

Сизенцов А.Н. Общая вирусология с основами таксономии вирусов позвоночных
.pdfформировании капсида. Например, геномные белки пикорнавирусов и аденовиру-
сов. Геномный белок ковалентно связан с нуклеиновой кислотой и участвует в ее репликации.
Сложные белки представлены гликопротеинами (обозначают как gp) и липо-
протеинами. Наличие гликопротеина определяет присутствие в вирионе углевод-
ного компонента, который может быть представлен олигосахаридами маннозного типа, галактозой, N-ацетилглюкозамином или нейраминовой кислотой. Вирусные гликопротеины, как правило, экспонированы на наружной поверхности ВЧ и вы-
полняют три основные функции: обеспечивают связывание вириона с клеточным рецептором (функция прикрепительного белка), обладают фузионной активностью
(обеспечивают слияние мембран) и определяют антигенные свойства вирусов. В то же время, вирусные гликопротеины могут быть и неструктурными белками и, оста-
ваясь в интегральной форме в мембране шероховатого эндоплазматического рети-
кулюма (ШЭР), выполнять функции транслоказ, обеспечивая транспорт вирусных компонентов в его просвет.
Вирусные липопротеины представлены белками, ацилированными, как пра-
вило, миристиновой (C14) кислотой. Остатки жирных кислот, соединенные с моле-
кулой белка, выполняют функцию липофильного якоря.
Вирусные белки-ферменты могут входить в состав вирусной частицы или яв-
ляться неструктурными белками и появляться в клетке после экспрессии вирусного генома. Наиболее оснащенным ферментами является вирион вируса оспы, который имеет практически полный набор энзимов, необходимых для независимой внутри-
клеточной репликации вируса. В то же время, мелкие просто организованные изо-
метрические вирусы с позитивным РНК-геномом могут не иметь никаких фермен-
тов в составе вириона.
Функционально активные белки вирусов представлены, первую очередь,
ферментами нуклеинового обмена, обеспечивающими сложные механизмы репли-
кации/транскрипции вирусного генома; ферментами, осуществляющими посттранс-
ляционный процессинг и модификацию белков, и ферментами, участвующими в проникновении вирионов в клетку хозяина.
31
Первая группа ферментов наиболее многочисленна и включает как аналоги
клеточных ферментов, так и вирус-специфические ферменты. |
|
|
|
|||
ДНК-зависимая |
ДНК-полимераза – осуществляет |
синтез |
ДНК |
на |
матрице |
|
ДНК (вирус оспы). |
|
|
|
|
|
|
ДНК-зависимая |
РНК-полимераза – осуществляет |
синтез мРНК |
на |
матрице |
||
ДНК (вирус оспы). |
|
|
|
|
|
|
РНК-зависимая |
РНК-полимераза – осуществляет |
синтез |
РНК |
на |
матрице |
|
РНК. Выполняет |
функции транскриптазы и репликазы. Впервые обнаружена в |
|||||
1970 г. Балтимором у вируса везикулярного стоматита. Входит в |
состав вирионов |
|||||
или является NS-белком РНК-содержащих вирусов. |
|
|
|
|
||
Обратная |
транскриптаза или ревертаза или РНК-зависимая |
ДНК- |
||||
полимераза осуществляет синтез ДНК на матрице РНК. Впервые открыта в 1970 г. у
ретровирусов Темином и Мизутани.
Хеликаза – осуществляет расплетете двухнитевой структуры ДНК. Кроме это-
го хеликазы обладают нуклеотидтрифосфат-зависимой РНК-хеликазной активно-
стью, которая включает три процесса: связывание дезоксинуклеотидтрифосфата, его гидролиз и за счет этой энергии расплетение двухнитевой РНК.
мРНК-модифицирующие ферменты: поли-А-полимераза – аденилирует 3'-
конец РНК за счет энергии АТФ; Кэп-энзим и метилтрансферазный комплекс– ка-
тализирует образование на 5'-конце кэп-структуры.
АТФ-аза, ГТФ-аза – осуществляют гидролиз соответствующих энергетиче-
ских субстратов.
Рибонуклеаза Н – разрушает РНК, находящуюся в дуплексе с ДНК. Вторая группа вирусных ферментов – ферменты белкового обмена.
Здесь мы приведем лишь некоторые из них:
Протеиназы – ферменты, участвующие в посттрансляционном процессинге полипротеинов. Являются NS-белками РНК-содержащих вирусов;
Протеинкиназы – ферменты, фосфорилирующие структурные белки вирио-
нов. Обнаружены в составе вируса везикулярного стоматита, вируса бешенства,
альфавирусов и ретровирусов. Примерами ферментов, участвующих в проникнове32
нии вирусов в клетку, являются лизоцим бактериофагов и нейраминидаза вируса гриппа.
Липиды. Все оболочечные РНК-содержащие почкующиеся вирусы имеют ли-
пиды клеточного происхождения, входящие в состав суперкапсида (от 15 % до 30 %
от сухого веса). От 50 % до 60 % липидов представлены фосфолипидами, от 20 % до
30 % составляет холестерин.
У ДНК-геномных вирусов липиды содержат вирусы оспы, герпеса, гепатита B.
Это непочкующиеся вирусы. У вируса оспы липиды не образуют дифференциро-
ванной оболочки, которая формируется в цитоплазме в процессе морфогенеза -по ксвириона. Липиды вируса гепатита B образуются путем инвагинации мембран эн-
доплазматического ретикулюма (ЭПР). Липидсодержащая оболочка вируса герпеса формируется при прохождении внутреннего компонента вириона через ядерную мембрану. Следовательно, в состав вирусной оболочки герпесвирусов входят липи-
ды ядерной мембраны.
Нуклеиновые кислоты. Клетки всех живых организмов содержат два вида нуклеиновой кислоты – ДНК (двухнитевая ДНК клеточного генома) и РНК (мРНК,
тРНК, рРНК). В отличие от клеток, вирионы содержат только один вид нуклеиновой кислоты – ДНК или РНК. И та и другая являются хранителями наследственной ин-
формации и выполняют функции генома. Однако следует учитывать, что наличие одного вида нуклеиновой кислоты является характеристикой вириона, но не вируса.
В жизненном цикле вируса его геномная нуклеиновая кислота транскрибируется, то есть ДНК-содержащие вирусы образуют РНК. Ряд РНК-содержащих вирусов имеют в цикле репродукции стадию обратной транскрипции и синтезируют ДНК на матри-
це РНК. Примерно 20 % всех вирусов имеют ДНК-геном, 80 % – РНК-геном. Спо-
собность РНК хранить наследственную информацию– уникальное свойство виру-
сов. Размеры вирусных геномов (длина нуклеотидных последовательностей, выра-
женная в нуклеотидах) варьируют в широких пределах– от 1,7 тысяч нуклеотидов
(т.н.) у цирковируса свиней до 300 т.н. у фикоднавирусов архибактерий.
Кроме того, что геном вирусов может быть представлен или ДНК или РНК, он может находиться в разных видах– в виде двухнитевой (дн) или однонитевой (он)
33
формы, в виде линейной или кольцевой, в виде непрерывной или сегментированной формы.
Многообразие видов РНК геномов расширяется за счет существования после-
довательностей, отличающихся направлением связей сахаро-фосфатного остова.
Однонитевые РНК могут иметь позитивную полярность– (+) РНК, негатив-
ную полярность – (-) РНК или могут быть представлены обоюдозначащей цепью–
(+,-) РНК (амбисенс стратегия кодирования). В свою очередь, РНК позитивной по-
лярности могут иметь разную структурную организацию: могут, являясь матричной РНК, иметь на 5'-конце кэп (7-метилгуанозин, Сар), а на 3'-конце – поли-А (poly-A)
последовательность; могут не иметь кэпа или поли-А; могут иметь на 5'-конце ге-
номный белок; могут иметь на 3'-конце тРНК-подобную или шпильковую структу-
ру.
Виды геномов вирусов легли в основу их классификации. Однако следует учи-
тывать, что вид генома в настоящее время не является формальным таксоном и ис-
пользуется для удобства ориентации в многообразии вирусов.
Углеводы. Углеводный компонент вирусов находится в составе гликопротеи-
дов. Количество сахаров в составе гликопротеидов может быть достаточно боль-
шим, достигая от 10 % до 13 % от массы вириона. Химическая специфичность их полностью определяется клеточными ферментами, обеспечивающими перенос и присоединение соответствующих сахарных остатков. Обычными сахарными остат-
ками, обнаруживаемыми в вирусных белках, являются фруктоза, сахароза, манноза,
галактоза, нейраминовая кислота, глюкозамин. Таким образом, подобно липидам,
углеводный компонент определяется клеткой-хозяином, благодаря чему один и тот же вирус, выращенный в клетках разных видов, может значительно отличаться по составу сахаров в зависимости от специфичности клеточных гликозилтрансфераз.
Углеводный компонент гликопротеидов играет существенную роль в структу-
ре и функции белка. Он является каркасом для локальных участков гликопротеида,
обеспечивая сохранение конформации белковой молекулы, и обусловливает защиту молекулы от протеаз. Возможны и другие функции углеводов, пока достоверно не установленные.
34
Компоненты клетки-хозяина. В составе вирионов могут находиться компо-
ненты клетки-хозяина. К таким компонентам могут относиться белки, и даже целые клеточные структуры. Так, например, в составе ряда оболочечных вирусов может находиться белок цитоскелета актин, в составе паповавирусов содержатся клеточ-
ные гистоны. Ряд вирусов содержит клеточные ферменты, например, протеинкина-
зы. В составе аренавирусов обнаружены рибосомы.
Клеточные компоненты могут включаться в вирион случайно или закономер-
но, В некоторых случаях они играют существенную роль в репродукции вируса, как,
например, гистоны в репродукции паповавирусов.
3 Морфология, морфогенез, биофизические свойства и генетика
вирусов
3.1 Архитектура вирионов
Внеклеточная форма вируса – вирион, предназначенная для сохранения и пе-
реноса нуклеиновой кислоты вируса, характеризуется собственной архитектурой,
биохимическими и молекулярно-генетическими особенностями. Под архитектурой вирионов понимают ультратонкую структурную организацию этих надмолекуляр-
ных образований, различающихся размерами, формой и сложностью строения. Для описания архитектуры вирусных структур разработана номенклатура терминов:
Белковая субъединица – единая, уложенная определенным образом полипеп-
тидная цепь.
Структурная единица (структурный элемент) – белковый ансамбль более высокого порядка, образованный несколькими химически связанными идентичными или неидентичными субъединицами.
Морфологическая единица – группа выступов (кластер) на поверхности кап-
сида, видимая в электронном микроскопе. Часто наблюдаются кластеры, состоящие из пяти (пентамер) и шести (гексамер) выступов. Это явление получило название
35
пентамерно-гексамерной кластеризации. Если морфологическая единица соответст-
вует химически значимому образованию(сохраняет свою организацию в условиях мягкой дезинтеграции), то применяют термин капсомер.
Капсид – внешний белковый чехол или футляр, образующий замкнутую сферу вокруг геномной нуклеиновой кислоты.
Кор (core) – внутренняя белковая оболочка, непосредственно примыкающая к нуклеиновой кислоте.
Нуклеокапсид – комплекс белка с нуклеиновой кислотой, представляющий собой упакованную форму генома.
Суперкапсид или пеплос– оболочка вириона, образованная липидной мем-
браной клеточного происхождения и вирусными белками.
Матрикс – белковый компонент, локализованный между суперкапсидом и
капсидом.
Пепломеры и шипы – поверхностные выступы суперкапсида.
Как уже отмечалось, вирусы могут проходить через самые микроскопические поры, задерживающие бактерии, за что и были названы фильтрующимися агентами.
Свойство фильтруемости вирусов обусловлено размерами, исчисляемыми наномет-
рами (нм), что на несколько порядков меньше, чем размеры самых мелких микроор-
ганизмов. Размеры вирусных частиц, в свою очередь, колеблются в относительно широких пределах. Самые мелкие просто устроенные вирусы имеют диаметр чуть больше 20 нм (парвовирусы, пикорнавирусы, фаг Qβ), вирусы средних размеров – от
100 до 150 нм (аденовирусы, коронавирусы). Наиболее крупными признаны вирус-
ные частицы осповакцины, размеры которых достигают 170 ´ 450 нм. Длина ните-
видных вирусов растений может составлять 2000 нм.
Представители царства Vira характеризуются разнообразием форм. По своей структуре вирусные частицы могут быть простыми образованиями, могут пред-
ставлять собой достаточно сложные ансамбли, включающие несколько структурных элементов.
Существует два типа вирусных частиц(ВЧ), принципиально отличающихся друг от друга:
36
1)ВЧ, лишенные оболочки (безоболочечные или непокрытые вирионы);
2)ВЧ, имеющие оболочку (оболочечные или покрытые вирионы).
Строение вирионов, лишенных оболочки. Выделено три морфологических типа вирионов, лишенных оболочки: палочковидные (нитевидные), изометрические и булавовидные (рис. 2). Существование первых двух типов непокрытых вирионов определяется способом укладки нуклеиновой кислоты и ее взаимодействием с бел-
ками.
1 Белковые субъединицы связываются с нуклеиновой кислотой, располагаясь вдоль нее периодическим образом так, что она сворачивается в спираль и образует структуру под названием нуклеокапсид. Такой способ регулярного, периодического взаимодействия белка и нуклеиновой кислоты определяет образование палочковид-
ных и нитевидных вирусных частиц.
2 Нуклеиновая кислота не связана с белковым чехлом(возможные некова-
лентные связи очень подвижны). Такой принцип взаимодействия определяет обра-
зование изометрических (сферических) вирусных частиц. Белковые оболочки виру-
сов, не связанные с нуклеиновой кислотой, называют капсидом.
3 Булавовидные вирионы обладают дифференцированной структурной орга-
низацией и состоят из ряда дискретных структур. Основными структурными эле-
ментами вириона являются изометрическая головка и хвостовой отросток. В зави-
симости от вируса в структуре вириона также могут присутствовать муфта, шейка,
воротничок, хвостовой стержень, хвостовой чехол, базальная пластинка и фибрил-
лы. Наиболее сложную дифференцированную структурную организацию имеют бактериофаги T-четной серии, вирион которых состоит из всех перечисленных структурных элементов.
Вирионам или их компонентам могут быть присущи два основных типа сим-
метрии (свойство тел повторять свои части) – спиральный и икосаэдрический. В том случае, если компоненты вириона обладают разной симметрией, то говорят о ком-
бинированном типе симметрии ВЧ (рисунок 2 а).
Спиральная укладка макромолекул описывается следующими параметрами:
числом субъединиц на виток спирали(u, число необязательно целое); расстоянием
37
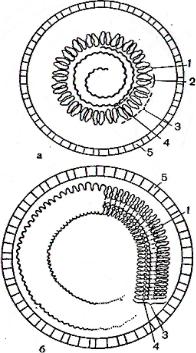
между субъединицами вдоль оси спирали(p); шагом спирали (P); P = pu. Классиче-
ским примером вируса со спиральным типом симметрии является вирус табачной мозаики (ВТМ). Нуклеокапсид этого палочковидного вируса размером18 ´ 300 нм состоит из 2130 идентичных субъединиц, на виток спирали приходится 16 1/3 субъ-
единиц, шаг спирали составляет 2,3 нм (рисунок 2 б).
1-структурная единица капсида; 2-морфологическая единица капсида (капсо-
мер); 3-капсид; 4-нуклеиновая кислота; 5-суперкапсид.
Рисунок 2 – Кубическая (а) и спиральная (б) симметрия капсидов сложно уст-
роенных вирусов (схема)
Икосаэдрическая симметрия – самая эффективная для конструирования замкнутого чехла из отдельных субъединиц. При рассмотрении элементов икосаэд-
рической симметрии следует различать понятия симметрия и форма. Симметрия в данном случае – это набор поворотов, которые переводят объект сам в себя, форма – это лишь общий вид кубической поверхности объекта(тетраэдр, октаэдр, додекаэдр и т.д.). Многие объекты, имея икосаэдрическую симметрию, не имеют икосаэдриче-
ской формы. Икосаэдр – это геометрическая фигура, имеющая 12 вершин, 20 граней, 20 ребер.
38
Наименьшее число структурных элементов, способных образовать икосаэдр,
равно 60, однако капсиды сложноустроенных вирусов могут быть образованы60 n
структурными элементами. Для описания икосаэдрической упаковки структурных элементов в капсиде введено так называемоетриангуляционное число (T). Это число, равное частному от деления числа субъединиц на60. Так, у вируса некроза табака и фага φX174 T = 1 (60 субъединиц), многие вирусы растений имеютT = 3 (180 субъединиц), вирус Синдбис имеет T = 4 (240 субъединиц), ротавирус имеет
T = 13 (780 субъединиц).
Многие крупные икосаэдрические вирусы для получения плотной упаковки капсида формируют субтриангуляции на основе структур меньших размеров, что предполагает наличие разных типов субъединиц на вершинах икосаэдра и наруше-
ние локальной симметрии в местах их контактов. В этом случае наблюдается расхо-
ждение между реально существующей симметрией ВЧ и видом структуры с соот-
ветствующим числом Т. Наиболее простую конструкцию капсида, построенного по такому принципу, имеют паповавирусы. Их капсид образован 72 морфологическими единицами, каждая построена из трех белковых субъединиц, организованных в пен-
тамеры, а ВЧ имеет вид структуры с Т = 7.
Более сложная структура вириона наблюдается у аденовируса, капсид которо-
го организован по принципу ансамблей, обладает строгой икосаэдрической симмет-
рией и имеет вид структуры с Т= 25. На вершинах икосаэдра находятся кластеры – пентоны, содержащие в основании так называемые фибры – стержень с утолщением на конце. Остальная структура капсида построена из гексонов. Гексоны и пентоны – это простейшие подструктуры капсида аденовирусов. Всего в состав аденовириона входит 12 оснований пентонов и 240 гексонов. При диссоциации в мягких условиях образуются надструктуры (капсомеры), состоящие из 9-ти гексонов.
Еще более сложноустроенные вирионы, на пример частицы бактериофагов T-
чётной серии, обладают комбинированным типом симметрии. Так, головка бакте-
риофага T4 имеет икосаэдрический тип симметрии, а сокращенный чехол хвостово-
го отростка обладает спиральным типом симметрии. В целом вирион фага T4 обла-
дает комбинированным типом симметрии.
39
Строение вирионов с оболочкой. Другой тип вирусных частиц – это покры-
тые или оболочечные вирионы. Оболочечные вирионы, также как и непокрытые,
могут быть палочковидными, нитевидными и изометрическими разной формы– от четко очерченных кирпичеобразных вирионов вируса оспы до плейоморфных час-
тиц вирусов герпеса и коронавирусов, имеющих различные размеры и форму.
Оболочка вириона (пеплос, суперкапсид) состоит из липидсодержащей мем-
браны клеточного происхождения (цитоплазматической мембраны, мембраны эндо-
плазматического ретикулюма или аппарата Гольджи, ядерной мембраны) и вирус-
ных гликопротеинов, встроенных в мембрану. Оболочку вирионы приобретают в процессе почкования через ту или иную мембрану.
Вирусные гликопротеины, находящиеся в мембране, как правило, формируют поверхностные выступы, называемые шипами и пепломерами. Эти поверхностные выступы характеризуются разной степенью упорядоченности и могут быть пред-
ставлены одним белком (вирус кори) или двумя разными белками(вирусы гриппа,
ретровирусы), могут быть образованы мономерами белка или его димерами и три-
мерами.
Таким образом, структурная организация вириона описывается двумя харак-
теристиками – наличием/отсутствием оболочки и типом симметрии капсида. Оболо-
чечные вирионы могут обладать икосаэдрической, спиральной и комбинированной симметрией капсида, также как и безоболочечные.
3.2 Морфогенез вирусов
При внутриклеточной репродукции вирусов формируютсяструктуры, отсут-
ствующие в незаряженных вирусом клетках. Эти образования – места синтеза и сборки субвирусных структур (компонентов дочерних вирионов) получили разные наименования – клеточные матриксы «фабрики», виропласты, включения. Эти структуры являются продуктами кооперативных процессов клетки и вируса, где главенствующая роль принадлежит клетке.
Морфологически матриксы выглядят по-разному у разных вирусов. Обычно
40