

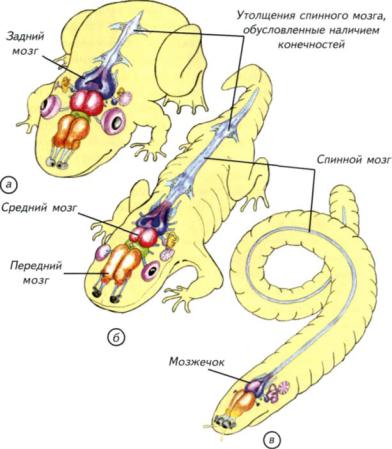
Рис. II-26. Общий план строения нервной системы амфибий, вписанный в контур тела представителя каждого из отрядов.
а — нервная система бесхвостых амфибий (Anura); б — нервная система хвостатых амфибий (Urodela); в
— нервная система безногих амфибий (Apoda). Передний мозг и мозжечок более всего развиты у безногих амфибий в связи с почвенным образом жизни. Средний мозг наиболее совершенен у бесхвостых амфибий, а водные хвостатые амфибии сохранили признаки иннервации большинства органов боковой линии рыб. У многих безногих амфибий отсутствуют утолщения спинного мозга, маркирующие конечности, что свидетельствует об их отсутствии у предковых форм.
224
§ 30. Особенности строения нервной системы амфибий
Нервная система амфибий имеет много сходных черт с рыбами, но обладает и рядом особенностей. Хвостатые и бесхвостые амфибии приобрели конечности, что повлекло за собой изменение организации спинного мозга. Спинной мозг лягушки короткий и оканчивается тонкой конечной ниточкой. В соответствии с числом позвонков у лягушки существует только 10 пар спинных нервов (см. рис. II-27); 2-я и 3-я пары сливаются и образуют плечевое сплетение, проводящее нервы в передние конечности, 7-я, 8-я, 9-я и 10-я пары соединяются для образования крестцово-поясничного сплетения, которое иннервирует задние конечности. Такая специализация нервов спинного мозга впервые встречается у амфибий и отсутствует у рыб. Есть ещё одно важное отличие в строении спинного мозга. У лягушки первый спинной нерв выполняет функцию XII черепно-мозгового нерва и иннервирует мышцы языка.
Головной мозг лягушки отличается от мозга рыб очень небольшим мозжечком, более крупным передним мозгом и хорошо заметным промежуточным мозгом (см. рис. II-26-II-28). Парные полушария переднего мозга имеют спереди крупные обонятельные луковицы, которые у бесхвостых амфибий срастаются между собой по средней линии. Промежуточный мозг расположен сразу за полушариями и имеет два дорсальных выроста, соответствующих эпифизу и парафизе. Нижний мозговой придаток промежуточного мозга — гипофиз — хорошо развит. Средний мозг амфибий представляет собой наиболее крупный отдел головного мозга. У большинства амфибий он формирует парные полусферы, в которые приходят зрительные тракты, продолжение зрительных нервов. Позади среднего мозга расположен очень маленький мозжечок. Он представляет собой небольшое дорсальное утолщение передней стенки IV желудочка. Полость IV желудочка открыта. Все черепно-мозговые нервы хорошо заметны, но XII пара отходит от центральной нервной системы вне мозгового черепа и поэтому часто ошибочно рассматривается как спинномозговой нерв. Добавочному (XI) нерву соответствует одна из ветвей блуждающего нерва (X). Блуждающий нерв иннервирует внутренние органы и гортань (производное жаберного аппарата рыб). У амфибий парасимпатические ветви блуждающего нерва начинают иннервировать лёгкие. Языкоглоточный нерв (IX) иннервирует полость рта, глотку и язык. Он
редко выходит из черепа отдельно, чаще сливаясь с блуждающим нервом. У бесхвостых амфибий тройничный нерв (V) может сливаться после выхода из мозга с лицевым и слуховым нервами (VI-VIII). Зрительный нерв (II) образует на
225
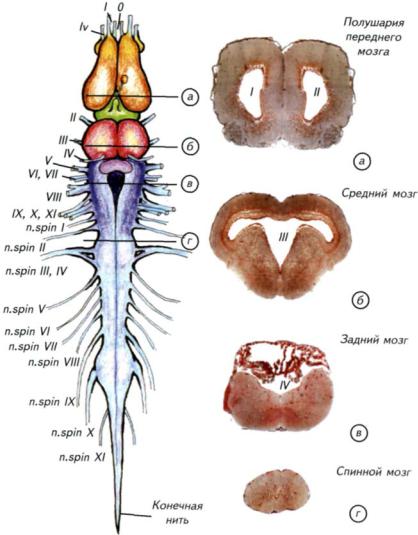
Рис. II-27. Головной и спинной мозг бесхвостой амфибии, основные нервы и микрофотографии гистологических срезов.
Слева — общий вид головного и спинного мозга амфибий на примере лягушки; а-г — гистологические срезы, соответствующие сечениям, указанным на левом рисунке. На срезах римскими цифрами обозначены мозговые желудочки, а на схеме головного и спинного мозга — черепно-мозговые нервы. Спинномозговые нервы — п.spin I-XI.
226
нижней стороне мозга срастание, напоминающее букву X, со сложным перекрёстом волокон, а у рыб это простое перекрещивание нервов. Обонятельный нерв (I) состоит из двух пучков волокон, которые у безногих амфибий разделяются и входят в мозг по отдельности. Один из них идёт от воздушного обонятельного мешка, а второй — от хеморецепторов якобсонова органа, который впервые появляется у амфибий и выполняет функции дополнительного органа обоняния (Panchen, 1967; Nilsson, 1983; Halpern, 1987).
Основные органы чувств амфибий изменены в соответствии с приспособлением к жизни в новой среде. У личинок и постоянно живущих в воде видов имеется сложная боковая линия. Она состоит из нескольких рядов чувствительных луковиц, которые позволяют воспринимать колебания воды. У наземных видов эти органы заменены специализированными осязательными пятнами. Органами вкуса являются луковицеобразные тельца, разбросанные в слизистой оболочке нёба и языка. Носовые мешки, имеющие две связанные камеры, способны к хеморецепции как воздуха, так и воды благодаря расположенным в них рецепторным клеткам. Они открываются ноздрёй наружу и специальным отверстием — хоаной в ротовую полость. Так как вдыхание и выдыхание воздуха происходят обычно через ноздри, носовая полость амфибий не только является обонятельным органом, но и входит в состав дыхательных путей.
Глаза амфибий позволяют различать цвета предметов и движущиеся объекты. Земноводные обладают хорошим зрением. Хрусталик их глаза сзади более выпукл, чем спереди, и в обычном состоянии установлен для зрения вдаль. Аккомодация глаза достигается сокращением лежащей вокруг хрусталика мышцы, перемещающей его по направлению к роговице. У безногих амфибий в связи с преимущественным
обитанием в почве глаза невелики или рудиментарны и спрятаны под кожей (см. рис. II-29, д).
Орган слуха включает среднее ухо, которое впервые встречается у амфибий. Снаружи оно ограничено барабанной перепонкой, за которой лежит большое пространство — барабанная полость, соединённая с ротовой полостью короткой евстахиевой трубой. В барабанной полости расположено стремя, передающее колебания барабанной перепонки на специальное окно внутреннего уха. Колебания воздуха вызывают движение барабанной перепонки, которое передаётся посредством стремени к эластичному окну внутреннего уха с механочувствительными клетками. Соединение евстахиевой трубы с ротовой полостью позволяет выравнивать давление воздуха в барабанной полости с атмосферным. Понятно, что такой орган слуха не отличается
227
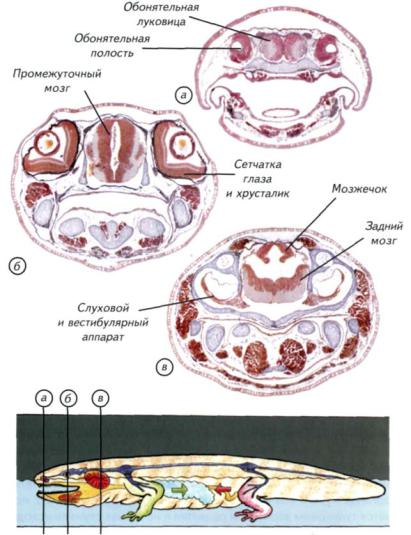
Рис. II-28. Гистологические срезы через голову кавказской саламандры (Mertensiella caucasica, Urodela).
Срезы соответствуют положению сечений на схеме (а-в). В связи с жизнью в лабиринтах сформировалась слуховая система, первоначально воспринимавшая колебания воды, а затем и воздуха. Водно-воздушное обоняние также сформировалось на границе сред. Форма и положение конечностей оптимально соответствуют движениям вперёд и назад в узких лабиринтах, что предопределило форму и положение коленных суставов.
228
высокой чувствительностью и не позволяет воспринимать высокие звуки. Тем не менее способность воспринимать колебания воздуха, а не воды впервые возникла у амфибий.
Адаптация амфибий к наземному существованию повлекла за собой изменение поведения этих животных. Амфибии больше, чем рыбы, способны к индивидуальному поведению, которое координируется центральной нервной системой. Они проявляют различные формы заботы о потомстве и способности к обучению. Земноводные могут проходить простые лабиринты. В этом отношении жабы несколько превосходят лягушек, а тритоны и саламандры с такой задачей не справляются.