

Поведение рептилий построено на врождённых реакциях, которые заметно индивидуализируются в процессе жизни. Этому способствует продолжительность приобретения индивидуального опыта. Крупные черепахи живут до 150-250 лет, змеи и ящерицы — 7-25 лет, а крокодилы — до 70-80 лет. У многих рептилий очень большую роль играет обучение. Они легко приручаются и быстро осваивают лабиринт или зрительные условные сигналы. Черепахи, принадлежащие к древнейшим анапсидам, могут вторично узнать фигуру, нарисованную сплошной линией, даже если она очерчена пунктиром или повёрнута на небольшой угол. Ящерицы и змеи способны к некоторому прогнозированию развития событий. Это позволяет им преследовать добычу уже после того, как она скрывается из виду. Однако водяные ужи, обученные правильно проходить Т- образный лабиринт, после линьки всё забывают. Аналогично ведёт себя шипохвост, который, привыкнув к новой территории, забывает её после кратковременного охлаждения. Крокодилы проявляют довольно изощрённую заботу о потомстве. Они готовят кучи из земли, листвы и веток, куда откладывают яйца. Пока зародыши развиваются, крокодилы косвенными способами контролируют температуру и охраняют кладку, а затем выкапывают вылупляющихся потомков.
У рептилий известна иерархическая организация компактных групп. Доминирующей игуане уступают место не только на своей территории, но и на соседних, когда она направляется на охоту в общие нейтральные зоны. Иерархия у гигантских черепах Галапагосских островов выражается в строго установленном порядке следования друг за другом во время миграций. Групповая организация рептилий является одной из сложных форм общественного поведения, которая была достигнута у холоднокровных животных. Перечисленные особенности современных животных только косвенно отражают зоопсихологические достижения вымерших рептилий. Однако всё разнообразие их поведения базируется на головном мозге общего архетипа, который заслуживает отдельного рассмотрения.
§ 37. Ассоциативный центр мозга рептилий
Рассмотрев общий план строения нервной системы, следует отдельно остановиться на новых принципах организации и работы мозга, впервые реализованных у рептилий. Нервная система архаичных амниот стала логическим развитием строения удачной амфибийной конструкции. Однако мозг амфибий практически выполнял функцию сложного рефлекторного аппарата, а его интеллектуальные возможности остались невостребованными. Эволюция амфибий решалась мышцами, зубами, линейными размерами и масштабами
271
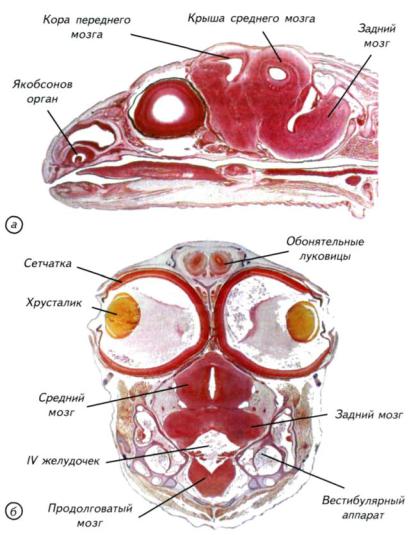
Рис. III-8. Сагиттальный (а) и горизонтальный (б) гистологические срезы через голову ящерицы (Lacerta agilis) в конце эмбрионального развития.
Относительный размер мозга в несколько раз больше, чем у амфибий. К моменту вылупления рептилии обладают эффективным вестибулярным аппаратом, зрением, слухом и обонятельной системой.
272
размножения. Шло элементарное освоение пищевых ресурсов, где для развития сложного поведения не было ни места, ни биологической необходимости. Со следами этого периода эволюции позвоночных мы сталкиваемся, пытаясь выработать условные рефлексы у различных представителей современных амфибий. Крайне низкая обучаемость и отсутствие долговременной памяти для накопления индивидуального опыта показывают, что сложных поведенческих задач перед древними амфибиями никогда в их эволюции не стояло.
Особенности развития органов чувств и признаки сложного поведения рептилий базируются на особенностях структурной организации головного мозга. Мозг рептилий отличается от мозга амфибий как в количественном, так и в качественном отношении. До появления амниот стратегии поведения или реакция на конкретный раздражитель выбирались по принципу доминантности (см. рис. III-6, е). Этот принцип состоит в том, что выраженного крупного ассоциативного центра мозга у многих первичноводных позвоночных или амфибий нет (см. рис. III-6, e). Выбор формы поведения происходит на основании сравнения активностей примерно равноценных отделов мозга, обслуживающих различные органы чувств. Решающее значение играет уровень возбуждения мозговых аналитических центров одного из анализаторов. Представительство органа чувств, достигшее в мозге наибольшего возбуждения, и становится основной областью для принятия решения. После выбора одной из инстинктивных реакций происходит её поведенческая реализация. Этот процесс осуществляется под контролем того же простого сравнения доминантностей. Если в процессе осуществления реакции возникает новое раздражение, которое изменяет соотношение возбуждений органов чувств, то поведенческая реализация инстинктивного процесса останавливается. Каждая конкретная ситуация отличается от предыдущей, но задействуется тот же набор органов чувств. Если наибольшее возбуждение достигается в той же сенсорной системе, то поведение сохраняется, а если в другой, то изменяется. Поскольку абсолютно идентичные условия в естественной жизни практически не встречаются, поведение даже самых примитивных анамний будет бесконечно разнообразно. Следовательно, поведение каждой особи будет индивидуальным с довольно высокой динамической адаптивностью.
Первые признаки ассоциативного центра ПОЯВИЛИСЬ ещё в мозге амфибий. У них таким центром мог стать средний или промежуточный мозг. Для этого были все основания. В промежуточном мозге находятся нейроэндокринные центры, контролирующие половое поведение, миграции и энергетический баланс организма анамний. Через
273
активизацию центров промежуточного мозга запускаются инстинктивные программы поведения, которые контролируют работу других отделов мозга. Казалось бы, промежуточный мозг мог стать аналитическим центром поведения анамний, а затем и амниот. Однако в этом случае система реализации поведенческих реакций состояла бы не только в работе нервной системы. Каждый раз любое поведенческое событие приводило бы к стимуляции работы нейроэндокринных центров. Гормональная регуляция поведения реализуется долго, а неврологическая — быстро. При быстрой смене форм поведения наступал бы конфликт между инертными гормональными и динамичными нейральными программами поведения. У насекомых этот конфликт был решён в пользу нейрогормональных центров и чисто инстинктивного поведения.
С первичноводными позвоночными и амфибиями сложилась довольно нестабильная ситуация. С одной стороны, роль гормонально-инстинктивной регуляции поведения у амфибий очень велика и явно доминирует при выборе стратегий поведения. С другой стороны, нейроморфологический субстрат развит вполне достаточно для небольшой индивидуализации поведения при реализации этих стратегий. Возникла оригинальная система гормонально-доминантного выбора форм поведения из стандартного инстинктивного набора. У амфибий стратегия поведения определяется нейрогормональным состоянием особи. При реализации выбранной формы поведения она адаптируется к конкретным условиям при помощи сравнения доминантностей, которое было описано выше. В такой схеме контроля поведения анамний не остаётся места для ассоциативного центра. Он мог бы понадобиться только тогда, когда возникла бы потребность в быстрой адаптивной индивидуализации поведения. Такая ситуация может сложиться только в том случае, когда последовательная реализация инстинктивных форм поведения будет прямо зависеть от постоянно меняющейся ситуации.
В таких нестабильных условиях окружающей среды оказались архаичные рептилии. По-видимому, требования к быстрой индивидуализации поведения и памяти возросли, а реализация гормональнодоминантного принципа выбора из стандартного инстинктивного набора форм поведения стала неэффективной. Возник совершенно новый тип принятия решений, который сохранился в мозге современных рептилий, принадлежащих к отдалённым систематическим группам. Их всех объединяет одно принципиально новое качество мозга — выраженный ассоциативный центр (см. рис III-7).
274
Основной ассоциативный центр рептилий сформировался в крыше среднего мозга (см. рис. III-5, в; III-6; III-7, б). Он возник на базе нескольких органов чувств, которые имели представительство в этом отделе головного мозга. Основную часть крыши среднего мозга занимает представительство зрительной системы. Зрительные нервы после прохождения хиазмы перекрещиваются и поднимаются к крыше среднего мозга. Аксоны ганглиозных клеток сетчатки оканчиваются на нейронах крыши среднего мозга, которые организованы в стратифицированные структуры (см. рис. III-5, в; III-7, б). Существует чёткая топографическая связь между определённым участком сетчатки и зоной крыши среднего мозга. При этом соблюдаются форма изображения и взаимное расположение его элементов. Довольно долго переднюю часть крыши среднего мозга считали исключительно мозговым центром зрительного анализатора. Однако функционально-морфологические исследования показали, что это далеко не так.
Наряду с представительством зрительной системы в крышу среднего мозга приходят информация о соматической (кожной) чувствительности, двигательном анализаторе, вестибулярные и слуховые сигналы (см. рис. III-6, д). Слуховой анализатор у рептилий значительно увеличивает своё представительство в этом центре. В результате у многих рептилий в задней части крыши среднего мозга появляются малозаметные парные выпячивания — задние или нижние бугорки. Крыша среднего мозга становится не однородным анатомическим образованием, как у анамний, а четверохолмием. В ней сосредоточивается представительство практически всех основных дистантных и контактных анализаторов. Даже обонятельная система имеет своё представительство в крыше среднего мозга. За исключением обонятельной системы, практически все сенсорные проекции в крышу среднего мозга рептилий носят топологический характер. Это означает, что информация от каждого конкретного участка тела представлена в строго определённом участке крыши среднего мозга. Сохраняется принцип карты тела, которая точечно переносится в мозг.
Таким образом, в крыше среднего мозга рептилий сосредоточена разнообразная информация о состоянии собственного организма и окружающего мира, которая объединена по топологическому принципу. Рассмотрим, что происходит в крыше среднего мозга, если передняя правая конечность рептилии просто стоит на необычной поверхности. При оценке такой ситуации в крыше среднего мозга осуществляется сравнительный анализ соматической, сенсомоторной, слуховой и зрительной информации. Это легко сделать, поскольку все сигналы концентрируются в одном центре, а зачастую и друг над
275
другом, как в слоистом пироге. Средний мозг проводит комплексный анализ многих факторов одного явления, что позволяет выбрать наиболее адекватную реакцию. Для этого идеально подходит стратифицированная структура крыши среднего мозга.
Предельно упрощая реальную ситуацию, можно сказать, что в крыше среднего мозга представительство различных сенсорных систем расположено на условных «этажах», организованных в горизонтальной плоскости. Каждый этаж занимает своеобразная карта. Она может быть сиюминутной информационной картой рецепторных сигналов от поверхности тела, изображением на сетчатке или акустическим полем. Все эти карты ориентированы на своих «этажах» так, что отражают примерно одно и тоже направление в пространстве. Слуховой сигнал от передней правой конечности лежит под её зрительной картой и над соматическим сигналом от кожи стопы. Специализированные «этажи» соотнесены между собой при помощи