

головного мозга у других амниот нет (Савельев, 2001).
§ 46. Обзор классификации млекопитающих
Класс современных млекопитающих разделяют на два подкласса: первозвери (Prototheria) и настоящие звери (Theria). К первозверям относят однопроходных млекопитающих (Monotremata), которые могут откладывать яйца, но выкармливают детёнышей молоком и имеют своеобразный волосяной покров. Настоящие звери объединяют две обширные группы НИЗШИХ зверей (Metatheria) и высших зверей (Eutheria). Низшие звери представляют собой сумчатых млекопитающих американского и австралийского происхождения, а к высшим зверям относят всех остальных млекопитающих, объединённых в 18 обширных и часто искусственных отрядов (см. рис III-17). Основным отличием низших зверей от высших является морфофункциональный тип организации внутриматочного питания эмбрионов и плодов. Низшие звери рождают очень небольших детёнышей, которые скорее напоминают эмбрионов. Их дальнейшее развитие проходит в специальной сумке, куда открываются молочные железы. Высшие млекопитающие рождают, как правило, крупных и более сформированных детёнышей. Развитие эмбрионов и плодов высших млекопитающих обеспечивается
316
специальным приспособлением — плацентой. Она имеет смешанное строение и состоит из материнской децидуальной выстилки матки и специализированного участка оболочек плода. У высших млекопитающих плаценту называют аллантоисной и инвазивной. В состав её стенки входят (снаружи внутрь) эктодермальный трофобласт, не образующая сосудов соматическая мезодерма, образующая сосуды мезодерма и энтодермальный слой клеток. Вне плаценты у высших млекопитающих оболочку плода составляют эктодермальный трофобласт и мезодерма, не образующая сосудов. Через внеплацентарные оболочки обычно могут проникать электролиты и вода. Основной обмен осуществляется только через плаценту. Между разветвлениями капилляров плодной части плаценты выбрасывается материнская кровь. Затем происходит обмен кислорода и продуктов метаболизма. Обогащённая кровь плода оттекает к зародышу, а материнская возвращается в родительский организм.
У низших зверей встречается настоящая инвазивная плацента (бандикут), но в большинстве случаев она неинвазивная, не образует капиллярной сети со стороны плода и играет скромную роль в обеспечении плода кислородом и метаболитами. У низших зверей плацента желточная и бывает трёх типов. Если она состоит из трофобласта, мезодермы двух типов и стенки желточного мешка, то может образовывать капиллярную сеть или обходиться без неё. Если желточная плацента состоит только из трофобласта и стенки желточного мешка, она не имеет сосудистой системы. Обмен кислорода и метаболитов происходит через обширную стенку желточной плаценты, которая может сочетаться с инвазивной (бандикут) и неинвазивной аллантоисной плацентой (коала). Однако в большинстве случаев аллантоисная плацента вообще отсутствует (опоссум). Желточная плацента позволяет путём диффузии выращивать относительно небольшие зародыши внутри матки. Доращивание происходит в сумке матери. У высших млекопитающих аллантоисная плацента снимает ограничение на размер зародыша. Он зависит только от морфологии или размера тазовых костей самки.
Среди малочисленной группы однопроходных млекопитающих наиболее интересен утконос (Ornithorhynchus anatinus). Он имеет кожистый клюв, перепончатые лапы и температуру тела около 25°С. Самка утконоса откладывает в норах небольшие кожистые яйца диаметром около 15 мм, которые нужно согревать в течение 7-11 дней. Вылупившиеся детёныши питаются молоком, стекающим из простых молочных желёз по волоскам на брюхе матери. Ехидны (Tachyglossidae) также откладывают яйца, которые вынашивают в примитивной сумке. Их детёныши питаются молоком, стекающим по волоскам из примитивных молочных желёз, как и у утконоса. Ехидны имеют нестабильную температуру тела
317

Рис. III-17. Разнообразие современных млекопитающих.
Современные млекопитающие освоили практически все среды обитания и представляют бесконечное разнообразие форм биологических адаптаций, в основе которых лежит развитие сложного ассоциативного мозга.
318
около 30°С. Кратковременная спячка позволяет ехиднам и утконосам переживать неблагоприятные периоды. При всех особенностях морфологического строения и древней биологии размножения однопроходные обладают весьма совершенной нервной системой (Griffiths, 1978).
Головной мозг утконоса весит около 16 г, парные полушария переднего мозга полностью скрывают средний мозг и часть мозжечка. Полушария переднего мозга не имеют извилин (лиссэнцефальны), но мозжечок развит, как и у высших млекопитающих. Утконос способен к электрорецепции и обладает рядом сложных соматических рецепторов, не встречающихся у других млекопитающих. Ответы электрорецепторов клюва утконоса были обнаружены при изменении напряжённости электрического поля. Утконос воспринимает не только напряжённость, но и частотные характеристики электромагнитного поля. Электромагнитная чувствительность и адаптированная сенсомоторная система позволяют утконосу находить пищу в мутной воде. Мозг утконоса представляет собой настолько глубоко специализированную конструкцию, что не может рассматриваться в качестве примера архаичной или примитивной организации.
У ехидны мозг не только крупного размера, но и с развитыми бороздами и извилинами на поверхности неокортекса (рис. III-21, а). Иначе говоря, у яйцекладущих млекопитающих развитый, совершенный мозг со сложноорганизованным неокортексом. Судя по его строению, эта группа является скромным биологическим следом древней и обширной ветви яйцекладущих млекопитающих. Мозг ехидн сходен по строению с мозгом крупных кенгуру или широконосых обезьян, но не с мозгом насекомоядных или грызунов. Однопроходные настолько специализированны, что архаичных следов нервной системы у них намного меньше, чем у Eutheria. Единственным принципиальным отличием мозга однопроходных и сумчатых является отсутствие мозолистого тела, которое является крупной межполушарной комиссурой (см. рис. III-21, г, д). Поэтому не приходится надеяться, что морфологическое строение современных однопроходных сможет оказать действенную помощь в восстановлении истории происхождения нервной системы млекопитающих.
Среди первозверей обширную группу составляют американские и австралийские сумчатые (Marsupialia). Сложная биология их размножения связана с ранним рождением детёнышей и их длительным доращиванием в специальной сумке матери. Детёныши присасываются в сумке к молочным железам и быстро растут. Размеры и формы современных сумчатых очень различны. Наиболее крупные экземпляры серого кенгуру достигают длины 3 м и массы 80 кг, а сумчатые мыши
319
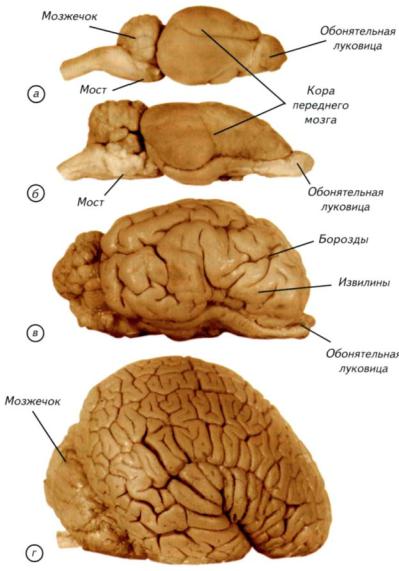
Рис. III-18. Головной и спинной мозг современных млекопитающих.
а — вскрытый головной мозг обезьяны с двумя утолщениями спинного мозга со спинной стороны; б — крот (Talpa europaea) с отпрепарированным мозгом; в — макака-резус (Масаса mulatta) с видом на мозг с латеральной поверхности; г — сагиттальный разрез головы свиньи (Sus scrofa).
320
могут быть меньше 10 см и массой 50-90 г. Среди сумчатых большую группу составляют передвигающиеся на задних конечностях кенгуру (Macropodidae). У крупных кенгуру (Macropus, Megaleia) прыжки достигают 10 м, а скорость движения около 40 км/ч. Прыгая на задних конечностях, кенгуру используют хвост как балансир, что требует хорошей координации движений и контроля за центром тяжести тела. Этим задачам соответствуют развитый мозжечок и неокортекс. У большинства мелких сумчатых лиссэнцефальный (лишённый борозд и извилин) передний мозг. Хищные сумчатые (Dasyuridae), австралийские бандикуты (Peramelidae) и сумчатые медведи (Phascolarctidae) имеют развитую гирификацию или одиночные извилины.
Насекомоядные (Insectivora) представляют собой наиболее примитивную в неврологическом отношении группу высших зверей. Строение их головного мозга сохранило множество архаических черт. Он лиссэнцефален, имеет небольшой неокортекс и развитые обонятельные центры (см. рис. III-18; III-19). Биология размножения насекомоядных традиционна и достаточно прогрессивна для млекопитающих. Она включает не только выкармливание детёнышей молоком, но и довольно продолжительную заботу о потомстве. По-видимому, насекомоядные специализировались на заре эволюции млекопитающих, что позволило им сохранить некоторые признаки наиболее древних черт морфологического строения. Надо подчеркнуть, что ежи (Erinaceidae), гимнуровые (Echinosoricidae), тенрековые (Tenrecidae) и кротовые (Talpidae) намного примитивнее в неврологическом отношении, чем современные сумчатые. Это делает их
привлекательными объектами для эволюционной неврологии. Представители этой группы наиболее близки к архетипу строения мозга древнейших млекопитающих.
Среди всего разнообразия млекопитающих, кроме насекомоядных, следует выделить грызунов и зайцеобразных, которые могут пролить свет на историю прогрессивного развития сложной архитектуры переднего мозга (см. рис. III-18; III-19; III-21). Вместе с тем ни в одной группе животных до млекопитающих мозг не достигал таких гигантских размеров, как у приматов, хоботных и китообразных (см. рис. III-19, г; III-22, б). Ни в одной группе позвоночных не было столь избыточного неврологического субстрата для развития памяти, индивидуального научения и ассоциативного мышления. В головном мозге высших млекопитающих сложились совершенно новые принципы иерархической обработки информации в распределённых системах, которые стали основой для мышления. Мозг высших млекопитающих отличается межполушарной системой связей — мозолистым телом. Это образование состоит из отростков нейронов,
321
Рис. III-19. Головной мозг современных млекопитающих.
а — ёж (Erinaceus europaeus); б — кролик (Oryctolagus cuniculus); в — домашняя корова; г — афалина
(Tursiops truncatus). Головной мозг кролика и ежа лиссэнцефальный, полушария переднего мозга коровы и дельфина содержат большое количество борозд и извилин.
322
расположенных в одном полушарии и передающих информацию в другое. Обычно это нейроны, принадлежащие к одной сенсорной или моторной системе. С их помощью происходит сравнение сигналов, приходящих в правое и левое полушарие. Учитывая размер мозолистого тела, можно сказать, что это одно из принципиальных эволюционных приобретений высших зверей.
Сложность поведения млекопитающих не нуждается в подробном иллюстрировании. Тем не менее надо ясно представлять себе реальную ситуацию с интеллектуальными возможностями большей части представителей этой группы. Говоря о поведении млекопитающих, обычно подразумевают приматов, китообразных, наземных хищников или адаптированных домашних животных. Интеллектуальные возможности этих животных, безусловно, намного выше таковых у беспозвоночных, анамний и рептилий. Большинство из них могут прогнозировать результат своих действий, выражать эмоции, играть, устанавливать иерархические