

Протисты / Карпов С.А. Строение клеток протистов
.pdfГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
Рис. 4.89. Схема строения пузулы у динофлагеллаты Codinium. (Ïî: Hausmann, Hülsmann, 1996.)
ок – общая камера, в которую выводится содержимое из собирающих каналов (ск); тп – текальные пластинки.
4.6.3. Аппарат Гольджи
Он обнаружен практически у всех исследованных видов протистов. До изучения с помощью электронного микроскопа эта органелла была известна у протистов, как парабазальное тельце, которое наиболее сильно развито у трихомонадид и гипермастигид – парабазалий (рис. 2.43, 2.44).
280

4.6. ДРУГИЕ ЦИТОПЛАЗМАТИЧЕСКИЕ ОРГАНЕЛЛЫ
Чаще всего аппарат Гольджи расположен по соседству с ядром и представлен одной или несколькими стопками плоских цистерн – диктиосомами, – окруженных мелкими пузырьками (рис. 4.90). Однако аппарат Гольджи не всегда встречается в виде диктиосом. У некоторых протистов он представлен одиночными цистернами. Отсутствие диктиосом обычно трактуется как примитивный признак. Вероятно, это вполне справедливо, т.к. стопки цистерн аппарата Гольджи работают эффективнее, чем одиночные цистерны. Однако отсутствие диктиосом у современных протистов не может однозначно свидетельствовать о примитивности этих организмов, т.к. диктиосомы – весьма лабильные образования, а их формирование и разборка в значительной степени зависит от внешних воздействий на клетку (например, снижение концентрации кислорода в среде) или от физиологических перестроек самого протиста (переход к инцистированию).
Рис. 4.90. Схема строения типичного аппарата Гольджи. (По: Заварзин и др., 1992.)
1 – цис-, 2 – средняя и 3 – транс-части диктиосомы, 4 – цистерны, 5 – транспортные и 6 – секреторные пузырьки, 7 – распределительный отдел транс-части диктиосомы.
281

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
4.6.4. Лизосомы и другие органеллы и включения
Âклетках протистов, как и в клетках многоклеточных животных, присутствуют лизосомы. Эти цитоплазматические тельца в форме мелких пузырьков (первичные лизосомы) образуются в аппарате Гольджи. В них локализованы пищеварительные гидролитические ферменты. На электронограммах первичные лизосомы выглядят как тельца округлой формы с гомогенным внутренним содержимым, диаметром меньше 1 мкм. Они расположены во всех участках клетки. Вторичные лизосомы, или пищеварительные вакуоли, хорошо выражены только у гетеротрофных протистов, питающихся путем фагоцитоза.
Âэндоплазме разных протистов в большем или меньшем количестве присутствуют резервные питательные вещества, используемые в процессах метаболизма. Чаще всего это различные полисахариды, нередко – липиды и иные жировые включения. Для некоторых групп протистов характерны специфические запасные вещества. Например, эвгленовые запасают парамилон, который не встречается у других протистов, а красные водоросли – багрянковый крахмал. Количество резервных веществ зависит от физиологического состояния простейшего, характера
èколичества пищи, от стадии жизненного цикла и поэтому варьирует в широких пределах.
Аппарат проникновения в клетку
Паразитические протисты обладают различными структурами для проникновения в клетку хозяина. Одной из широко распространенных структур такого рода является апикальный комплекс. Он характерен прежде всего для споровиков (рис. 4.91)
èхищных жгутиконосцев из отряда Colpodellida (=Spiromonadida). Ранее его считали уникальным признаком типа Apicomplexa. В его состав входит коноид, апикальные кольца и полярное кольцо с отходящими от него микротрубоч- ками (рис. 4.91). Внутри апикального комплекса обычно обнаруживаются роптрии и микронемы (рис. 2.36 А). Роптрии – крупные мешковидные образования железистого типа, откры-
282

4.6. ДРУГИЕ ЦИТОПЛАЗМАТИЧЕСКИЕ ОРГАНЕЛЛЫ
вающиеся на переднем конце клетки. Они выделяют специальные белки (секрет проникновения), которые, по-видимому, участвуют в разрушении покровов клетки хозяина или жертвы. Микронемы представлены многочисленными мелкими извитыми канальцами, выполняющими, вероятно, аналогичную функцию.
Менее сложный апикальный комплекс обнаружен также у хищного бесцветного жгутиконосца Katablepharis, имеющего на апикальном конце клетки коноидальное кольцо, от которого отходят микротрубочки. Внутри конуса микротрубочек обнаружены и подобные микронемам структуры.
Апикальный комплекс колподеллид и катаблефариса служит для питания, а у споровиков – для проникновения в клетку, но, по-видимому, основная его функция – разрушение покровов клетки.
По-иному устроен аппарат проникновения в клетку у других внутриклеточных паразитов. У микроспоридий и плазмо-
Рис. 4.91. Строение коноида Toxoplasma gondii. (Ïî: Nichols, Chiappino, 1987.)
вмк – внутренний мембранный комплекс, вмт – внутренние микротрубочки, зпк – заднее преконоидальное кольцо, кп – комплекс полярного кольца, мт – 22 микротрубочки, пл – плазмалемма, ппк – переднее преконоидальное кольцо, ск – субъединицы коноида.
283

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
диофорид существует для этой цели специальный аппарат экструзии (рис. 4.92). Принцип его действия у представителей обеих групп один и тот же. Внутри споры находится небольшая вакуоль, которая в начале процесса экструзии быстро набухает и выталкивает из споры свернутую трубку. У плазмодиофорид эта трубка сравнительно короткая и имеет на конце специальный стилет, пробивающий клеточную стенку растений. У микроспоридий она может достигать значительной длины, не несет на конце какого-либо вооружения, поскольку пробивает менее прочные покровы (плазмалемму) клеток многоклеточных. Зародышевая плазма с ядром передвигается по этой трубке из споры паразита и попадает в клетку хозяина. Поскольку микро-
Рис. 4.92. Схема проникновения плазмодиофориды в клетку растения. (По: Dylewski, 1990.)
А – интактная спора на поверхности клетки, Б – прикрепление споры к клеточной стенке растения, В – пробивание клеточной стенки специальным наконечником, Г – переход спороплазмы из споры в клетку растения, который вызывается набуханием вакуоли (в) на этапах Б–Г. кс – клеточная стенка, на – наконечник, сп – спора, я – ядро спороплазмы.
284

4.6. ДРУГИЕ ЦИТОПЛАЗМАТИЧЕСКИЕ ОРГАНЕЛЛЫ
споридии и плазмодиофориды не связаны между собой филогенетически, можно предполагать, что такой аппарат проникновения в клетку возник в этих двух группах независимо.
Аппарат питания
Аппарат питания безусловно и неразрывно связан с функцией питания, поэтому его следовало бы рассматривать в главах, посвященных физиологии клетки. Однако его строение у некоторых протистов составляет существенную часть цитоскелета. Кроме того, при сравнении его с другими структурами клетки возникают аналогии с особенностями строения аппарата проникновения в клетку.
Принято выделять два основных способа поглощения пищи у гетеротрофов: пиноцитоз и фагоцитоз. Пиноцитоз представляет собой по сути дела заглатывание растворенных органических веществ или мелких частиц, посредством образования мелких пузырьков с обкладкой из белковой клатриновой сеточки (окаймленные пузырьки). В клеточной биологии для описания этого процесса принято использовать термин «рецепторный эндоцитоз». Он характерен, по-видимому, для всех протистов независимо от наличия других способов питания. Его распространение лимитируется лишь образованием жестких покровных структур вроде пелликулы или клеточной стенки. Так называемое «всасывание всей поверхностью клетки» как раз и представляет собой пиноцитоз.
При фагоцитозе, или голозойном питании, происходит заглатывание крупных объектов. Разнообразие способов заглатывания пищи довольно велико. Здесь мы остановимся только на тех, которые требуют специальных сложных систем заглатывания пищи.
Еще один способ гетеротрофного питания – мизоцитоз – характерен для хищных протистов и представляет собой высасывание цитоплазмы жертвы после разрушения ее покровов.
По способу заглатывания пищевых объектов протистов можно разделить по меньшей мере на 3 группы:
1) одни заглатывают как мелкие, так и крупные объекты при помощи псевдоподий (амебы, воротничковые жгутиконосцы, солнечники, церкомонады, хризомонады и другие);
285

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
2)другие (инфузории, многие жгутиконосцы), так называемые фильтраторы, или седиментаторы, постоянно подгоняют жгутиками или ресничками мелкие пищевые объекты (преимущественно бактерий) в ротовое отверстие;
3)третьи захватывают и заглатывают крупные пищевые объекты, а также высасывают жертву (мизоцитоз) при помощи специальных органелл цитостома и цитофаринкса (инфузории, жгутиконосцы).
При захвате пищи при помощи псевдоподий постоянные ротовые структуры отсутствуют, а сам процесс хорошо известен и обеспечивается микрофиламентами цитоскелета. Иногда ему предшествует иммобилизация пищевого объекта при помощи слизи или экструсом. Надо заметить, что такой тип захвата пищи характерен не только для амебоидных организмов. Им широко пользуются протисты с другой организацией тела – жгутиконосцы, солнечники, радиолярии.
Рис. 4.93. Схема расположения кинетосом и связанных с ними фибрилл в цитостомальной зоне инфузории
Tetrahymena. (Ïî: Sleigh, 1989.)
пф – посторальные фибриллы, ск – соматические кинетосомы, ум – кинетосомы «ундулирующей мембраны», 1–3 – кинетосомы соответствующих мембранелл.
286

4.6.ДРУГИЕ ЦИТОПЛАЗМАТИЧЕСКИЕ ОРГАНЕЛЛЫ
Óфильтрующих организмов имеется постоянное ротовое отверстие и глоточный аппарат. Наиболее характерен этот способ заглатывания пищи для инфузорий, которые часто имеют высоко дифференцированную ротовую (оральную) цилиатуру (рис. 4.93). Многие прикрепленные жгутиконосцы также являются фильтраторами, часто с постоянным местом захвата пищи, но цитостом/цитофарингальные структуры формируются не всегда, а захват пищи идет при помощи псевдоподий.
Наиболее сложные ротоглоточные образования характерны для хищников. Процесс заглатывания пищи предполагает сна- чала обездвиживание жертвы при помощи парализующих ее экструсом, а затем заглатывание целиком или высасывание цитоплазмы. Интересно, что в обоих случаях, будь то ротоглоточ- ный аппарат хищной инфузории или щупальце суктории, используется одинаковая конструкция – цитофарингальная корзинка (рис. 4.94–4.96). Непосредственно под плазмалеммой глотки проходят одиночные микротрубочки с отходящими от
Рис. 4.94. Схемати- ческое изображение ротового аппарата хищных эвгленовых на поперечном срезе. (По: Margulis et al., 1993.)
1 – плазмалемма глоточного канала, 2 – микротрубочки, 3 – фибриллярные пластинки.
287
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
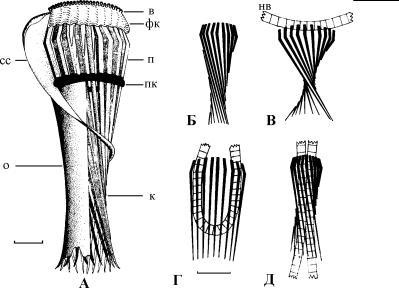
Рис. 4.95. Схема строения и функционирования глоточного аппарата у инфузории Nassula. (Ïî: Tucker, 1968.)
А – общий вид и строение глоточного аппарата, Б – состояние корзинки у непитающейся особи, В–Д – последовательные изменения конфигурации корзинки при захвате и заглатывании нитчатой водоросли (нв). в – воротничок на апикальном конце глоточного аппарата, к – корзинка глотки, образованная палочками (п), которые состоят из микротрубочек, о – общая оболочка корзинки, пк – плотное кольцо, связывающее палочковый аппарат, сс – серповидная складка, фк – фибриллярное кольцо. Масштабная линейка: 5 мкм.
них фибриллярными пластинками, или ленты микротрубочек. Глубже в цитоплазме скелет глотки составляют пучки связанных между собой микротрубочек (немадесмы), часто в виде треугольных призм (рис. 4.94). При прохождении крупного объекта глотка растягивается, а немадесмы служат опорными структурами. Так, в сложно устроенной глотке Nassula происходит последовательное пространственное преобразование скелетных элементов глотки, что позволяет этой инфузории перед заглатыванием складывать длинную нить водоросли пополам (рис. 4.95).
288

4.6. ДРУГИЕ ЦИТОПЛАЗМАТИЧЕСКИЕ ОРГАНЕЛЛЫ
Транспортировка же пищи обеспечивается ламеллами, которые связаны с микрофиламентами. В области ламелл обнаруживается АТФ-азная активность, а среди белков найден актин. По-видимому, транспорт пищевого объекта осуществляется за счет известного механизма скольжения вдоль микротрубочек при помощи филаментов. При этом филаменты прикрепляются к наружной поверхности мембраны, окружающей жертву. Этот механизм осуществляется, по-видимому, и при всасывании цитоплазмы жертвы сосущими инфузориями (рис. 4.96).
Принципиально так же устроена глоточная система у кинетопластид, хищных бесцветных жгутиконосцев и в щупальце фаготрофных динофитовых. Хорошо развитая ротоглоточная система свободноживущих кинетопластид и эвгленовых является существенной частью цитоскелета клетки, обеспечивая своеобразие формы клетки этих протистов.
4.6.5. Фоторецептор
Фоторецепторный аппарат характерен для водорослей и обычно состоит из глазного пятна (глазка, или стигмы) и той или иной части жгутикового аппарата (рис. 4.74, 4.97). Чувствительность к свету, однако, выражена не только у водорослей, но и у многих гетеротрофных протистов, хотя они и не имеют специальных структур для фоторецепции.
Принцип работы фоторецептора тот же, что и у животных. Глобулы глазка поглощают свет, который отражается от лежащих за ними мембран или фибриллярных уплотнений и фокусируется на определенном участке жгутика, в котором обычно расположен собственно рецепторный аппарат.
Строение фоторецепторов среди водорослей весьма различ- но, но всех их объединяет наличие следующих необходимых для фоторецепции элементов: 1) богатые каротиноидами липидные глобулы (собственно стигма, или глазок), расположение которых варьирует от беспорядочного до высокоупорядоченных гексагональных структур; 2) расположение глазка (обычно одиночного) на периферии клетки, которое четко определено по отношению к жгутику или плоскости его биения; 3) связь стиг-
289