

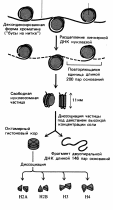
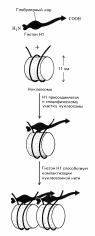
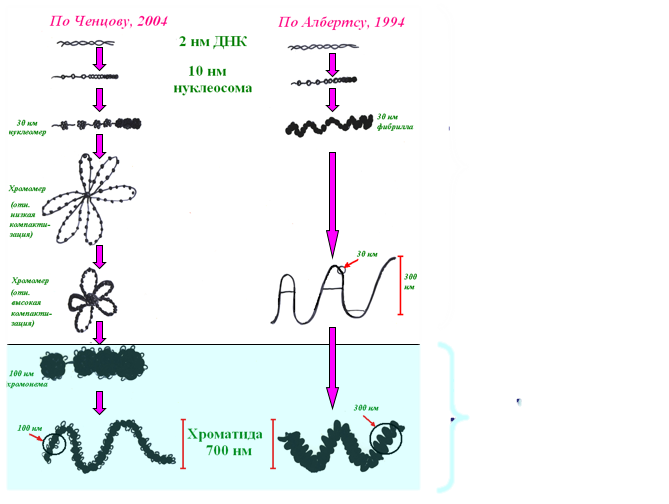
Уровни суперспирализации днк в митотической хромосоме.
Нуклеосома => Фибрилла => Нуклеомер => Петлевые домены => Хромомеры

ЭКЗАМЕНАЦИОННЫЙ БИЛЕТ №6
Транспортная функция плазмалеммы. Пассивный и активный транспорт.
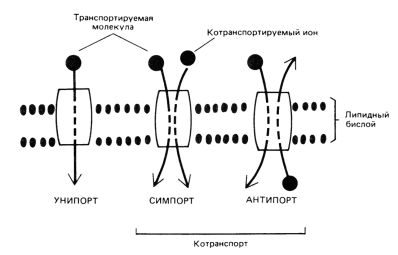
Существует 2 основных механизма транспорта через мембрану малых молекул: пассивный и активный транспорт. В случае пассивного транспорта перенос веществ происходит по градиенту концентрации и не требует затрат энергии. В случае активного транспорта все наоборот: транспорт происходит против градиента концентрации и для него необходима энергия.
 Транспортные
белки различаются по:
Транспортные
белки различаются по:
-специфичности
-направлению транспортировки
-механизму работы
По механизму работы транспортные белки разделяются на каналообразующие белки и белки-переносчики.
Каналообразующие белки – трансмембранные белки, пронизывающие мембрану несколько раз насквозь, в результате чего внутри образуется полость, изолирующая переносимое вещество от липидного бислоя, что и обеспечивает саму транспортировку.
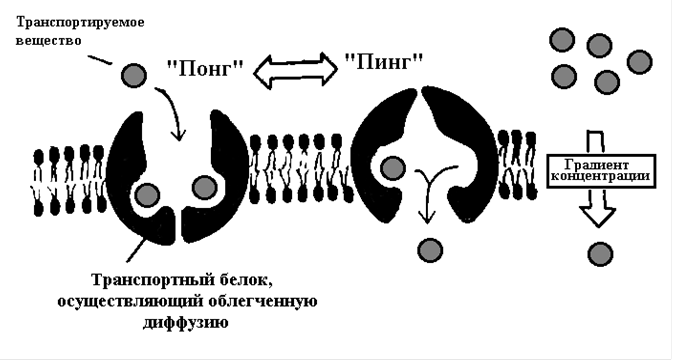 Принцип
работы белков-переносчиков
получил название «пинг-понг».
Принцип
работы белков-переносчиков
получил название «пинг-понг».
Таким
образом, пассивный
транспорт
может осуществляться путем простой
диффузии
(физическая диффузия через липидный
бислой, диффузия через каналообразующие
белки) и путем
облегченной диффузии
(с помощью белков-переносчиков).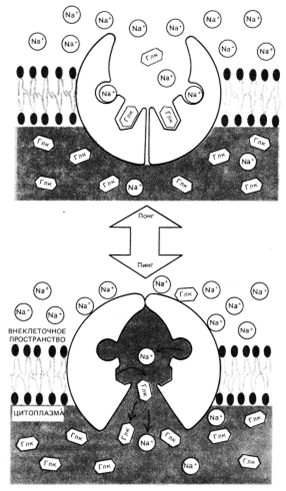
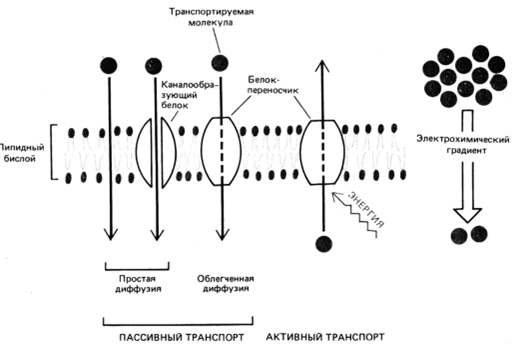 Активный
транспорт
происходит также с помощью
белков-переносчиков.
Но для их работы необходима энергия
(АТФ или протонного градиента).
Активный
транспорт
происходит также с помощью
белков-переносчиков.
Но для их работы необходима энергия
(АТФ или протонного градиента).
В ходе работы К+/Na+ насоса происходят конформационные изменения, напоминающие принцип “пинг-понг” за исключением того, что переходы индуцируются фосфолирированием и дефосфолирированием белка. На обеспечение работы этого насоса тратится довольно много энергии.
Транспорт больших молекул осуществляется принципиально другим способом (по сравнению с транспортом малых молекул)– путем везикулярного переноса. При этом транспортируемые вещества упаковываются в пузырьки – везикулы – и выводятся из клетки или поглощаются клеткой. Этот процесс энергозависим, поэтому его можно отнести к активному транспорту.
В зависимости от направления транспортировки различают эндоцитоз (транспорт в клетку) и экзоцитоз (из клетки).
Эндоцитоз в свою очередь подразделяют на пино – и фагоцитоз в зависимости от величины поглощаемых частиц.
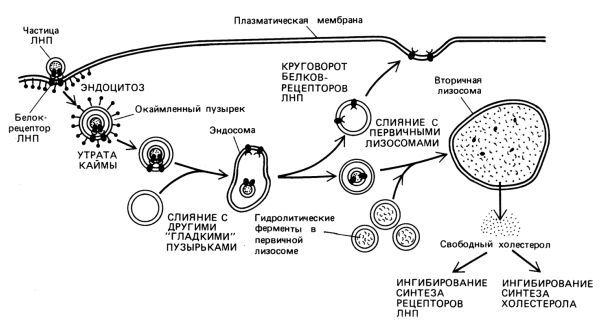
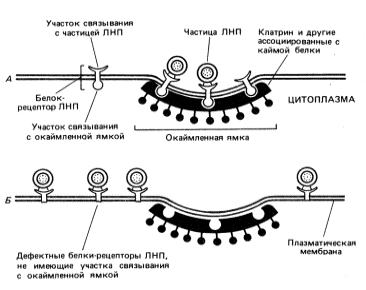 Эндоцитоз,
включая пино- и фагоцитоз, может быть
неспецифическим
(постоянным, конститутивным) и
специфическим
(рецепторным).
Эндоцитоз
(как специфический, так и неспецифический)
происходит не по всей клеточной
поверхности, а только в определенных
участках. Эти специализированные участки
называют окаймленными
ямками.
Эндоцитоз,
включая пино- и фагоцитоз, может быть
неспецифическим
(постоянным, конститутивным) и
специфическим
(рецепторным).
Эндоцитоз
(как специфический, так и неспецифический)
происходит не по всей клеточной
поверхности, а только в определенных
участках. Эти специализированные участки
называют окаймленными
ямками.
С экзоцитозом связано выделение синтезированных в клетке разнообразных веществ. Экзоцитоз может происходить постоянно и независимо от внешней среды (секреция коллагена, фибронектина фибробластами) или в ответ на внешний сигнал (нервный импульс, воздействие гормона и др.).
Эу- и гетерохроматин. Механизмы контроля экспрессии генов на уровне структурной организации хроматина
Еще в начале 30-х годов было замечено Э. Гейтцем, что в интерфазных ядрах существуют постоянные участки конденсированного хроматина, наличие которого не зависит от степени дифференцированнности ткани или от функциональной активности клеток. Такие участки получили название гетерохроматина, в отличие от остальной массы хроматина – эухроматина (собственно хроматина). В составе ядерного хроматина только лишь некоторые участки практически никогда не теряют особого конденсированного состояния. Такими постоянно конденсированными зонами чаще всего являются центромерные и теломерные участки хромосом.
Такие постоянно конденсированные участки хромосом в интерфазных ядрах сейчас принято называть конститутивным (постоянным, структурным) гетерохроматином. Здесь же необходимо отметить, что участки конститутивного гетерохроматина обладают целым рядом особенностей, которые отличают его от остального хроматина. Конститутивный гетерохроматин генетически не активен, он не транскрибируется, реплицируется он позже всего остального хроматина; он локализован в центромерных, теломерных и интеркалярных зонах митотических хромосом.
Вся остальная, основная масса хроматина ядра может менять степень своей компактизации в зависимости от функциональной активности, она относится к эухроматину. Эухроматические неактивные участки, которые находятся в конденсированном состоянии, стали называть факультативным гетерохроматином, подчеркивая необязательность такого его состояния. Хорошим примером факультативного гетерохроматина может служить X-хромосома в организме человека. В клетках мужской особи X-хромосома деконденсирована, она активна, транскрибируется и морфологически не выявляется из-за своего рыхлого, диффузного состояния. В клетках женского организма, где присутствуют две X-хромосомы, одна из них находится в активном, диффузном состоянии, а вторая – в неактивном, конденсированном, она временно гетерохроматизована. В этом состоянии она может существовать в течение всей жизни организма. Но потомки ее, попадая в клетки мужского организма следующего поколения, снова будут активированы.
Эухроматин (разрыхленный) - имеет менее компактную организацию, деспирализуется в конце митоза, образует слабоокрашенные нитчатые структуры содержит структурные транскрибируемые гены.