

всего, таким животным было плоское червеобразное существо, похожее на современных турбеллярий.
Надо подчеркнуть, что только у кишечнополостных, плоских червей и головохордовых нервные клетки не унипополярны, как у остальных беспозвоночных. В этих группах нейроны имеют биполярную и мультиполярную организацию отростков, сходную с клетками нервной системы большинства позвоночных. Более того, только у кишечнополостных, плоских червей и головохордовых есть подэктодермальные нервные сплетения, которые не встречаются в других группах беспозвоночных. Тяж будущей центральной нервной системы возник после слияния дорсальных нервных стволов или поворота червеобразного предка хордовых на 90°. Это могло произойти при переходе к ильному существованию «на ребре» или без поворота. В обоих случаях результат был одинаков: один из нервных тяжей оказался расположен дорсально, а другой — вентрально. Из верхнего тяжа сформировалась центральная, а из нижнего — периферическая нервная система, иннервирующая внутренние органы. Полость нервной трубки возникла после продольного слияния ганглиев дорсального нервного ствола из
192
нейропиля, который освободился от отростков нервных клеток. Они ВЫШЛИ на внешнюю поверхность слившихся ганглиев, сформировав слои белого вещества вокруг тел клеток. Сегментация центральной нервной системы хордовых возникла в соответствии с количеством ганглиев и мышечных сегментов, иннервируемых из нервных стволов червеобразного предка, ползавшего по дну. Вентральные корешки спинного мозга сформировались из парных боковых комиссур, а дорсальные (смешанные) нервы возникли из чувствительных и моторных нервов дорсальной или латеральной части тела червеобразного предка.
193
Первичноводные позвоночные
Тип позвоночных (Vertebrata) включает в себя подтип бесчерепных (Acrania) и подтип черепных (Craniata). Единственными представителями бесчерепных являются многочисленные ланцетники (Amphioxi), которые рассматривались в предыдущем разделе, посвящённом происхождению позвоночных. Настоящий раздел охватывает обширную и разнообразную группу первичноводных черепных позвоночных. Они сформировались и долгое время морфологически изменялись в водной среде. В процессе эволюции именно этой группы сложилась нервная система, характерная для всех остальных позвоночных. Впервые возникло трубчатое строение нервной системы. Полости желудочков головного и спинного мозга стали непрерывными, появилась циркуляция внутрижелудочковой мозговой жидкости. В сочетании с эффективной кровеносной системой и внутренним скелетом это создало потенциальную возможность увеличивать размеры нервной системы первых водных позвоночных практически без ограничений. Эти эволюционные перспективы наращивания размеров мозга и параллельного увеличения сложности поведения не были реализованы в водной среде. Потребовались особые условия выхода на сушу, которые стали стимулом для развития новых качеств мозга и его ассоциативных функций (Northcutt, Davis, 1983).
Однако благодаря первичноводным позвоночным нервная система приобрела те универсальные черты, которые послужили морфологической основой для дальнейшей эволюции. Его конструкция оказалась настолько эффективной, что по существу не подверглась принципиальным изменениям за всю историю ЭВОЛЮЦИИ позвоночных (Nieuwenhuys, 1982). Конечно, мозг адаптивно перестраивался во всех систематических группах, но принципы структурной организации центральной и периферической нервной системы сохранились без изменений. На конструкцию нервной системы радикально не повлияли появление ни челюстного аппарата, ни плавников, ни конечностей, ни выход на сушу, ни адаптация к полёту, ни теплокровность.
Нервная система, как и позвоночник, оказалась тем органом, который обеспечивал выживание и прогресс всей группы позвоночных. Морфофункциональные отделы головного мозга, как и их основные функции, остаются неизменными на всём протяжении эволюции от миноги до человека. Эта универсальность строения нервной системы говорит об общности происхождения всех позвоночных. Делать предположения о полифилитичности происхождения позвоночных
194
довольно трудно. Допущение параллельного возникновения и эволюции сотен связанных между собой структур мозга весьма сомнительно.
§ 27. Мозг первичноводных позвоночных
Рассмотрим основные принципы строения универсальной конструкции нервной системы первичноводных позвоночных (рис. II-17; II-18). Её морфологической осью является центральная нервная система, которая расположена над позвоночником (рис. II-19, а) и включает в себя два отдела: головной и спинной мозг (см. рис. II-17, а). С головным и спинным мозгом соединены периферические нервы. Они проходят, переключаясь или не переключаясь, через ганглии и связывают периферические органы с центральной нервной системой.
Периферические нервы неодинаковы; часть из них двигательные, или эффекторные. Они передают сигналы из нервной системы к органам-мишеням. Такими органами могут быть поперечнополосатая или гладкая мускулатура, эндокринные железы или секреторные клетки. У ланцетника сами мышечные клетки образовывали отростки, соединяющие их с нервной трубкой. У остальных позвоночных ситуация обратная: двигательные отростки нейронов выходят из мозга или ганглиев, достигают мышц и оканчиваются на поверхности волокон, образуя нервно-мышечные синапсы.
Другая часть периферических нервов чувствительная. Они, наоборот, передают сигналы от внутренних органов и дистантных анализаторов в головной и спинной мозг. Концевые чувствительные участки рецепторных нервов очень сильно различаются по морфологическому строению и рецепторным возможностям. Это позволяет получать дифференцированную информацию различного типа. Особую группу
чувствительных клеток и волокон представляют собой дистантные рецепторные органы, или внешние органы чувств. У первичноводных позвоночных к внешним органам чувств относят обоняние, зрение, вкусовые рецепторы, органы боковой линии, рецепторы углового и линейного (гравитационного) ускорения, электрорецепторы и осязание.
Внешние органы чувств сосредоточены в головной части позвоночных, а их нервы приходят в головной мозг. В головной мозг приходит и большая часть информации от внутренних органов, мускулатуры и поверхности тела. По сути дела головной мозг является скоплением клеток, обрабатывающих информацию, поступающую от органов чувств. В соответствии с этими функциями головной мозг имеет специализированные центры, которые обслуживают несколько внешних или внутренних органов чувств. Такие центры по традиции называют отделами мозга. В головном мозге всех позвоночных
195
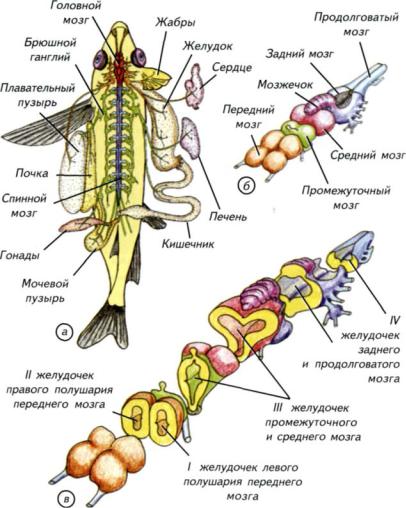
Рис. II-17. Основной план строения нервной системы черепных.
а — общий вид нервной системы и иннервируемых органов костистой рыбы. Синим цветом обозначен спинной мозг, красным — головной, жёлтым и зелёным — периферическая нервная система и головные нервы; б — головной мозг с основными анатомическими отделами. Мозжечок является частью заднего мозга; в — головной мозг с сечениями через основные отделы. Плоскости срезов жёлтые; б, в — цвета отделов совпадают.
196
выделяют 5 специализированных отделов: передний мозг (telencephalon), промежуточный (diencephalon), средний (mesencephalon), задний мозг с мозжечком (metencephalon, cerebellum) и продолговатый мозг (metencephalon) (см. рис. II-17, б). У первичноводных позвоночных отсутствуют полушария мозжечка, а следовательно, и мост, поэтому определить анатомическую границу между задним и продолговатым мозгом невозможно. Это приводит к тому, что во многих сравнительно-анатомических работах название «задний» и «продолговатый» мозг не применяется. Вместо них используется более общее название — «ромбэнцефалон» (rhombencephalon), которое включает в себя оба отдела.
Понятно, что отсутствие преемственности в названиях не сказывается на гомологизацию этих участков мозга как у первичноводных позвоночных, так и у млекопитающих. Гомология отделов головного мозга сохраняется во всех группах позвоночных (см. рис II-18). Надо отдать должное неординарному сравнительному нейроанатому H.H. Миклухо-Маклаю. Ещё в 70-х годах XIX в. он провёл детальные исследования головного мозга различных первичноводных позвоночных и их эмбрионов. Он показал, что основные отделы мозга позвоночных гомологичны друг другу и, несмотря на все эволюционные специализации, сохраняют общность

морфологического строения. Результатом многолетних трудов H.H. Миклухо-Маклая стала первая обоснованная гомологизация отделов мозга позвоночных (Миклухо-Маклай, 1952). Уже во 2-м издании сравнительной анатомии К. Гегенбаур (Gegenbaur, 1898) воспользовался этой работой и ввёл представление о нейрогомологиях отделов головного мозга в научный обиход.
Проводя морфологический анализ мозга первичноводных позвоночных, H.H. Миклухо-Маклай показал, что при изучении отделов мозга необходимо опираться как на внешнее строение, так и на морфологию внутренних полостей — мозговых желудочков. Они имеют специальные названия в каждом из отделов. В парных полушариях переднего мозга находятся первый (I) и второй (II) латеральные желудочки (см. рис. II-17, в; рис. II-20, a-в). Эти два желудочка соединены между собой межжелудочковым отверстием. Они переходят в третий (III) желудочек, который лежит внутри промежуточного и среднего мозга. В латеральных желудочках расположено непрерывное переднее сосудистое сплетение, которое распространяется примерно на 1/2 III желудочка (см. рис. II-20, а-в, д). В свою очередь он переходит в четвёртый (IV) желудочек, а последний продолжается в центральный канал спинного мозга (см. рис. II-17, б, в). Сверху IV желудочек прикрыт монослоем клеток и содержит заднее сосудистое сплетение (см. рис. II-20, к-м).
197
Рис. II-18. Романтический мир первичноводных позвоночных крайне разнообразен как по биологическим формам, так и по организации мозга.
Вверху — обитатели Атлантики в океанариуме Кейптауна (ЮАР); внизу — вид головного мозга различных видов хрящевых и костистых рыб.
а — акула-нянька (Ginglymostoma cirratum); б — длиннокрылая акула (Carcharhinus longimanus); в — радужная форель (Salmo irideus); г — чёрный марлин (Makaira indica); д — стерлядь (Acipenser ruthenus).
198
Важнейшую часть центральной нервной системы первичноводных позвоночных представляет спинной мозг. Граница между спинным и головным мозгом достаточно условна, поскольку отростки клеток из головного мозга проникают в спинной и наоборот (Nieuwenhuys, 1998). Головной мозг, как правило, расположен дорсально и окружён хрящами или костями черепной коробки. Спинной мозг лежит в полости позвоночного канала, который образован невральными дугами позвонков (см. рис. II-19, а).
У большинства позвоночных СПИННОЙ мозг имеет однотипную гистологическую структуру. Тела нервных клеток (серое вещество) обычно расположены вокруг центрального канала в виде классической «бабочки», свойственной большинству амниот. У анамний картина несколько смазана, и расположение тел нейронов на разрезах спинного мозга в виде «бабочки» обычно не встречается (см. рис. II-19, а, б). Верхнюю часть серого
вещества называют дорсальными (спинными) чувствующими рогами, а нижнюю — вентральными (брюшными) двигательными рогами спинного мозга. Спинные рога обычно содержат мелкие вставочные нейроны, а брюшные — крупные моторные. Через спинномозговые нервы осуществляются соматическая и висцеральная чувствительность, проходит соматически-двигательная и висцерально-двигательная иннервация.
Соматическая чувствительность включает в себя рецепцию кожных, сухожильных, связочных и мышечных сигналов. Висцеральная чувствительность включает вкусовую рецепцию и сигналы от внутренних органов. Соматически-двигательная иннервация обслуживает скелетные мышцы, а висцерально-двигательная — железы, глоточную, лицевую мимическую и челюстную мускулатуру, гладкую мускулатуру кожи, сосудов и внутренних органов. В состав спинного мозга традиционно включают ганглий дорсального корешка спинного мозга, или спинальный ганглий, в котором расположены тела афферентных (чувствительных) соматических и висцеральных нейронов (см. рис. II-19, а, б). Эти клетки связаны со спинным мозгом коротким отростком, который оканчивается в дорсальных рогах серого вещества. Вентральная часть спинного мозга составлена из эфферентных (двигательных) волокон. Они начинаются от клеток, расположенных в вентральных рогах спинного мозга (Савельев, 2001).
Не у всех позвоночных есть чёткое разделение корешков спинного мозга на моторные и сенсорные. У многих первичноводных позвоночных и амфибий висцеральные эфферентные волокна выходят из спинного мозга как через дорсальные, так и через вентральные корешки. У ланцетника, миног и миксин висцеральные эфферентные
199
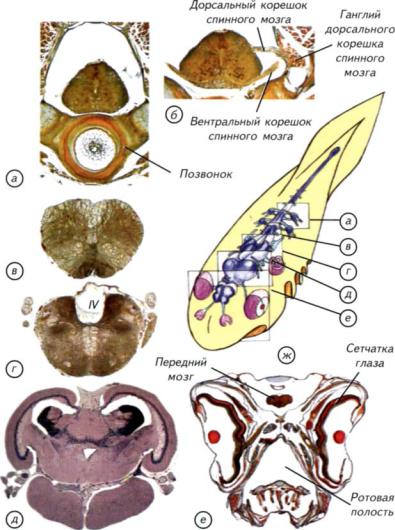
Рис. II-19. Гистологические срезы через голову и мозг рыбы.
а — через спинной мозг и позвонок с невральными дужками; б — через спинной мозг в месте отхождения моторных и сенсорных нервов; в — через продолговатый мозг; г — через задний мозг; д — через средний мозг; e — через голову в зоне полушарий переднего мозга. Обозначения сечений на схеме ж соответствуют буквенным обозначениям срезов. IV — четвёртый желудочек.
200
волокна преимущественно выходят через дорсальный корешок спинного мозга. Надо отметить, что СЛИЯНИЯ дорсальных и вентральных корешков спинного мозга у ланцетника и круглоротых не происходит. Они в виде самостоятельных нервов достигают иннервируемых органов. У большинства высших позвоночных дорсальные и вентральные корешки спинного мозга выходят на одном уровне. У ланцетника, миног, миксин и