

24
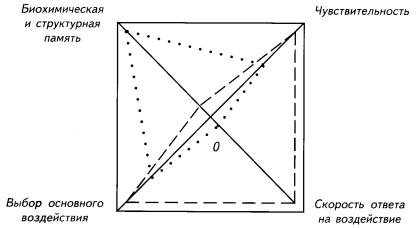
Рис. I-3. Донервная интеграция ответов на внешние воздействия у растений (точки) и свободноживущих простейших (пунктир).
Различия сводятся к скорости ответа на воздействие, которая зависит от размера тела. Скорость реакций высокая у простейших, но у растений есть структурная «память», сохраняющая результаты предыдущих воздействий. Площади образовавшихся полей могут рассматриваться как критерии оценки направлений адаптивности донервных организмов.
построена на биохимических внутриклеточных процессах. Это делает невозможным использование механизмов индивидуального научения. Следовательно, основным принципом донервного взаимодействия с внешней средой является общая клеточная чувствительность, которая имеет избирательный характер. Общая клеточная чувствительность построена на трёх различных рецепторных механизмах: механочувствительности, хемочувствительности и электромагнитной рецепции. Однако эти условные «сенсорные варианты восприятия мира» далеко не равноценны. Надо подчеркнуть, что иллюзию осознанности поведения одноклеточных формирует скорость ответа на неспецифические влияния. Основные различия построены на скорости реакции при внешнем воздействии на организм. Если организм по сравнению с нами маленький и реагирует быстро, то нам кажется, что реакции осознанны и сходны с поведением животных, имеющих нервную систему. Отсутствие оперативной индивидуальной памяти компенсируется генетически детерминированными реакциями или направленным морфогенезом. Если бы растения и простейшие располагали индивидуальной памятью, они были бы самыми опасными существами на планете. К нашему счастью, для этого нужна нервная система.
25
Зачем нужна нервная система?
Нервная система нужна не всем. Она не нужна тем, кто будет тысячелетиями неподвижен. Их «быстрый ответ» на внешние воздействия растягивается на десятки лет, им не надо ни быстрой реакции, ни мгновенной перестройки организма. Растения решили свои проблемы при помощи автотрофности, размера и времени жизни. Существует и другой вариант жизни без нервной системы. Можно поселиться в чудесном месте, где много пищи, организм защищен и согрет. Жизнь солитёра вполне соответствует этим требованиям. Солитёр и растения не обладают нервной системой. У растений нервной системы никогда не было, а у солитёра она полностью редуцирована. В обоих случаях её функции выполняют отдельные клетки, обладающие химической, электромагнитной и механической чувствительностью. Действительно, в оптимальных условиях питания и размножения нервная система вообще не нужна. Однако завидная для многих приматов судьба паразитических червей скорее исключение, чем правило в животном мире. Для большинства организмов окружающий мир слишком нестабилен и требует постоянного приспособления к нему. Адаптация к изменяющимся условиям должна быть генерализованной и охватывать все структуры организма. Таким органом быстрого и интегрированного реагирования стала нервная система.
Нервная система нужна тем, кто быстро двигается, активно вступает в контакт с разнообразными условиями внешнего мира и вынужден постоянно приспосабливаться. Быстротечный конформизм — причина формирования нервной системы. Действительно, нервная система малоподвижных организмов окажется невероятно упрощённой по сравнению с нервной системой активных животных. Актинии, ас-цидии, малоподвижные моллюски с крупными раковинами, коралловые полипы и многие другие животные имеют несложную нервную систему (рис. 1-4, а). У животных, прикреплённых к конкретному месту и занимающихся фильтрацией или захватом проплывающей пищи, очень простые задачи. Первая — рецепция пищевого объекта, вторая — его захват и переваривание. Для этого достаточно простых контактных рецепторов и органов удержания пищи, что, собственно, мы и видим у свободноживущих гидр и полипов. Их диффузная нервная система имеет небольшое окологлоточное нервное кольцо, которое и интегрирует несложные рефлексы.
Тем не менее эти простые реакции протекают на несколько порядков быстрее, чем морфогенетические перестройки тела у растений