

Экзаменационный билет №2
История развития и основные положения клеточной теории.
Роберт Гук (1665) первым наблюдал с помощью увеличительных линз подразделение тканей пробки на «ячейки», или «клетки». Его описания послужили толчком для появления систематических исследований анатомии растений (Мальпиги, 1671; Грю, 1671), которые подтвердили наблюдения Роберта Гука и показали, что разнообразные части растений состоят из тесно расположенных «пузырьков», или «мешочков». Позднее А. Левенгук (1680) открыл мир одноклеточных организмов и впервые увидел клетки животных (эритроциты). Позднее клетки животных были описаны Ф. Фонтана (1781); но эти и другие многочисленные исследования не привели в то время к пониманию универсальности клеточного строения, к четким представлениям о том, что же являет собой клетка. Прогресс в изучении микроанатомии и клетки связан с развитие микроскопирования в XIX в. К этому времени изменились представления о строении клеток: главным в организации клетки стала считаться не клеточная стенка, а собственно ее содержимое, протоплазма (Пуркиня, 1830). В протоплазме был открыт постоянный компонент клетки – ядро (Браун, 1833). Все эти многочисленные наблюдения позволили Т. Шванну в 1838 г. сделать ряд обобщений. Он показал, что клетки растений и животных принципиально сходны между собой (гомологичны). Дальнейшее развитие эти представления получили в работах Р. Вирхова (1858).
В настоящее время клеточная теория постулирует:
Клетка – элементарная единица живого: – вне клетки нет жизни.
Клетка – единая система, состоящая из множества закономерно связанных друг с другом элементов, представляющих собой определенное целостное образование, состоящее из сопряженных функциональных единиц – органелл или органоидов.
Клетки сходны – гомологичны – по строению и по основным свойствам.
Клетки увеличиваются в числе путем деления исходной клетки после удвоения ее генетического материала (ДНК): клетка от клетки.
Многоклеточный организм представляет собой новую систему, сложный ансамбль из множества клеток, объединенных и интегрированных в системы тканей и органов, связанных друг с другом с помощью химических факторов, гуморальных и нервных (молекулярная регуляция).
Клетки многоклеточных организмов тотипотентны, т.е. обладают генетическими потенциями всех клеток данного организма, равнозначны по генетической информации, но отличаются друг от друга разной экспрессией (работой) различных генов, что приводит к их морфологическому и функциональному разнообразию – к дифференцировке.
Строение и функции постоянных межклеточных контактов.
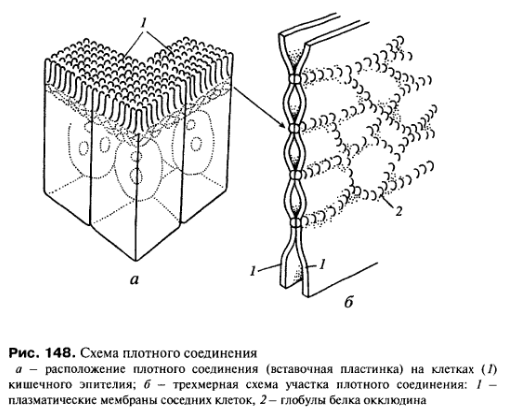 Запирающее
или плотное
соединение
характерно для однослойных эпителиев.
Это зона, где внешние слои двух
плазматических мембран максимально
сближены. На плоскостных препаратах
разломов плазматической мембраны в
зоне плотного контакта с помощью метода
замораживания и скалывания было
обнаружено, что точки соприкосновения
мембран представляют собой ряды глобул.
Это белки окклудин и клаудин, специальные
интегральные белки плазматической
мембраны, встроенные рядами. Такие ряды
глобул или полоски могут пересекаться
так, что образуют на поверхности скола
как бы решетку или сеть. Таким образом,
каждая клетка пласта как бы обведена
лентой этого контакта. Такие структуры
при специальных окрасках можно видеть
и в световом микроскопе. Они получили
у морфологов название замыкающих
пластинок.
Запирающее
или плотное
соединение
характерно для однослойных эпителиев.
Это зона, где внешние слои двух
плазматических мембран максимально
сближены. На плоскостных препаратах
разломов плазматической мембраны в
зоне плотного контакта с помощью метода
замораживания и скалывания было
обнаружено, что точки соприкосновения
мембран представляют собой ряды глобул.
Это белки окклудин и клаудин, специальные
интегральные белки плазматической
мембраны, встроенные рядами. Такие ряды
глобул или полоски могут пересекаться
так, что образуют на поверхности скола
как бы решетку или сеть. Таким образом,
каждая клетка пласта как бы обведена
лентой этого контакта. Такие структуры
при специальных окрасках можно видеть
и в световом микроскопе. Они получили
у морфологов название замыкающих
пластинок.
Таким образом, плотные контакты являются барьерами не только для макромолекул, но и непроницаемы для жидкостей и ионов.
Замыкающий, или плотный, контакт встречается между всеми типами однослойного эпителия (эндотелий, мезотелий, эпендима).
З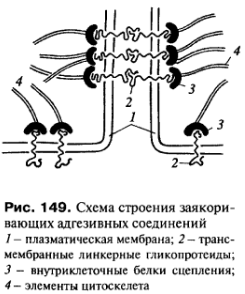 аякоривающие
или сцепляющие
соединения
или контакты так называются из-за того,
что они соединяют не только плазматические
мембраны соседних клеток, но и связываются
с фибриллярными элементами цитоскелета
(рис.
149). Для этого рода соединений характерным
является наличие двух типов белков.
Один из них - это трансмембранные
линкерные (связующие) белки, которые
участвуют или в собственно межклеточном
соединении или в соединении плазмолеммы
с компонентами внеклеточного матрикса
(базальная мембрана эпителиев, внеклеточные
структурные белки соединительной
ткани). Второй - внутриклеточные белки,
соединяющие или заякоревающие за
мембранные элементы такого контакта
цитоплазматические фибриллы цитоскелета.
аякоривающие
или сцепляющие
соединения
или контакты так называются из-за того,
что они соединяют не только плазматические
мембраны соседних клеток, но и связываются
с фибриллярными элементами цитоскелета
(рис.
149). Для этого рода соединений характерным
является наличие двух типов белков.
Один из них - это трансмембранные
линкерные (связующие) белки, которые
участвуют или в собственно межклеточном
соединении или в соединении плазмолеммы
с компонентами внеклеточного матрикса
(базальная мембрана эпителиев, внеклеточные
структурные белки соединительной
ткани). Второй - внутриклеточные белки,
соединяющие или заякоревающие за
мембранные элементы такого контакта
цитоплазматические фибриллы цитоскелета.
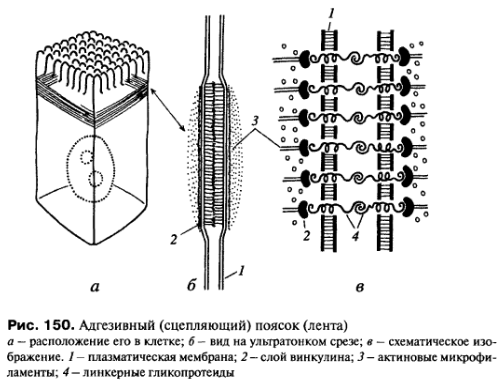 К
заякоревающим
соединениям относятся межклеточные
сцепляющие точечные контакты, сцепляющие
ленты, фокальные контакты или бляшки
сцепления - все эти контакты связываются
внутри клеток с актиновыми микрофиламентами.
К
заякоревающим
соединениям относятся межклеточные
сцепляющие точечные контакты, сцепляющие
ленты, фокальные контакты или бляшки
сцепления - все эти контакты связываются
внутри клеток с актиновыми микрофиламентами.
Другая группа заякоревающих межклеточных соединений - десмосомы и полудесмосомы - связываются с другими элементами цитоскелета, а именно с промежуточными филаментами.
Межклеточные точечные сцепляющие соединения обнаружены у многих неэпителиальных тканей, но более отчетливо описана структура специальных (адгезивных) лент в однослойных эпителиях (рис. 150). Это структура опоясывает весь периметр эпителиальной клетки, подобно тому как это происходит в случае плотного соединения. Чаще всего такой поясок или лента лежит ниже плотного соединения. В этом месте плазматические мембраны не сближены, а даже несколько раздвинуты на расстояние 25-30 нм, и между ними видна зона повышенной плотности.
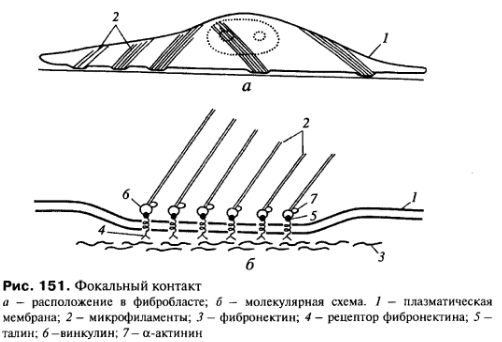 Фокальные
контакты
или бляшки
сцепления
встречаются у многих клеток и особенно
хорошо изучены у фибробластов. Они
построены по общему плану со сцепляющими
лентами, но выражены в виде небольших
участков - бляшек на плазмолемме. В этом
случае трансмембранные линкерные
белки-интегрины специфически связываются
с белками внеклеточного матрикса
(например с фибронектином) (рис.
151). Функциональное значение фокальных
контактов заключается как в закреплении
клетки на внеклеточных структурах, так
и создании механизма, позволяющего
клеткам перемещаться.
Фокальные
контакты
или бляшки
сцепления
встречаются у многих клеток и особенно
хорошо изучены у фибробластов. Они
построены по общему плану со сцепляющими
лентами, но выражены в виде небольших
участков - бляшек на плазмолемме. В этом
случае трансмембранные линкерные
белки-интегрины специфически связываются
с белками внеклеточного матрикса
(например с фибронектином) (рис.
151). Функциональное значение фокальных
контактов заключается как в закреплении
клетки на внеклеточных структурах, так
и создании механизма, позволяющего
клеткам перемещаться.
Десмосомы, структуры в виде бляшек или кнопок также соединяют клетки друг с другом (рис. 152, 153а). В межклеточном пространстве здесь также виден плотный слой, представленный взаимодействующими интегральными мембранными кадгеринами - десмоглеинами, которые сцепляют клетки друг с другом. С цитоплазматической стороны к плазмолемме прилежит слой белка-десмоплакина, с которым связаны промежуточные филаменты цитоскелета. Десмосомы встречаются чаще всего в эпителиях, в этом случае промежуточные филаменты содержат кератины.
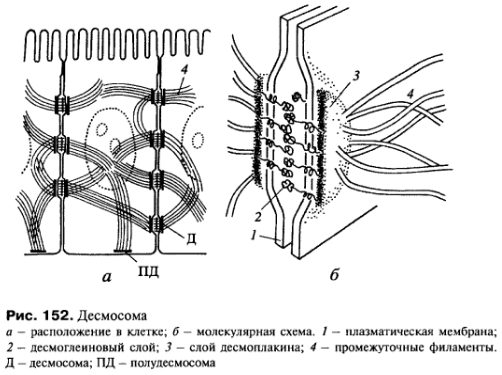 Полудесмосомы
- в принципе сходны по строению с
десмосомой, но представляют собой
соединение клеток с межклеточными
структурами. Так в эпителиях линкерные
гликопротеиды (интегрины) десмосомы
взаимодействуют с белками т.н. базальной
мембраны, куда входят коллаген, ламинин,
протеогликаны и др.
Полудесмосомы
- в принципе сходны по строению с
десмосомой, но представляют собой
соединение клеток с межклеточными
структурами. Так в эпителиях линкерные
гликопротеиды (интегрины) десмосомы
взаимодействуют с белками т.н. базальной
мембраны, куда входят коллаген, ламинин,
протеогликаны и др.
Функциональная роль десмосом и полудесмосом сугубо механическая - они сцепляют клетки друг с другом и с подлежащим внеклеточным матриксом прочно, что позволяет эпителиальным пластам выдерживать большие механические нагрузки.
 Щелевые
контакты
считаются коммуникационными соединениями
клеток; это структуры, которые участвуют
в прямой передаче химических веществ
из клетки в клетку, что может играть
большую физиологическую роль не только
при функционировании специализированных
клеток, но и обеспечивать межклеточные
взаимодействия при развитии организма,
при дифференцировке его клеток.
Щелевые
контакты
считаются коммуникационными соединениями
клеток; это структуры, которые участвуют
в прямой передаче химических веществ
из клетки в клетку, что может играть
большую физиологическую роль не только
при функционировании специализированных
клеток, но и обеспечивать межклеточные
взаимодействия при развитии организма,
при дифференцировке его клеток.
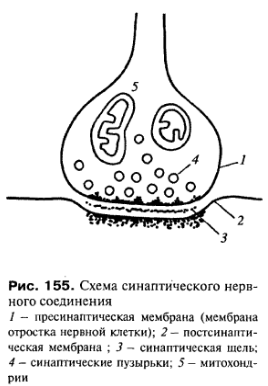
Синаптический контакт (синапсы). Этот тип контактов характерен для нервной ткани и встречается как между двумя нейронами, так и между нейроном и каким-либо иным элементом - рецептором или эффектором (например, нервно-мышечное окончание). Синапсы - участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому. Синапсы образуются на отростках нервных клеток - это терминальные участки дендритов и аксонов.
П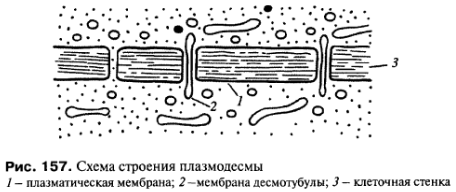 лазмодесмы.
Этот тип межклеточных связей встречается
у растений. Плазмодесмы представляют
собой тонкие трубчатые цитоплазматические
каналы, соединяющие две соседние клетки.
Диаметр этих каналов обычно составляет
20-40 нм. Ограничивающая эти каналы мембрана
непосредственно переходит в плазматические
мембраны соседствующих клеток. Плазмодесмы
проходят сквозь клеточную стенку,
разделяющую клетки (рис.
156, 157). Таким образом, у некоторых
растительных клеток плазмодесмы
соединяют гиалоплазму соседних клеток,
поэтому формально здесь нет полного
разграничения, отделения тела одной
клетки от другой, это скорее представляет
собой синцитий:
объединение
многих клеточных территорий с помощью
цитоплазматических мостиков.
лазмодесмы.
Этот тип межклеточных связей встречается
у растений. Плазмодесмы представляют
собой тонкие трубчатые цитоплазматические
каналы, соединяющие две соседние клетки.
Диаметр этих каналов обычно составляет
20-40 нм. Ограничивающая эти каналы мембрана
непосредственно переходит в плазматические
мембраны соседствующих клеток. Плазмодесмы
проходят сквозь клеточную стенку,
разделяющую клетки (рис.
156, 157). Таким образом, у некоторых
растительных клеток плазмодесмы
соединяют гиалоплазму соседних клеток,
поэтому формально здесь нет полного
разграничения, отделения тела одной
клетки от другой, это скорее представляет
собой синцитий:
объединение
многих клеточных территорий с помощью
цитоплазматических мостиков.