лучепреломления и называются изотропными, или I-дисками.
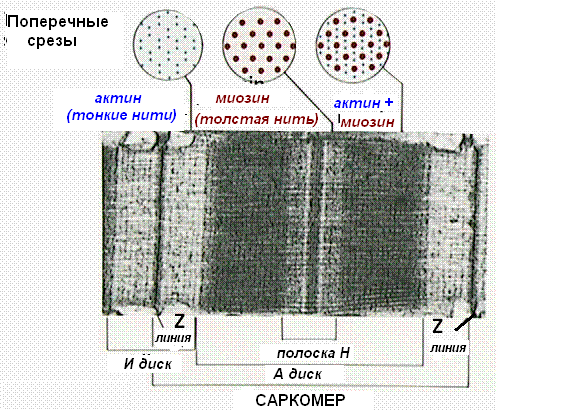
Посередине I-диска проходит тёмная полоска, которая называется Z-линией, или телофрагмой. На поперечном разрезе телофрагма представляет собой решётку, в узлах которой закрепляются актиновые филаменты.
В центре А-диска находится более светлая полоска Н, а посередине её проходит тёмная линия М, или мезофрагма.
Участок миофибриллы, лежащий между соседними Z-линиями, называется саркомером. Саркомер – структурно-функциональная единица миофибриллы. В состав саркомера последовательно входят:
Z-линия;
½ диска I;
диск А;
½ диска I
вторая Z-линия.
Каждый саркомер состоит из тонких актиновых и толстых миозиновых филаментов.
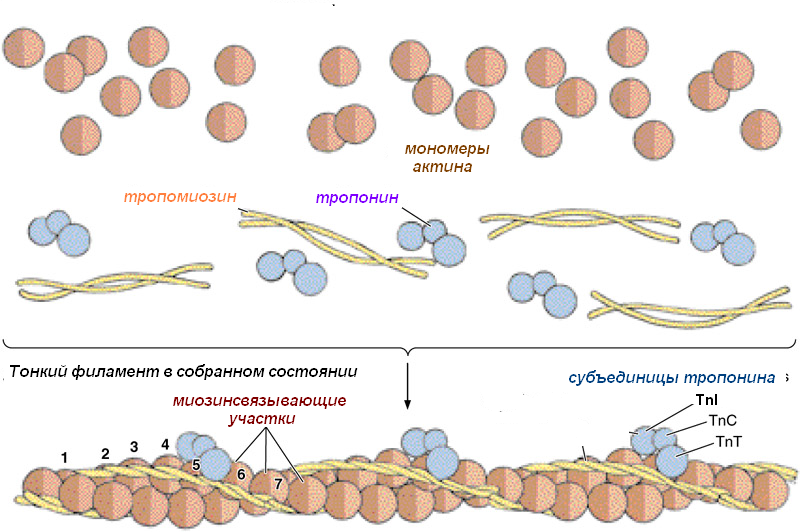
В составе тонких (диаметр 5 нм) актиновых филаментов входят белки:
актин;
тропонин;
тропомиозин.
Молекулы актина имеют глобулярное строение – G-актин. Эти молекулы соединяются вместе в длинные цепочки – фибриллярный, F-актин. В актиновых филаментах две цепи F-актина образуют двойную спираль. В бороздках между цепями спирали лежат молекулы тропомиозина. К молекулам тропомиозина на равных расстояниях друг от друга прикрепляются молекулы тропонина. Молекула тропонина состоит из трёх субъединиц: TnT, TnI, TnC. TnT осуществляет прикрепление тропонина к тропомиозину. TnC отвечает за связывание с ионами кальция. TnI препятствует взаимодействию миозина с актином.
Толстые филаменты (диаметр 12 нм) содержат белок миозин. Каждая молекула миозина состоит из двух частей: головки и хвоста и может сгибаться в двух местах – шарнирных участках. Головка миозина имеют АТФ-азную активность и способна расщеплять АТФ с образованием энергии. Молекулы миозина соединяются в пучки и образуют толстые миозиновые филаменты. По периферии толстых филаментов находятся участки, содержащие головки миозина. Центральная часть не содержит головок.
В составе саркомера толстые филаменты лежат только в диске А. Тонкие филаменты расположены в диске I, но концами частично заходят в диск А между миозиновыми филаментами. Та часть диска А, которая содержит и актиновые и миозиновые филаменты, выглядит на срезах более тёмной, а та его часть, которая содержит только миозиновые филаменты, светлее. Эта часть диска А, которая содержит только миозиновые филаменты, и составляет полоску Н. Таким образом:
диск I состоит из актиновых филаментов;
полоска Н диска А состоит из миозиновых филаментов;
на периферии диска А есть зона пересечения актиновых и миозиновых филаментов.
На поперечном срезе миофибриллы можно видеть, что в зоне пересечения вокруг одной толстой филаменты лежат шесть тонких филамент.
Тонкие филаменты неподвижно прикреплены к Z-линиям. В состав Z-линий входят белки α-актинин, десмин, виментин.
Линия М в центре Н-полоски – место соединения всех миозиновых филаментов друг с другом. В их скреплении участвуют белки миомезин и С-белок.