

Timin__Lektsii_po_biokhimii
.pdfbiokhimija.ru |
Тимин О.А. Лекции по биологической химии |
161 |
|
|
|
|
|
|
Р Е П ЛИ К АЦ И Я Д Н К
Синтез ДНК происходит не беспорядочно, а в строго определенный период жизни клетки. Всего выделяют 4 фазы клеточного цикла: митоз (М), синтетическую (S), пресинтетическую (G1, от англ. gap – интервал), постсинтетическую (G2).
Важное участие в регуляции смены фаз клеточного цикла занимают циклины – белки массой 35-90 кДа, уровень которых меняется в ходе клеточного цикла. По функции циклины – это активаторные субъединицы ферментов циклин-зависимых киназ (ЦЗК). Активные комплексы циклин-ЦЗК фосфорилируют внутриклеточные белки, изменяя их актив-
ность. Этим обеспечивается продвижение по клеточному циклу.
Синтез (репликация, удвоение) ДНК происходит в S-фазу клеточного цикла, когда клетка готовится к делению. Механизм репликации, как установили Мэтью Мезельсон и Франклин Сталь в 1957 г, полуконсервативный, т.е. на каждой нити материнской ДНК синтезируется дочерняя копия.
Как любой матричный биосинтез, репликация требует наличия нескольких компонен-
тов:
o матрица – в ее роли выступает материнская нить ДНК, o растущая цепь – дочерняя нить ДНК,
o субстраты для синтеза – dАТФ, dГТФ, dЦТФ, ТТФ, o источник энергии – dАТФ, dГТФ, dЦТФ, ТТФ,
o ферменты.
Синтез ДНК начинается в определенных участках, получивших название точка ori (англ. origin – начало). На каждой ДНК млекопитающих точек ori насчитывается около 100. Репликация распространяется от этих участков в обе стороны по нитям ДНК с образованием
репликативных "пузырей". В каждом таком "пузыре" имеются две репликативные "вил-
ки", в которых происходит расплетание, раскручивание и непосредственный синтез ДНК. При этом репликативные вилки удаляются друг от друга. В целом вся репликация ДНК у эукариот заканчивается за 9 часов.

Матричные биосинтезы |
162 |
|
|
Синтез новой цепи ДНК идет в направлении от 5'-конца к 3'-концу, т.е. 5'-конец новой ДНК остается свободным, следующие нуклеотиды присоединяются к 3'-гидроксильной группе предыдущего нуклеотида со скоростью порядка 100 нуклеотидов в секунду.
В репликативной вилке в направлении 5' 3' непрерывно (т.е. обычным заурядным присоединением последующих нуклеотидов к предыдущим через С3 и С5) синтезируется только одна нить, а именно та, для которой направление синтеза совпадает с направлением движения репликативной вилки и соответствует направлению материнской нити 3' 5'. По мере расплетания ДНК и движения репликативной вилки на этой материнской нити открываются участки, где возможно безостановочное удлинение ведущей дочерней нити.
Направление 5' 3' для другой дочерней нити ДНК противоположно движению репликативной вилки. Поэтому синтез этой отстающей нити (в направлении 5' 3') возможен только после расплетания части ДНК и освобождения участка для синтеза.

biokhimija.ru |
Тимин О.А. Лекции по биологической химии |
163 |
|
|
|
Таким образом, синтез дочерней ДНК идет фрагментарно. По имени японского исследователя синтезируемые на отстающей цепи отрезки ДНК назвали фрагменты Оказаки.
В хронологическом порядке события репликации развертываются примерно следующим образом:
1.ДНК-топоизомеразы, находясь перед репликативной вилкой, разрезают молекулу ДНК для облегчения ее расплетания и раскручивания.
2.ДНК-хеликазы, следуя за топоизомеразами, раскручивают и расплетают молекулу ДНК.
3.ДНК-связывающие белки (ДСБ) связывают расплетенные нити ДНК и стабилизируют их, не допуская обратного "слипания" друг с другом.
4.ДНК-полимераза (греч.: – дельта), согласовано со скоростью движения репликативной вилки, осуществляет элонгацию ведущей цепи дочерней ДНК в направлении 5' 3' на матрице одной из нитей материнской ДНК (скорость 100 пар нуклеотидов в секунду).
5.ДНК-полимераза присоединяется к другой нити ДНК сразу после расплетания и в направлении 5' 3' синтезирует праймер (РНК-затравку) – последовательность РНК на матрице ДНК длиной от 10 до 200 нуклеотидов. После этого фермент удаляется с нити ДНК.
6.ДНК-полимераза проводит синтез фрагмента (длина 150-200 нуклеотидов) отстающей цепи дочерней ДНК в продолжение праймера. Она работает до тех пор, пока не встретит праймер предыдущего фрагмента Оказаки (синтезированного ранее). После этого данный фермент удаляется с цепи.
7.ДНК-полимераза встает вместо ДНК-полимеразы , движется в том же направлении (5' 3') и удаляет рибонуклеотиды праймера, одновременно встраивая дезоксирибонуклеотиды на их место. Фермент работает до полного удаления праймера, после чего сходит с цепи.
8.ДНК-лигаза производит сшивку отрезка, синтезированного ДНК-полимеразой , и участка отстающей цепи дочерней ДНК, встроенного ДНК-полимеразой , (фрагментов Ока-
заки).

Матричные биосинтезы |
164 |
|
|
|
|
П О В Р Е ЖД Е Н И Я И Р Е П АР А Ц И Я Д Н К
Так как на геном любой неделящейся клетки постоянно оказывает влияние окружающая среда, то вполне вероятны повреждения в составе нуклеотида, также возможно встраивание неправильного нуклеотида при репликации. Такие нарушения быстро определяются специальными ферментами, пораженный участок удаляется экзонуклеазами, заполняется
ДНК-полимеразой и сшивается ДНК-лигазой.
В случае изменения структуры основания (например, его дезаминирование) это основание удаляется ДНК-N-гликозидазой, затем другими ферментами удаляется дезоксирибоза и на ее место ДНК-полимеразой и ДНК-лигазой встраивается нужный нуклеотид.
Отрыв пуриновых и пиримидиновых оснований от дезоксирибозы устраняется ДНК-инсертазами, которые присоединяют к оставшейся дезоксирибозе соответствующие основания.
Р Е Г У Л Я Ц И Я Р Е П Л И К А Ц И И
Лекарственная регуляция
1.Дауномицин и актиномицин D интеркалируют, т.е. встраиваются между нитей ДНК и препятствуют продвижению ферментов репликации.
2.Новобиоцин снижает активность ДНК-гиразы у бактерий, подавляя их размножение.
3.Мелфалан и циклофосфамид превращаются в организме в соединения, которые вызывают присоединение группы CH3 или C2H5 к атому N7 гуанозина обеих цепочек молекулы ДНК, что приводит к перекрестной сшивке двух спиралей ДНК и остановке репликации.
Г И Б Р И Д И З АЦ И Я Д Н К - Д НК И Д Н К - Р Н К
Если нагреть раствор ДНК выше температуры 90С или сдвинуть рН в резко щелочную или резко кислую стороны, то водородные связи между нитями ДНК разрушаются и двойная спираль расплетается. Происходит денатурация ДНК или, по-другому, плавление. Если удалить агрессивный фактор, то происходит ренатурация или отжиг. При отжиге нити ДНК "отыскивают" комплементарные участки друг у друга и, в конце концов, вновь сворачиваются в двойную спираль.
Если в одной "пробирке" провести плавление и отжиг смеси ДНК человека и мыши, то некоторые участки цепей ДНК мыши будут воссоединяться с комплементарными участками цепей ДНК человека с образованием гибридов. Число таких участков зависит от степени родства видов. Чем ближе виды между собой, тем больше участков комплементарности нитей ДНК. Это явление называется гибридизация ДНК-ДНК.
Если в растворе присутствует РНК, то можно осуществить гибридизацию ДНК-РНК. Такая гибридизация помогает установить близость определенных последовательностей ДНК с какой-либо РНК.
Гибридизация ДНК-ДНК и ДНК-РНК используется как эффективное средство в молекулярной генетике.
Т Р АН С К Р И П Ц И Я
Прежде чем начнут синтезироваться белки, информацию об их строении необходимо "достать" из ДНК и доставить ее к месту синтеза белков. Этим занимаются информационные или матричные РНК. Одновременно клетке нужны транспортеры аминокислот –
biokhimija.ru |
Тимин О.А. Лекции по биологической химии |
165 |
|
|
|
транспортные РНК и структурные компоненты органелл, синтезирующих белок, – рибосомальные РНК. Вся информация о строении транспортных и рибосомальных РНК также находится в ДНК.
Поэтому существует процесс переписывания или транскрипции данных с ДНК на РНК (англ. transcription – переписывание) – биосинтез РНК на матрице ДНК.
Как в любом матричном биосинтезе в транскрипции выделяют 5 необходимых элемен-
тов:
o матрица – одна из цепей ДНК, o растущая цепь – РНК,
o субстрат для синтеза – рибонуклеотиды (УТФ, ГТФ, ЦТФ, АТФ), o источник энергии – УТФ, ГТФ, ЦТФ, АТФ.
o ферменты РНК-полимеразы и белковые факторы транскрипции.
Биосинтез РНК происходит в участке ДНК, который называется транскриптон, с одного края он ограничен промотором (начало), с другого – терминатором (конец).
РНК-полимеразы эукариот имеют по две больших субъединицы и несколько малых субъединиц.
С Т А Д И И Т Р А Н С К Р И П Ц И И
Выделяют три стадии транскрипции: инициация, элонгация и терминация.
ИНИЦИАЦИЯ
Промотор содержит стартовый сигнал транскрипции – ТАТА-бокс – определенную последовательность нуклеотидов ДНК, связывающую первый фактор инициации ТАТА-фактор. Этот ТАТА-фактор обеспечивает присоединение РНК-полимеразы к той нити ДНК, которая будет использоваться в качестве шаблона для транскрипции (матричная нить ДНК). Так как промотор ассиметричен ("ТАТА"), то он связывает РНК-полимеразу только в одной ориентации, что определяет направление транскрипции от 5'-конца к 3'-концу (5' 3'). Для связывания РНК-полимеразы с промотором необходим еще один фактор инициации – -фактор (греч. – "сигма"), но сразу после синтеза затравочного фрагмента РНК (длиной 8-10 рибонуклеотидов) -фактор отрывается от фермента.
Другие факторы инициации раскручивают спираль ДНК перед РНК-полимеразой.
ЭЛОНГАЦИЯ
Белковые факторы элонгации обеспечивают продвижение РНК-полимеразы вдоль ДНК и расплетают молекулу ДНК на протяжении примерно 17 нуклеотидных пар. РНК-полимераза продвигается со скоростью 40-50 нуклеотидов в секунду в направлении 5' 3'. Фермент использует АТФ, ГТФ, ЦТФ, УТФ одновременно в качестве субстрата и в качестве источника энергии.

Матричные биосинтезы |
166 |
|
|
ТЕРМИНАЦИЯ
РНК-полимераза остановится, когда достигнет терминирующих кодонов. С помощью белкового фактора терминации, так называемого -фактора (греч. – "ро"), от матрицы ДНК отделяются фермент и синтезированная молекула РНК, которая является первичным транскриптом, предшественником мРНК или тРНК или рРНК.
П Р О Ц Е С С И Н Г Р Н К
Сразу после синтеза первичные транскрипты РНК по разным причинам еще не имеют активности, являются "незрелыми" и в дальнейшем претерпевают ряд изменений, которые называются процессинг. У эукариот процессингу подвергаются все виды пре-РНК, у прокариот – только предшественники рРНК и тРНК.
ПРОЦЕССИНГ ПРЕДШЕСТВЕННИКА РРНК
Предшественники рРНК являются более крупными молекулами по сравнению со зрелыми рРНК. Их созревание сводится к разрезанию прерибосомной РНК на более мелкие формы, которые уже непосредственно участвуют в формировании рибосомы. У эукариот существуют 5S-, 5,8S-, 18S-, и 28S-рРНК. При этом 5S-рРНК синтезируется отдельно, а большая прерибосомная 45S-РНК расщепляется специфичными нуклеазами с образованием 5,8S-рРНК, 18S-рРНК, и 28S-рРНК.
Упрокариот молекулы рибосомальной РНК совсем иные по своим свойствам (5S-, 16S-
,23S-рРНК), что является основой изобретения и использования ряда антибиотиков в медицине
ПРОЦЕССИНГ ПРЕДШЕСТВЕННИКА МРНК
При транскрипции участков ДНК, несущих информацию о белках, образуются гетерогенные ядерные РНК, по размеру намного превосходящие мРНК. Дело в том, что из-за мо-

biokhimija.ru |
Тимин О.А. Лекции по биологической химии |
167 |
|
|
|
заичной структуры генов эти гетерогенные РНК включают в себя информативные (экзоны)
инеинформативные (интроны) участки.
1.Сплайсинг (англ. splice – склеивать встык) – особый процесс, в котором при участии малых ядерных РНК происходит удаление интронов и сохранение экзонов.
2. Кэпирование (англ. cap – шапка) – происходит еще во время транскрипции. Процесс состоит в присоединении к 5'-трифосфату концевого нуклеотида пре-мРНК 5'-углерода N7-метил-гуанозина.
"Кэп" необходим для защиты молекулы РНК от экзонуклеаз, работающих с 5'-конца, а также для связывания мРНК с рибосомой и для начала трансляции.
3. Полиаденилирование – при помощи полиаденилат-полимеразы с использованием молекул АТФ происходит присоединение к 3'-концу РНК от 100 до 200 адениловых нуклеотидов, формирующих поли (А)-хвост. Поли (А)-хвост необходим для защиты молекулы РНК от экзонуклеаз, работающих с 3'-конца.
ПРОЦЕССИНГ ПРЕДШЕСТВЕННИКА ТРНК
1.Формирование на 3'-конце последовательности Ц-Ц-А. Для этого у одних пре-тРНК с 3'-конца удаляются лишние нуклеотиды до "обнажения" триплета Ц-Ц-А, у других идет присоединение этой последовательности.

Матричные биосинтезы |
168 |
|
|
2.Формирование антикодоновой петли происходит путем сплайсинга и удаления интрона в средней части пре-тРНК.
3.Модификация нуклеотидов в молекуле путем дезаминирования, метилирования, восстановления. Например, образование псевдоуридина и дигидроуридина.
Р Е Г У Л Я Ц И Я Т Р А Н С К Р И П Ц И И
Так как транскрипция связывает ядро – "мозг" клетки, ее "банк знаний" и белки, "рабочих лошадок" клетки, то от качества и активности транскрипции зависит объем синтеза тех или иных белков, жизнедеятельность клетки, ее способность адаптироваться к окружающей обстановке.
У прокариот и эукариот регуляция транскрипции происходит, естественно, по-разному, хотя некоторые моменты похожи.
ПРОКАРИОТЫ
Регуляция биосинтеза белка у прокариот осуществляется на уровне транскрипции мРНК. В настоящее время принята теория оперона, сформулированная Франсуа Жакобом и Жаком Моно. В основе теории лежат следующие понятия:
o конституитивные ферменты – те, которые присутствуют в клетках всегда, независимо от ее активности,
o индуцибельные ферменты – те, которые синтезируются при появлении субстрата (структурные гены),
o оперон – группа тесно связанных между собой генов (несколько структурных генов и один ген-оператор), которые регулируют образование ферментов в организме,
o ген-регулятор – ген, регулирующий работу оперона, но не входящий в его состав. Он синтезирует белок-регулятор (чаще называемый белок-репрессор), который может быть в активной или неактивной форме.
biokhimija.ru Тимин О.А. Лекции по биологической химии 169
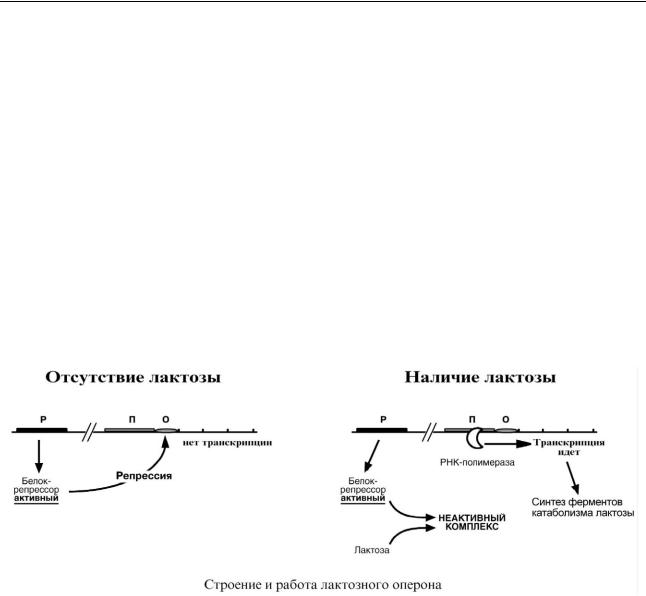
Лактозный оперон
Лактозный оперон в целом отвечает за катаболизм лактозы.
При изучении E.coli было замечено, что в клетке может быть две взаимоисключающие ситуации:
o активность одного из ферментов катаболизма лактозы низка, если в среде имеется глюкоза.
o но активность этого фермента резко повышается в обратной ситуации, т.е. при от-
сутствии глюкозы и при наличии лактозы.
На основании этих наблюдений была предложена схема регуляции оперона по меха-
низму индукции:
1.В отсутствие лактозы активный белок-репрессор связывается с оператором и блокирует синтез мРНК, кодирующей ферменты катаболизма лактозы. В результате эти ферменты
не образуются.
2.Если глюкозы нет, а лактоза есть, то последняя связывается с белком-репрессором и ингибирует его, не давая связаться с геном-оператором. Это позволяет РНК-полимеразе считывать информацию, отвечающую за синтез ферментов катаболизма лактозы, и синтезировать мРНК.
Таким образом, лактоза является индуктором транскрипции.
Триптофановый оперон
Триптофановый оперон в целом отвечает за синтез триптофана.
Функционирование триптофанового оперона в некотором смысле противоположно лактозному. Регуляция осуществляется по механизму репрессии.
1.В отличие от лактозного оперона, белок-репрессор синтезируется в неактивном состоянии и не может заблокировать транскрипцию генов, кодирующих ферменты синтеза триптофана. Синтез этой аминокислоты будет в клетке продолжаться до тех пор, пока в питательной среде не появится триптофан.
2.Триптофан соединяется с белком-репрессором и активирует его. Далее такой активный комплекс присоединяется к гену-оператору и блокирует транскрипцию. Таким образом, при наличии триптофана в среде прекращается его внутриклеточный синтез, экономятся ресурсы и энергия бактериальной клетки.
Вэтом случае триптофан является репрессором транскрипции.

Матричные биосинтезы |
170 |
|
|
ЭУКАРИОТЫ
Внутриклеточная регуляция
Существенное усложнение эукариотических организмов повлекло за собой появление новых способов регуляции активности транскрипции.
Амплификация – это увеличение количества генов, точнее многократное копирование одного гена. Естественно, все полученные копии равнозначны и одинаково активно обеспечивают транскрипцию.
Энхансеры (англ. to enhance усиливать) – это участки ДНК в 10-20 пар оснований, способные значительно усиливать экспрессию генов той же ДНК. В отличие от промоторов они значительно удалены от транскрипционного участка и могут располагаться от него в любом направлении (к 5'-концу или к 3'-концу). Сами энхансеры не кодируют какие-либо белки, но способны связываться с регуляторными белками (подавляющими транскрипцию).
Сайленсеры (англ. silence – молчание) – участки ДНК, в принципе схожие с энхансерами, но они способны замедлять транскрипцию генов, связываясь с регуляторными белками (которые ее активируют).
Перестройка генов. К подобным процессам относится кроссинговер – обмен участками гомологичных хромосом, и более сложный процесс – сайт-специфичная рекомбинация, которая изменяет положение и порядок нуклеотидных последовательностей в геноме.
Нуклеотидные последовательности, кодирующие одну белковую молекулу, могут быть разделены на отдельные сегменты, не связанные между собой. При дифференцировке В-лимфоцитов значительно удаленные сегменты генетического материала переносятся и группируются рядом. Затем на их основе синтезируется единая мРНК.
Процессинг мРНК – некоторые пре-мРНК подвергаются разным вариантам сплайсинга (альтернативный сплайсинг) в результате чего образуются разные мРНК, и соответственно, белки с разной функцией.
Примером может служить образование двух типов тяжелых цепей IgM в В-лимфоцитах, один из которых удерживает IgM на мембране, другой позволяет антителу нормально секретироваться наружу.
Изменение стабильности мРНК – чем выше продолжительность жизни мРНК в цитозоле клетки, тем больше синтезируется соответствующего белка.
Установлено, что при наличии пролактина в клетках молочной железы время полужизни мРНК белка казеина значительно увеличивается, а эстрадиол продлевает время полужизни мРНК белка вителлогенина в десятки раз.