

Концентрация головных и туловищных ганглиев членистоногих шла различными путями. Самым общим принципом стали укорочение комиссур и слияние ганглиев правой и левой стороны тела (см. рис. II-7, а-в). Этот процесс может сопровождаться рострокаудальным уменьшением количества нервных сегментов. В конечном счёте может происходить полная интеграция брюшной нервной цепочки до двух или одного анатомического центра: головных и грудных ганглиев. Среди насекомых своеобразным венцом совершенства организации такого типа являются двукрылые (Diptera). B этом отряде, как правило, брюшные нервные ганглии отсутствуют (см. рис. II-7, б). У десятиногих раков (Decapoda) хорошо выражены только головной и торакальный ганглии. При этом относительная длина продольных комиссур существенно меньше, чем у насекомых.
Однако латеромедиальное и каудоростральное слияние ганглиев может приводить к своеобразному строению нервной системы членистоногих и сходных с ними групп. Примером могут служить свободноживущие веслоногие ракообразные (Copepoda, Calanoida) (см. рис. II—7, г). Их головные и туловищные нервные ганглии сливаются в непрерывную структуру, которая мало напоминает нервную цепочку. Это связано с тем, что у этих в основном мелких (1-6 мм) ракообразных голова сливается с грудью, а поведение очень незатейливо. Находясь в толще воды, они являются специализированными фильтраторами. В этой группе ракообразных сердце стало редкостью, а полостные ЖИДКОСТИ передвигаются под механическим воздействием кишечника. Жабры обычно отсутствуют, а газообмен осуществляется через всю поверхность тела. Об уровне специализации веслоногих ракообразных говорит то, что края их жующих ротовых пластинок усилены кремниевыми соединениями, не встречающимися у других высших членистоногих. Надо отметить, что нервная система свободноживущих веслоногих ракообразных намного сложнее, чем у представителей этой группы, питающихся придонными осадками или ведущих паразитический образ жизни. Для последних характерно упрощение организации и уменьшение размеров всей нервной системы.
Самая высокая концентрация нервных ганглиев у ракушковых ракообразных (Ostracoda) (см. рис. II-7, д). Эти рачки обычно небольшого размера, у них утрачена сегментация. Тело рачка обычно заключено в двустворчатую хитинизированную и кальцифицированную раковину. Раковина часто может закрываться, а снаружи появляются только конечности. Биология этих ракообразных очень разнообразна. Среди них встречаются как фильтраторы, так и настоящие ХИЩНИКИ. ОНИ могут обитать и в морском грунте, и на суше, что говорит о
152
высокой адаптивности их нервной системы. Надо отметить, что у этих ракообразных встречается как партеногенетическое, так и половое размножение. Их половые органы крайне сложны в строении и использовании. Самцы должны ввести свой половой аппарат под панцирь самки и только потом выпустить сперматозоиды в защищённую полость. После оплодотворения самка обычно откладывает яйца в какое-либо укрытие. Ракушковые ракообразные проявляют своеобразную заботу о потомстве, поскольку оплодотворённые яйца могут развиваться до вылупления под раковиной матери. Все половые процессы у ракушковых ракообразных невероятно затруднены тем, что у многих видов длина сперматозоидов в несколько раз больше длины тела. Так, у представителей рода Pontocypris сперматозоид в 8 раз длиннее тела взрослой особи и достигает 6 мм. Ракушковые ракообразные являются своеобразными сексуальными рекордсменами, поскольку и относительные, и абсолютные размеры их сперматозоидов не имеют аналогов среди животных.
Таким образом, у членистоногих и сходных с ними групп мы видим значительное разнообразие анатомического строения брюшной нервной цепочки и головных ганглиев. Однако они вписываются в общий архетип и обеспечивают сложное инстинктивное поведение. Поведение свободноживущих форм как при лестничной нервной системе, так и при единственном цереброторакальном ганглии существенно не различается. В обоих случаях мы видим многоступенчатые инстинкты, заботу о потомстве, сложное пищевое и половое поведение. Структурная организация головных ганглиев и брюшной нервной цепочки даёт идеальную возможность для создания универсальной системы с запрограммированным, но адаптивным поведением.
Брюшной нервной цепочкой далеко не исчерпываются варианты строения нервной системы беспозвоночных. Существует множество групп этих животных, достойных рассмотрения как с позиций строения нервной системы, так и в связи со сложностью поведения. Весьма демонстративна организация моллюсков (Mollusca). У этих беспозвоночных встречаются самые контрастные формы строения нервной системы с невероятным разнообразием поведения. У моллюсков известны примитивнейшие нервные системы, состоящие из нескольких асимметричных ганглиев ИЛИ вообще без них. Такой тип нервной системы характерен для малоподвижных ИЛИ прикреплённых животных, и среди моллюсков есть настоящие рекордсмены. Самое крупное неподвижное животное на этой планете — тридактна (Г. gigas) достигает вместе с раковиной массы более 200 кг. Хорошо известны и свободноживущие брюхоногие (Gastropoda) и головоногие
153
(Cephalopoda) моллюски со сложным инстинктивным поведением и способностью решать сложные экспериментальные задачи, не встречающиеся в естественных условиях обитания.
§ 23. Нервная система моллюсков
Наибольший морфофункциональный контраст представляют собой организация нервной системы головоногих и двустворчатых моллюсков (рис. II-9; II-10, а). У двустворчатых моллюсков есть парные головные, висцеральные и педальные ганглии, соединённые комиссурами (см. рис. II-10, а). Нервные стволы, идущие от головных ганглиев, обычно замкнуты. Головные ганглии (cerebral ganglion) могут быть построены по различному типу. У мидий нейроны расположены в самом нейропиле ганглия (см. рис. II-9, и), а у сердцевидок и шаровок — по его периферии (см. рис. II-9, г). Различия в гистологическом строении ганглиев
не сказываются на их функциях. Головные ганглии в обоих случаях иннервируют ротовые лопасти, передний аддуктор и мантию. Головные ганглии двустворчатых моллюсков составного типа. Они образовались в результате слияния церебрального и плеврального ганглиев. Висцеральный ганглий (visceral ganglion) также образовался в результате слияния с париетальным ганглием. Он обычно несколько меньше головного и расположен в каудальной части моллюска (см. рис. II-9, д). Висцеральный ганглий иннервирует мышцы ретрактора, заднего аддуктора, сердце, почку, сифон, половые органы и жабры. Пара педальных ганглиев (pedal ganglion) иннервирует мускулатуру ноги (см. рис. II-9, е). Они меньше других ганглиев, но имеют разветвлённые эффекторные отростки, оканчивающиеся среди мышц. Таким образом, нервная система двустворчатых моллюсков представлена всего тремя парами ганглиев, которые образовались в результате слияния. Это упрощение нервной системы связано с малоподвижным образом жизни и отсутствием рецепторных головных структур. Органы чувств у двустворчатых моллюсков достаточно специфичны и распределены по всему телу. На мантии, сифонах и жаберных нитях встречаются вторичные глаза, которые не связаны непосредственно с головным ганглием. Они имеют относительно сложное строение и могут воспринимать не только освещённость, но и направление её изменения. Это позволяет некоторым двустворчатым моллюскам перемещаться, пользуясь ногой или применяя реактивное движение — хлопая створками, как морской гребешок. У двустворчатых моллюсков хорошо развиты хеморецепторы — осфрадии и механорецепторные осязательные клетки. Есть и статоцисты, которые локализованы в крае ноги вблизи от педального ганглия.
154
Двустворчатые моллюски в основном являются малоподвижными фильтраторами с довольно пассивным образом жизни. Среди них относительно активны только камнеточцы и корабельные черви. Камнеточцы делают ходы в камнях специальными зубчиками или при помощи кислоты, выделяемой железами мантии. Размножение сводится к откладке одиночных яиц и их наружному оплодотворению. Забота о потомстве практически отсутствует, хотя яйца и могут развиваться на жабрах.
По сравнению с двустворчатыми моллюсками брюхоногие моллюски (Gastropoda) являются образцом активного образа жизни и сложного поведения. Это единственные моллюски, перешедшие к наземному образу жизни. Их тело разделено на голову, ногу и туловище. В голове сосредоточены церебральные ганглии и основные органы чувств. Обычно это две пары щупалец и глаза, имеющие довольно сложное строение. Глаза могут представлять собой элементарные светочувствительные ямки или сложные системы с хрусталиком и аналогом стекловидного тела. На щупальцах расположены органы осязания и обоняния. Контактные хеморецепторы представлены осфрадиями, а органы равновесия — статоцистами. Статоцисты являются важнейшим органом чувств, особенно для наземных брюхоногих. Это отражено и в их иннервации. Как правило, статоцисты иннервируются волокнами головного ганглия.
Нервная система брюхоногих представлена 5-6 основными парами ганглиев, которые связаны между собой продольными нервными стволами и поперечными комиссурами (см. рис. II—8, д, е). Головные ганглии расположены над глоткой и иннервируют голову, глаза, щупальца и статоцисты. Плевральные ганглии расположены позади головных и иннервируют переднюю часть мантии. Под пищеводом расположены буккальные ганглии, иннервирующие глотку, пищевод и желудок. Этот комплекс из трёх сближенных ганглиев связан несколькими комиссурами и представляет собой головной центр — своеобразный мозг гастропод. Педальные ганглии служат исполнительным механизмом управления ногой. Висцеральные ганглии могут лежать сближенной парой или полностью сливаться. Они иннервируют внутренние органы: почки, половые органы, кишечник и сердце. Отдельно лежащая пара париетальных ганглиев иннервирует дыхательную систему жабр — ктенидий и хеморецепторные осфрадии. У брюхоногих в связи с развитием асимметрично закрученной раковины возникает торсия — перекручивание продольных нервных стволов. Это приводит к тому, что париетальные ганглии левой и правой стороны тела меняются местами, как показано на рис. II-8, е. Существует
155
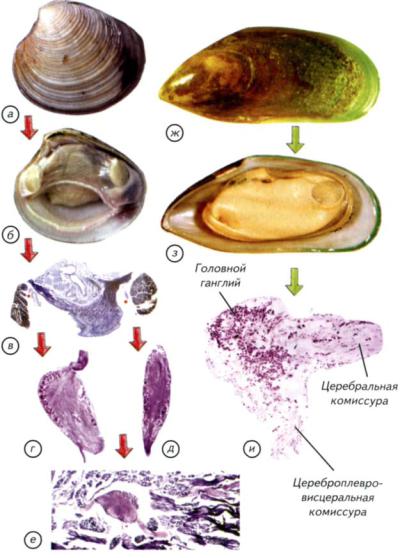
Рис. II-9. Нервная система двустворчатых моллюсков (Bivalvia).
а-е — С. galliпа corrugatula; ж-и — P. canaliculus. Красными точками на микрофотографии (в) показана локализация увеличенных ганглиев (г, д); г, и — головные ганглии (cerebral ganglion); д — висцеральный ганглий (visceral ganglion); e — педальный ганглий (pedal ganglion). Стрелками показаны этапы препаровки моллюсков.
156
несколько вариантов торсии, что зависит от расположения жаберного аппарата или лёгких.
Относительно скромная организация нервной системы брюхоногих моллюсков оказалась весьма эффективной как в воде, так и на суше. Большинство брюхоногих раздельнополы, хотя встречается и гермафродитизм с чередованием мужских и женских периодов размножения. В большинстве случаев гастроподы — подвижные животные, которые питаются растениями или хищничают. Однако встречаются и прикреплённые колониальные формы, такие, как каури. Подобные фильтраторы являются скорее исключением среди гастропод, чем правилом.
Поведение брюхоногих не отличается интеллектуальными поступками и достаточно однообразно. Обычно брюхоногие питаются растительными остатками, непосредственно растениями или грибами. В этом случае их активность сводится к поиску пригодной пищи. Однако среди брюхоногих есть и настоящие хищники. Примером агрессивного хищника является тестацелла (Г. maugei), которая питается дождевыми червями. Она обладает достаточно развитыми органами чувств, чтобы охотиться на дождевых червей в их норах. Многие хищные улитки охотятся на себе подобных, но не брезгуют никакой животной пищей. В той или иной степени к хищничеству могут прибегать представители разнообразных семейств.
Особую популярность в качестве объекта зоопсихологии приобрели головоногие моллюски (Cephalopoda). Осьминоги, кальмары и каракатицы являются, по мнению исследователей их поведения, своеобразными приматами мира беспозвоночных. Они ведут преимущественно активный образ жизни, перемещаясь по дну на многочисленных конечностях или плавают, используя принцип реактивного движения. Выталкивание воды из мантийной полости позволяет им быстро плавать в толще воды, но только задом наперёд. У головоногих в крайней форме проявляется цефализация нервных ганглиев (см. рис. II-10, в, ж, з). Ганглии с различными функциями сливаются над или под пищеварительной трубкой и образуют крупный головной ганглий. Поскольку головоногие могут достигать 20-метровой длины, размеры их головного ганглия могут быть
сопоставимы с размерами мозга собаки. При таких размерах конструкции и традиционной для беспозвоночных ганглиозной организации нервной системы трудно представить себе эффективное метаболическое обеспечение. Именно размеры головных ганглиев головоногих неизбежно привели к возникновению внутримозговых полостей. У большинства головоногих моллюсков головные ганглии внутри полые. Однако надо оговориться, что речь идёт только о надглоточной части головного ганглия.
157
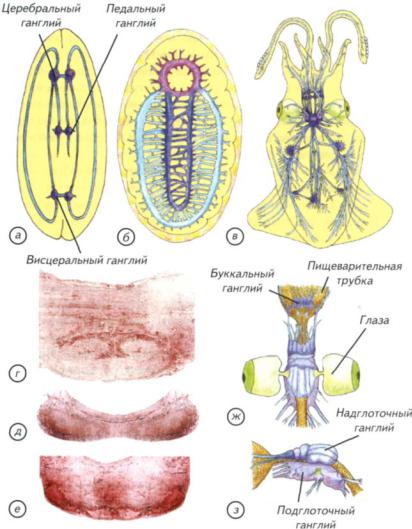
Рис. II-10. Нервная система двустворчатых, панцирных и головоногих моллюсков.
Схема строения нервной системы двустворчатых моллюсков (а) и хитона (б); внешний вид нервной системы (в) и головного мозга (ж, з) кальмара; микрофотографии срезов головных ганглиев осьминога (Octopus vulgaris): r — брахиальный ганглий; д — висцеральный ганглий; e — церебральный ганглий.
158
Подглоточная часть обычно не имеет внутренней полости. Разделение головного ганглия на подглоточный и надглоточный отделы функционально вполне обосновано (см. рис. II-10, ж, з). Подглоточная часть головного ганглия состоит из брахиальных, педальных и висцеральных ганглиев, объединённых общим нейропилем (см. рис. II-10, г, д). Педальные ганглии уже внутри общей структуры разделены на ганглии щупалец и ганглии воронки. Брахиальные и педальные ганглии иннервируют щупальца и мантийную полость. Висцеральные ганглии отвечают за иннервацию внутренних органов и содержат центр автоматической регуляции дыхания.
Полый внутри надглоточный ганглий является своеобразным интеллектуальным центром головоногих. В нём выделяют от 3 до 6 отделов. В самом сложном случае к ним относят буккальные ганглии (ротовые), три пары центральных и две пары церебральных ганглиев. Буккальные ганглии вынесены вперёд, не сливаются с головным ганглием и отвечают за иннервацию ротовой зоны передней части пищеварительной системы (см. рис. II-10, ж). Самыми ростральными надглоточными (уже головными) являются центральные ганглии, которые расположены поперёк оси головного центра. Их передняя пара контролирует механизм захвата, удержания, подведения к клюву, расчленения и заглатывания пищи; 2-я и 3-я пары центральных ганглиев хорошо развиты в связи с тем, что они отвечают за гидродинамическую систему плавания и контролируют обновление воды в воронке. Эти процессы осуществляются не непосредственно, а через контроль за исполнительными висцеральными центрами. Эти же центры контролируют перемещения головоногих при помощи щупалец, отвечают за осязательные ощущения и изменения направления движения при плавании. В центральных ганглиях находятся и центры управления окраской животных.
Надо отметить, что головоногие могут очень широко изменять свою окраску из-за прямой иннервации
пигментных клеток. Нервная система имеет непосредственные окончания на каждой пигментной клетке, что позволяет головоногим динамично изменять свой цвет при маскировке и смене настроения. Такой тип управления пигментацией позволил каракатицам использовать смену окраски тела как своеобразный цветовой язык. Во время брачных игр и оплодотворения самцы и самки ведут переговоры на языке смены цветовой гаммы покровов, поэтому не удивительно, что в тех же центральных ганглиях находятся и зрительные зоны. Туда приходит информация от глаз, которые у головоногих моллюсков самые большие среди животных. У крупных
159
глубоководных головоногих описаны глаза диаметром 40 см — абсолютный рекорд для фоторецепторных систем. Позади центральных ганглиев расположены две или три пары продольно ориентированных церебральных ганглиев (см. рис. II-10, е, ж, з). Они непосредственно не связаны ни с какими конкретными моторными или сенсорными системами.
Церебральные ганглии — своеобразная ассоциативная надстройка над подглоточными и надглоточными ганглиями. Они необходимы для принятия поведенческих решений, что предопределяет развитость их связей со всеми центрами головных ганглиев. Церебральные ганглии являются своеобразным аналогом ассоциативных «лобных долей» высших млекопитающих и человека. Однако приматам придётся скромно поджать хвост (у кого он есть), если сравнить относительные размеры лобной доли человека и церебральных ганглиев небольшого осьминога. Если мозг человека и осьминога уравнять в размерах, то ассоциативные центры головоногих окажутся в 3 раза больше, чем лобные доли человека. Эти достойные различия сказываются и на поведении головоногих.
Особое развитие их мозга связано с появлением головных щупалец, которые могут быть дифференцированы по функциям. Эти своеобразные манипуляторы требуют для своего управления развитого мозга. Обладая развитыми щупальцами, головоногие могут прекрасно различать предметы на ощупь, вырабатывать устойчивые навыки и надолго запоминать полученный опыт. У них прекрасно развита исследовательская деятельность. Они сами обучаются открывать банки, отвинчивать крышки, открывать двери и совершать многие другие антропоморфные действия, привыкают к экспериментаторам, узнают их индивидуально, умеют бесцельно играть с предметами и решать многоступенчатые задачи.
Головоногие моллюски давно бы стали безусловными интеллектуальными лидерами нашей водянистой планеты, но природа сыграла с ними довольно злую шутку. Наградив их развитым ассоциативным мозгом, сверхэффективными манипуляторами, осязанием, хеморецепторами и зрением, она лишила их времени — главного условия для накопления индивидуального опыта. Головоногие моллюски живут очень недолго, обычно не более трёх лет. Кроме того, большая часть видов этой группы моноцикличны. Они рождаются, растут, один раз размножаются и погибают. Продолжительность жизни в 1 год ограничивает реализацию интеллектуальных возможностей головоногих. Живи они подольше, и приматов могло бы не быть, а сушу осваивали бы каракатицы в скафандрах.
160
Среди головоногих как бы в насмешку существуют и настоящие долгожители — наутилусы (Nautiloidea). Все 6 видов этих головоногих обладают раковиной, которую они заполняют газом и добиваются нейтральной плавучести. Они плохо плавают, у них очень примитивно организованная нервная система. Окологлоточное нервное кольцо не имеет выраженных центров, как у других головоногих. Наоборот, нейроны не собраны в концентрированные ганглии, а довольно равномерно распределены по нервным стволам и комиссурам. Вместо головных ганглиев у наутилусов есть своеобразные нервные тяжи, которые огибают переднюю часть пищеварительной трубки. Над ней проходит дорсальный тяж, гомологичный церебральным ганглиям. Под пищеварительной трубкой проходят две комиссуры. Они гомологичны подпищеводным ганглиям. Передняя соответствует педальным ганглиям, а задняя — плевровисцеральным ганглиям. От последних идут два укороченных ствола, которые заканчиваются оформленными висцеральными ганглиями. Подобная распределённость нейронов по нервным тяжам характерна только для примитивных параллельных ганглиев кишечнополостных и панцирных моллюсков. Понятно, что столь примитивная «доганглиозная» нервная система не может претендовать на морфологический субстрат интеллекта. Однако именно наутилусы живут достаточно долго, чтобы приобрести индивидуальный опыт, который, правда, негде хранить.
Среди моллюсков особое положение занимает примитивная группа панцирных моллюсков, или хитонов (Loricata). Это уплощённые билатерально-симметричные моллюски с большим щитовидным панцирем на СПИННОЙ поверхности. Он состоит из 8 пластин, которые расположены, как черепица на крыше. Обтекаемая форма тела хитонов способна выдерживать прибрежные волны, а мускулистая подошва создаёт разрежение, удерживающее животных на субстрате. Голова анатомически отделена небольшой бороздкой от ноги, но на ней нет головных щупалец или специализированных глаз. Светочувствительными органами являются особые раковинные глазки, которые не встречаются у беспозвоночных других групп. Глазки появляются по мере роста раковинных пластинок около их края. Общее число этих простых глазков может достигать нескольких тысяч. Кроме светочувствительных глазков, у хитонов есть рецепторы давления и глоточные вкусовые рецепторы. Присутствуют и хеморецепторы — осфрадии, которые находятся возле жабр. Понятно, что от животных с такими органами чувств трудно ожидать занимательного пищевого или полового поведения. В основном они поедают растительные остатки, хотя не брезгуют и животной пищей. Размножение хитонов обычно
161
примитивно и состоит в выбрасывании в воду половых продуктов самками и самцами. Вероятностное оплодотворение компенсируется количеством яиц и сперматозоидов — некоторых видов бывает более миллиона. Однако при вынашивании потомства в жаберных складках число яиц снижается в тысячи раз.
Эта группа уникальна не столько поведением задумчивых моллюсков, поедающих всё, что можно найти в приливной зоне морей и океанов, сколько нервной системой. Под панцирем и жёсткой мантией скрывается одна из самых древних нервных конструкций среди билатерально-симметричных животных планеты. Основой